Opposite Roles of Tra2β and SRSF9 in the v10 Exon Splicing of CD44

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture, Transfection, RNA Extraction, and RT-PCR
2.2. Plasmid Construction
2.3. RNA Immunoprecipitation Assay
2.4. Immunoblotting Assay
2.5. RNA-Binding Motifs Prediction
2.6. Quantitation and Statistical Analyses
3. Results
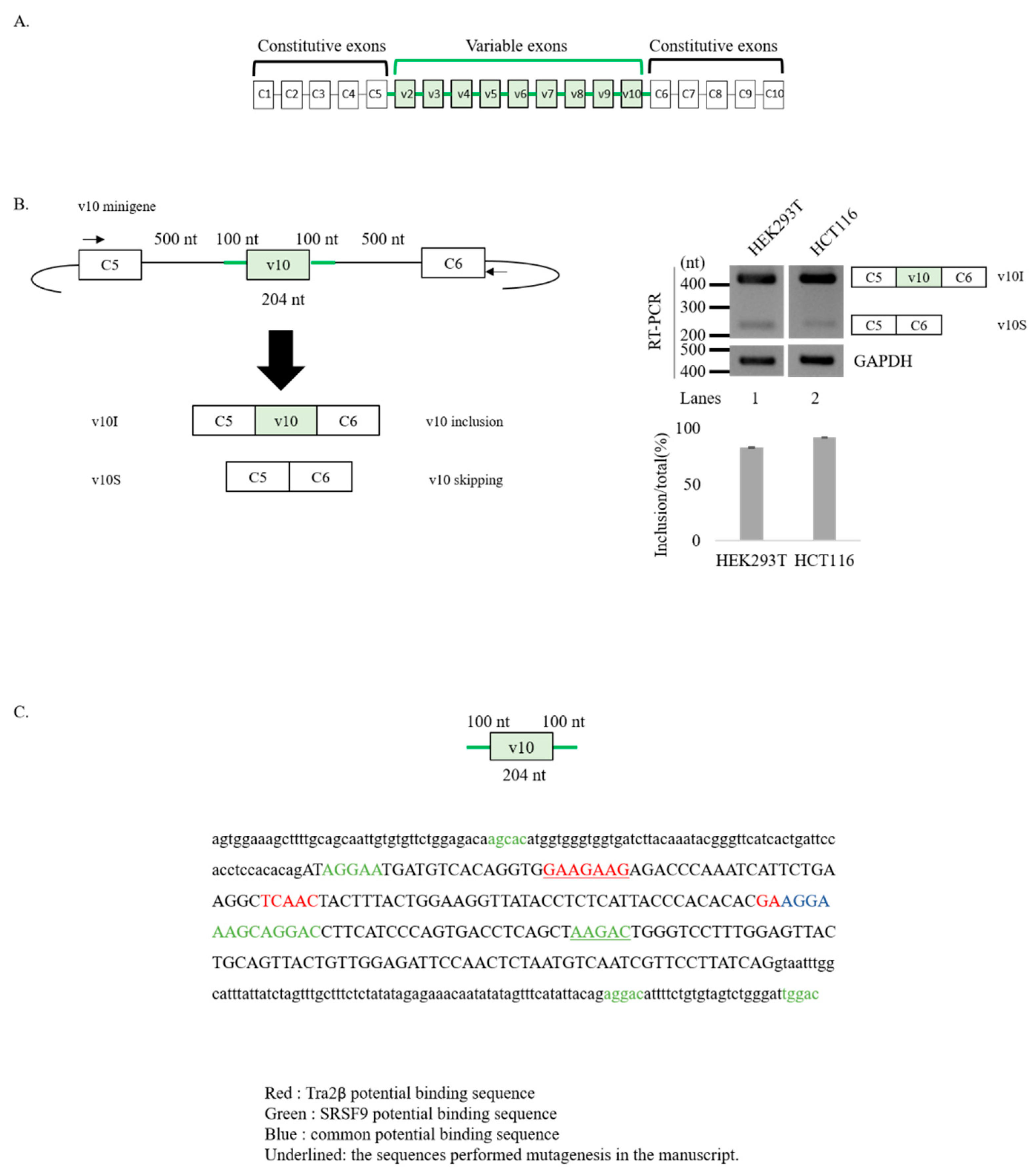
3.1. v10 Exon Contains Binding Motifs of Tra2β and SRSF9
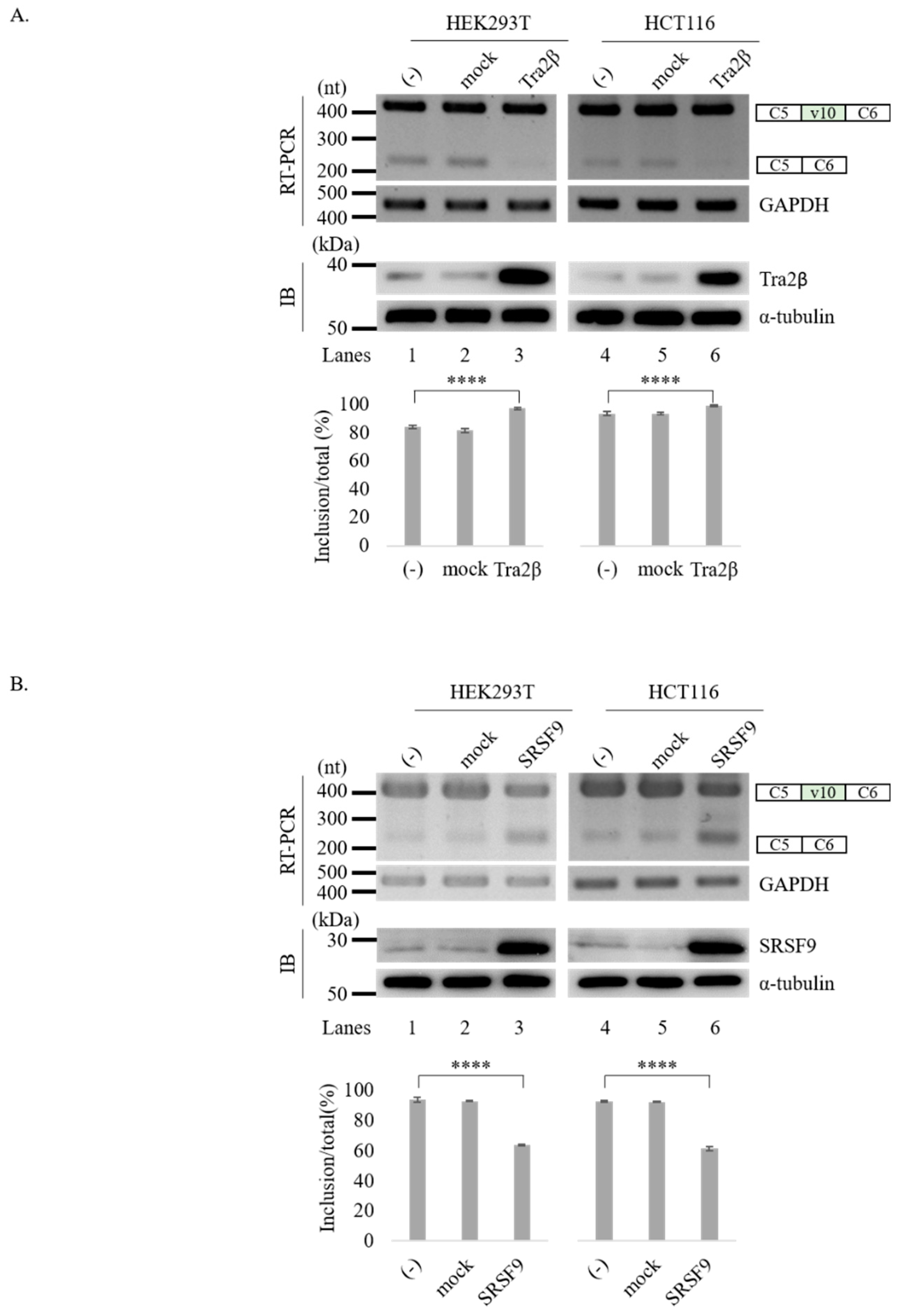
3.2. Tra2β and SRSF9 Regulate CD44 v10 Exon Splicing in Opposite Directions
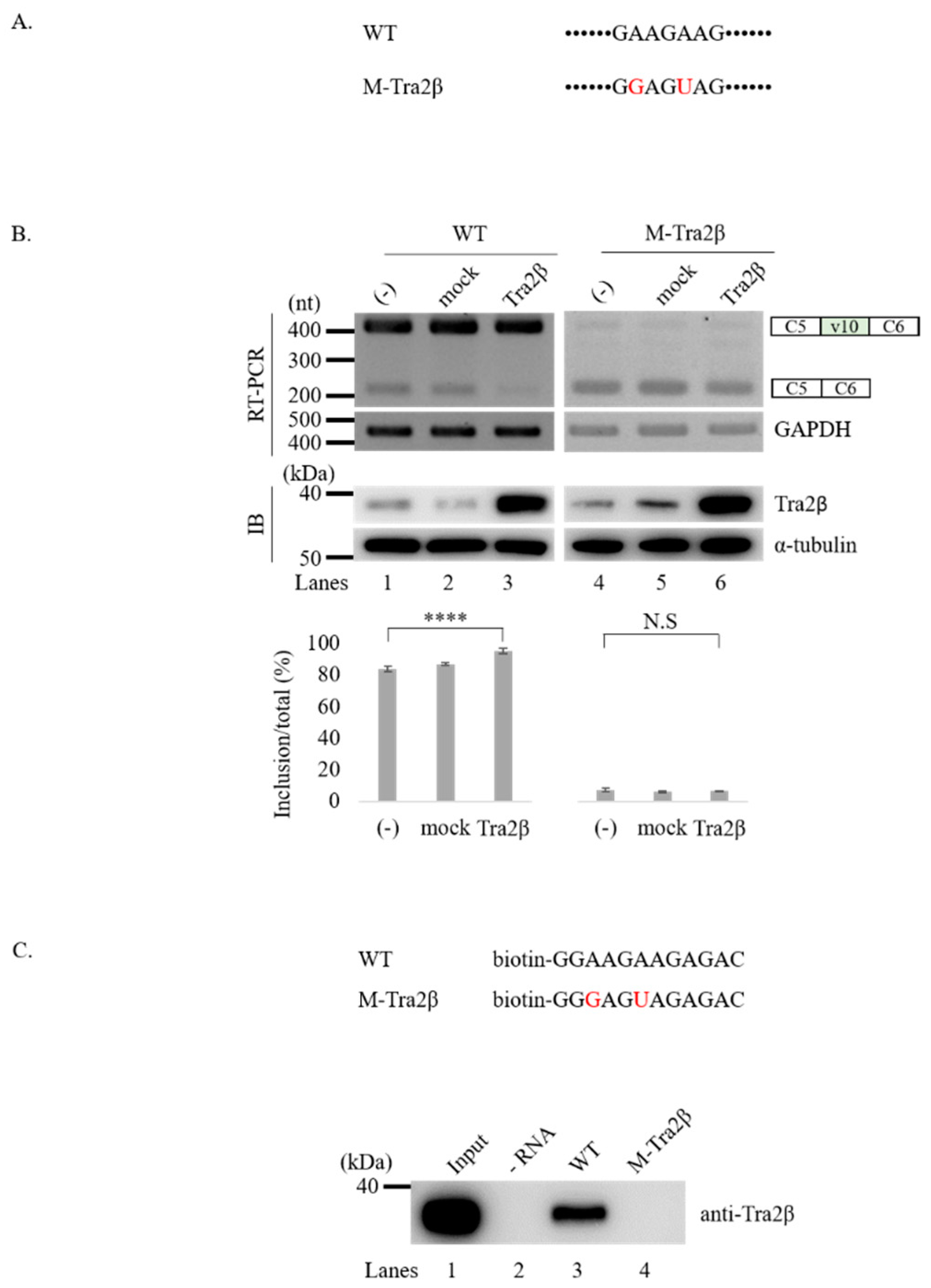
3.3. Tra2β Functionally Targets GAAGAAG Sequence in v10 Exon to Regulate Splicing of v10 Exon
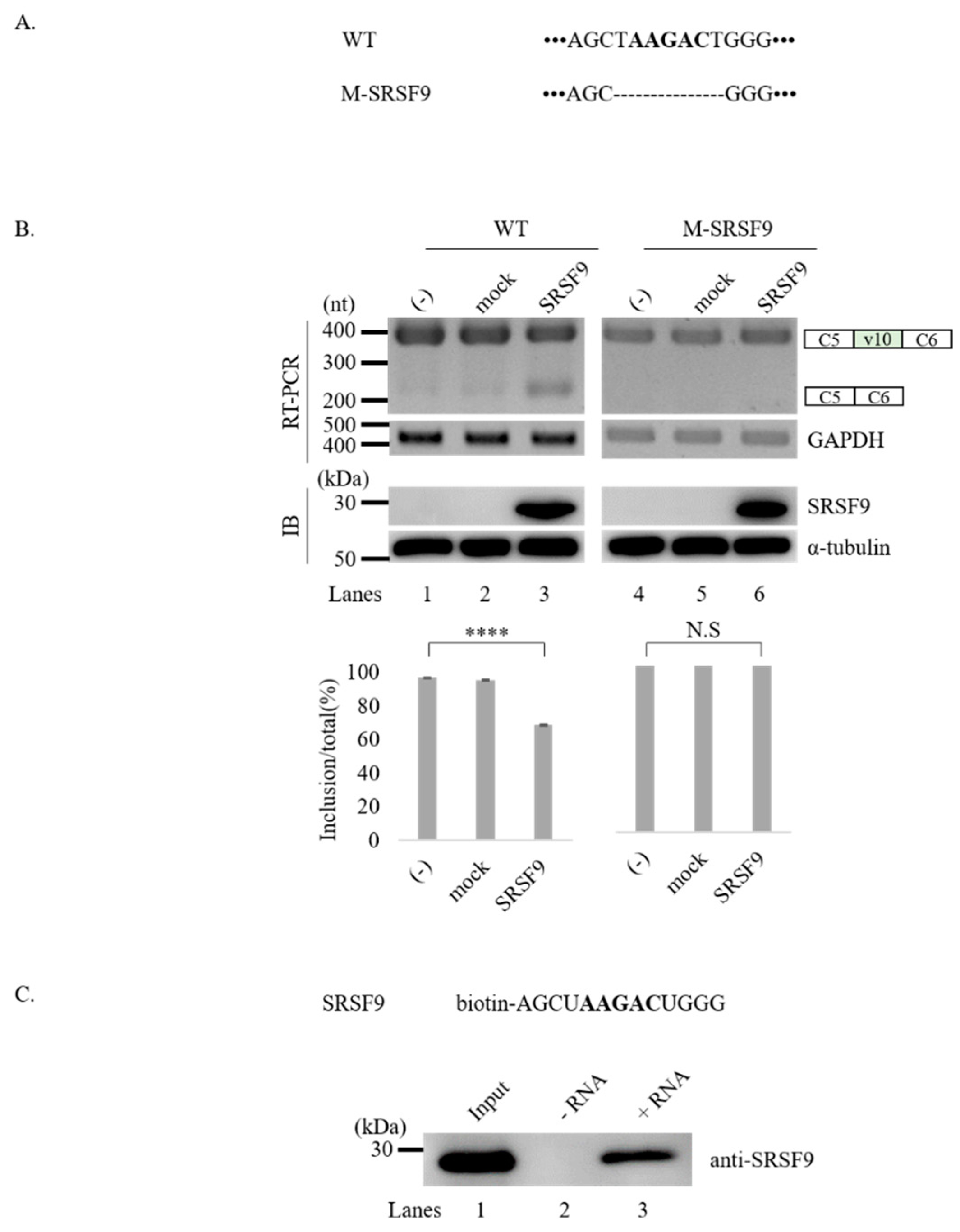
3.4. SRSF9 Targets AAGAC Sequence in v10 Exon to Inhibit v10 Exon Inclusion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Green, M.R. Pre-mRNA splicing. Annu. Rev. Genet. 1986, 20, 671–708. [Google Scholar] [CrossRef] [PubMed]
- Maniatis, T.; Reed, R. The role of small nuclear ribonucleoprotein particles in pre-mRNA splicing. Nature 1987, 325, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Bindereif, A.; Green, M.R. Identification and functional analysis of mammalian splicing factors. Genet. Eng. 1990, 12, 201–224. [Google Scholar] [CrossRef]
- Blencowe, B.J. Alternative splicing: New insights from global analyses. Cell 2006, 126, 37–47. [Google Scholar] [CrossRef]
- Bonnal, S.C.; Lopez-Oreja, I.; Valcarcel, J. Roles and mechanisms of alternative splicing in cancer—Implications for care. Nat. Rev. Clin. Oncol. 2020, 17, 457–474. [Google Scholar] [CrossRef]
- Black, D.L. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem. 2003, 72, 291–336. [Google Scholar] [CrossRef] [PubMed]
- Anczukow, O.; Krainer, A.R. Splicing-factor alterations in cancers. RNA 2016, 22, 1285–1301. [Google Scholar] [CrossRef]
- Biamonti, G.; Catillo, M.; Pignataro, D.; Montecucco, A.; Ghigna, C. The alternative splicing side of cancer. Semin. Cell Dev. Biol. 2014, 32, 30–36. [Google Scholar] [CrossRef]
- Munkley, J.; Livermore, K.; Rajan, P.; Elliott, D.J. RNA splicing and splicing regulator changes in prostate cancer pathology. Hum. Genet. 2017, 136, 1143–1154. [Google Scholar] [CrossRef]
- Gronthos, S.; Franklin, D.M.; Leddy, H.A.; Robey, P.G.; Storms, R.W.; Gimble, J.M. Surface protein characterization of human adipose tissue-derived stromal cells. J. Cell Physiol. 2001, 189, 54–63. [Google Scholar] [CrossRef]
- Prochazka, L.; Tesarik, R.; Turanek, J. Regulation of alternative splicing of CD44 in cancer. Cell Signal. 2014, 26, 2234–2239. [Google Scholar] [CrossRef]
- Yamano, S.; Gi, M.; Tago, Y.; Doi, K.; Okada, S.; Hirayama, Y.; Tachibana, H.; Ishii, N.; Fujioka, M.; Tatsumi, K.; et al. Role of deltaNp63(pos)CD44v(pos) cells in the development of N-nitroso-tris-chloroethylurea-induced peripheral-type mouse lung squamous cell carcinomas. Cancer Sci. 2016, 107, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Pradella, D.; Naro, C.; Sette, C.; Ghigna, C. EMT and stemness: Flexible processes tuned by alternative splicing in development and cancer progression. Mol. Cancer 2017, 16, 8. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.A.; Guo, W.; Liao, M.J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef]
- Brown, R.L.; Reinke, L.M.; Damerow, M.S.; Perez, D.; Chodosh, L.A.; Yang, J.; Cheng, C. CD44 splice isoform switching in human and mouse epithelium is essential for epithelial-mesenchymal transition and breast cancer progression. J. Clin. Investig. 2011, 121, 1064–1074. [Google Scholar] [CrossRef] [PubMed]
- Yae, T.; Tsuchihashi, K.; Ishimoto, T.; Motohara, T.; Yoshikawa, M.; Yoshida, G.J.; Wada, T.; Masuko, T.; Mogushi, K.; Tanaka, H.; et al. Alternative splicing of CD44 mRNA by ESRP1 enhances lung colonization of metastatic cancer cell. Nat. Commun. 2012, 3, 883. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.; Oh, S.; Shin, I. Ablation of CD44 induces glycolysis-to-oxidative phosphorylation transition via modulation of the c-Src-Akt-LKB1-AMPKalpha pathway. Biochem. J. 2016, 473, 3013–3030. [Google Scholar] [CrossRef]
- Bellerby, R.; Smith, C.; Kyme, S.; Gee, J.; Gunthert, U.; Green, A.; Rakha, E.; Barrett-Lee, P.; Hiscox, S. Overexpression of Specific CD44 Isoforms Is Associated with Aggressive Cell Features in Acquired Endocrine Resistance. Front. Oncol. 2016, 6, 145. [Google Scholar] [CrossRef]
- Kan, M.; Aki, M.; Akiyama, K.; Naruo, S.; Kanayama, H.; Kagawa, S. High-level expression of the CD44 variant sharing exon v10 in renal cancer. Jpn. J. Cancer Res. 1995, 86, 847–853. [Google Scholar] [CrossRef]
- Iida, N.; Bourguignon, L.Y. New CD44 splice variants associated with human breast cancers. J. Cell Physiol. 1995, 162, 127–133. [Google Scholar] [CrossRef]
- Freyschmidt-Paul, P.; Seiter, S.; Zoller, M.; Konig, A.; Ziegler, A.; Sundberg, J.P.; Happle, R.; Hoffmann, R. Treatment with an anti-CD44v10-specific antibody inhibits the onset of alopecia areata in C3H/HeJ mice. J. Investig. Dermatol. 2000, 115, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Rosel, M.; Khaldoyanidi, S.; Zawadzki, V.; Zoller, M. Involvement of CD44 variant isoform v10 in progenitor cell adhesion and maturation. Exp. Hematol. 1999, 27, 698–711. [Google Scholar] [CrossRef]
- Zoller, M.; Gupta, P.; Marhaba, R.; Vitacolonna, M.; Freyschmidt-Paul, P. Anti-CD44-mediated blockade of leukocyte migration in skin-associated immune diseases. J. Leukoc. Biol. 2007, 82, 57–71. [Google Scholar] [CrossRef]
- Iida, J.; Clancy, R.; Dorchak, J.; Somiari, R.I.; Somiari, S.; Cutler, M.L.; Mural, R.J.; Shriver, C.D. DNA aptamers against exon v10 of CD44 inhibit breast cancer cell migration. PLoS ONE 2014, 9, e88712. [Google Scholar] [CrossRef] [PubMed]
- Loh, T.J.; Cho, S.; Moon, H.; Jang, H.N.; Williams, D.R.; Jung, D.W.; Kim, I.C.; Ghigna, C.; Biamonti, G.; Zheng, X.; et al. hnRNP L inhibits CD44 V10 exon splicing through interacting with its upstream intron. Biochim. Biophys. Acta 2015, 1849, 743–750. [Google Scholar] [CrossRef]
- Graveley, B.R. Sorting out the complexity of SR protein functions. RNA 2000, 6, 1197–1211. [Google Scholar] [CrossRef] [PubMed]
- Long, J.C.; Caceres, J.F. The SR protein family of splicing factors: Master regulators of gene expression. Biochem. J. 2009, 417, 15–27. [Google Scholar] [CrossRef]
- Hoshijima, K.; Inoue, K.; Higuchi, I.; Sakamoto, H.; Shimura, Y. Control of doublesex alternative splicing by transformer and transformer-2 in Drosophila. Science 1991, 252, 833–836. [Google Scholar] [CrossRef]
- Belote, J.M.; Baker, B.S. The dual functions of a sex determination gene in Drosophila melanogaster. Dev. Biol. 1983, 95, 512–517. [Google Scholar] [CrossRef]
- Grellscheid, S.N.; Dalgliesh, C.; Rozanska, A.; Grellscheid, D.; Bourgeois, C.F.; Stevenin, J.; Elliott, D.J. Molecular design of a splicing switch responsive to the RNA binding protein Tra2beta. Nucleic Acids Res. 2011, 39, 8092–8104. [Google Scholar] [CrossRef]
- Venables, J.P.; Bourgeois, C.F.; Dalgliesh, C.; Kister, L.; Stevenin, J.; Elliott, D.J. Up-regulation of the ubiquitous alternative splicing factor Tra2beta causes inclusion of a germ cell-specific exon. Hum. Mol. Genet. 2005, 14, 2289–2303. [Google Scholar] [CrossRef] [PubMed]
- Lareau, L.F.; Inada, M.; Green, R.E.; Wengrod, J.C.; Brenner, S.E. Unproductive splicing of SR genes associated with highly conserved and ultraconserved DNA elements. Nature 2007, 446, 926–929. [Google Scholar] [CrossRef]
- Tsuda, K.; Someya, T.; Kuwasako, K.; Takahashi, M.; He, F.; Unzai, S.; Inoue, M.; Harada, T.; Watanabe, S.; Terada, T.; et al. Structural basis for the dual RNA-recognition modes of human Tra2-beta RRM. Nucleic Acids Res. 2011, 39, 1538–1553. [Google Scholar] [CrossRef]
- Clery, A.; Jayne, S.; Benderska, N.; Dominguez, C.; Stamm, S.; Allain, F.H. Molecular basis of purine-rich RNA recognition by the human SR-like protein Tra2-beta1. Nat. Struct. Mol. Biol. 2011, 18, 443–450. [Google Scholar] [CrossRef]
- Park, S.; Brugiolo, M.; Akerman, M.; Das, S.; Urbanski, L.; Geier, A.; Kesarwani, A.K.; Fan, M.; Leclair, N.; Lin, K.T.; et al. Differential Functions of Splicing Factors in Mammary Transformation and Breast Cancer Metastasis. Cell Rep. 2019, 29, 2672–2688. [Google Scholar] [CrossRef]
- Cloutier, P.; Toutant, J.; Shkreta, L.; Goekjian, S.; Revil, T.; Chabot, B. Antagonistic effects of the SRp30c protein and cryptic 5′ splice sites on the alternative splicing of the apoptotic regulator Bcl-x. J. Biol Chem 2008, 283, 21315–21324. [Google Scholar] [CrossRef]
- Simard, M.J.; Chabot, B. SRp30c is a repressor of 3’ splice site utilization. Mol. Cell Biol. 2002, 22, 4001–4010. [Google Scholar] [CrossRef]
- Young, P.J.; DiDonato, C.J.; Hu, D.; Kothary, R.; Androphy, E.J.; Lorson, C.L. SRp30c-dependent stimulation of survival motor neuron (SMN) exon 7 inclusion is facilitated by a direct interaction with hTra2 beta 1. Hum. Mol. Genet. 2002, 11, 577–587. [Google Scholar] [CrossRef]
- Fu, Y.; Huang, B.; Shi, Z.; Han, J.; Wang, Y.; Huangfu, J.; Wu, W. SRSF1 and SRSF9 RNA binding proteins promote Wnt signalling-mediated tumorigenesis by enhancing beta-catenin biosynthesis. EMBO Mol. Med. 2013, 5, 737–750. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.N.; Lee, M.; Loh, T.J.; Choi, S.W.; Oh, H.K.; Moon, H.; Cho, S.; Hong, S.E.; Kim, D.H.; Sheng, Z.; et al. Exon 9 skipping of apoptotic caspase-2 pre-mRNA is promoted by SRSF3 through interaction with exon 8. Biochim. Biophys. Acta 2014, 1839, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Piva, F.; Giulietti, M.; Burini, A.B.; Principato, G. SpliceAid 2: A database of human splicing factors expression data and RNA target motifs. Hum. Mutat. 2012, 33, 81–85. [Google Scholar] [CrossRef]
- Paz, I.; Kosti, I.; Ares, M., Jr.; Cline, M.; Mandel-Gutfreund, Y. RBPmap: A web server for mapping binding sites of RNA-binding proteins. Nucleic Acids Res. 2014, 42, W361–W367. [Google Scholar] [CrossRef] [PubMed]
- Screaton, G.R.; Bell, M.V.; Bell, J.I.; Jackson, D.G. The identification of a new alternative exon with highly restricted tissue expression in transcripts encoding the mouse Pgp-1 (CD44) homing receptor. Comparison of all 10 variable exons between mouse, human, and rat. J. Biol. Chem. 1993, 268, 12235–12238. [Google Scholar] [PubMed]
- Loh, T.J.; Moon, H.; Cho, S.; Jung, D.W.; Hong, S.E.; Kim, D.H.; Green, M.R.; Zheng, X.; Zhou, J.; Shen, H. SC35 promotes splicing of the C5-V6-C6 isoform of CD44 pre-mRNA. Oncol. Rep. 2014, 31, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Bian, T.; Feng, J.; Qian, L.; Zhang, J.; Jiang, D.; Zhang, Q.; Li, X.; Liu, Y.; Shi, J. miR-335 inhibited cell proliferation of lung cancer cells by target Tra2beta. Cancer Sci. 2018, 109, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Sharp, P.A. Regulation of CD44 alternative splicing by SRm160 and its potential role in tumor cell invasion. Mol. Cell Biol. 2006, 26, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Gao, X.D.; Lee, J.H.; Huang, H.; Tan, H.; Ahn, J.; Reinke, L.M.; Peter, M.E.; Feng, Y.; Gius, D.; et al. Cell type-restricted activity of hnRNPM promotes breast cancer metastasis via regulating alternative splicing. Genes Dev. 2014, 28, 1191–1203. [Google Scholar] [CrossRef]
- Zhang, H.; Brown, R.L.; Wei, Y.; Zhao, P.; Liu, S.; Liu, X.; Deng, Y.; Hu, X.; Zhang, J.; Gao, X.D.; et al. CD44 splice isoform switching determines breast cancer stem cell state. Genes Dev. 2019, 33, 166–179. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, J.; Liu, Y.; Choi, N.; Ha, J.; Pradella, D.; Ghigna, C.; Zheng, X.; Shen, H. Opposite Roles of Tra2β and SRSF9 in the v10 Exon Splicing of CD44. Cancers 2020, 12, 3195. https://doi.org/10.3390/cancers12113195
Oh J, Liu Y, Choi N, Ha J, Pradella D, Ghigna C, Zheng X, Shen H. Opposite Roles of Tra2β and SRSF9 in the v10 Exon Splicing of CD44. Cancers. 2020; 12(11):3195. https://doi.org/10.3390/cancers12113195
Chicago/Turabian StyleOh, Jagyeong, Yongchao Liu, Namjeong Choi, Jiyeon Ha, Davide Pradella, Claudia Ghigna, Xuexiu Zheng, and Haihong Shen. 2020. "Opposite Roles of Tra2β and SRSF9 in the v10 Exon Splicing of CD44" Cancers 12, no. 11: 3195. https://doi.org/10.3390/cancers12113195
APA StyleOh, J., Liu, Y., Choi, N., Ha, J., Pradella, D., Ghigna, C., Zheng, X., & Shen, H. (2020). Opposite Roles of Tra2β and SRSF9 in the v10 Exon Splicing of CD44. Cancers, 12(11), 3195. https://doi.org/10.3390/cancers12113195