Cancer Cell Lines Are Useful Model Systems for Medical Research



Abstract
1. Introduction
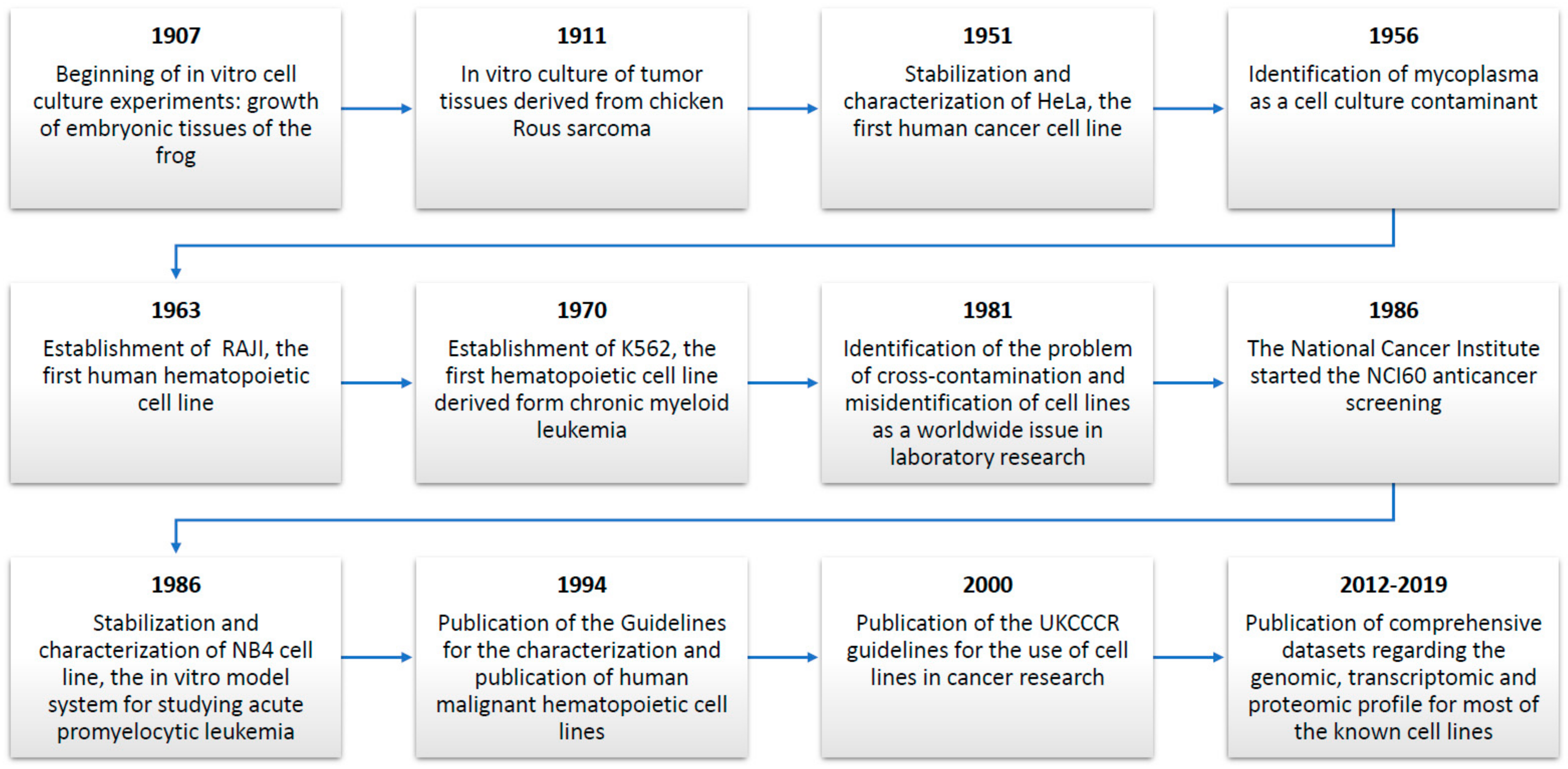
2. The Historical Progress of Cell Lines: Important Breakthroughs for Medical Research
3. Cell Lines in Modern Cancer Research: Toward the “Encyclopedia” of Cell Lines
4. Pitfalls in the Use of Cell Lines: Cross-Contamination and Mycoplasma Infection
5. Selection of the Most Appropriate Model System: The Case of Hematopoietic Cell Lines
6. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Masters, J.R.W. Human cancer cell lines: Fact and fantasy. Nat. Rev. Mol. Cell Biol. 2000, 1, 233–236. [Google Scholar] [CrossRef]
- Neve, R.M.; Chin, K.; Fridlyand, J.; Yeh, J.; Baehner, F.L.; Fevr, T.; Clark, L.; Bayani, N.; Coppe, J.P.; Tong, F.; et al. A collection of breast cancer cell lines for the study of functionally distinct cancer subtypes. Cancer Cell 2006, 10, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Matsuo, Y.; MacLeod, R.A.F. Continuous hematopoietic cell lines as model systems for leukemia-lymphoma research. Leuk. Res. 2000, 24, 881–911. [Google Scholar] [CrossRef]
- Asfari, M.; Janjic, D.; Meda, P.; Li, G.D.; Halban, P.A.; Wollheim, C.B. Establishment Of 2-mercaptoethanol-dependent differentiated insulin-secreting cell-lines. Endocrinology 1992, 130, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Sramkoski, R.M.; Pretlow, T.G.; Giaconia, J.M.; Pretlow, T.P.; Schwartz, S.; Sy, M.S.; Marengo, S.R.; Rhim, J.S.; Zhang, D.S.; Jacobberger, J.W. A new human prostate carcinoma cell line, 22R upsilon 1. In Vitro Cell. Dev. Biol. Animal 1999, 35, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, N.; Nakauchi, H.; Miwa, A.; Ishihara, T.; Eguchi, M.; Moroi, M.; Okada, M.; Sato, Y.; Wada, H.; Yawata, Y.; et al. Establishment And characterization of a human leukemic-cell line with megakaryocytic features–dependency on granulocyte-macrophage colony-stimulating factor, interleukin-3, or erythropoietin for growth and survival. Cancer Res. 1991, 51, 341–348. [Google Scholar]
- Sudo, H.; Kodama, H.A.; Amagai, Y.; Yamamoto, S.; Kasai, S. In vitro differentiation and calcification in a new clonal osteogenic cell-line derived from newborn mouse calvaria. J. Cell Biol. 1983, 96, 191–198. [Google Scholar] [CrossRef]
- Drexler, H.G.; Matsuo, Y. Guidelines for the characterization and publication of human malignant hematopoietic cell lines. Leukemia 1999, 13, 835–842. [Google Scholar] [CrossRef][Green Version]
- Matsuo, Y.; Drexler, H.G. Establishment and characterization of human B cell precursor leukemia cell lines. Leuk. Res. 1998, 22, 567–579. [Google Scholar] [CrossRef]
- Geraghty, R.J.; Capes-Davis, A.; Davis, J.M.; Downward, J.; Freshney, R.I.; Knezevic, I.; Lovell-Badge, R.; Masters, J.R.W.; Meredith, J.; Stacey, G.N.; et al. Guidelines for the use of cell lines in biomedical research. Br. J. Cancer 2014, 111, 1021–1046. [Google Scholar] [CrossRef]
- Monks, A.; Scudiero, D.; Skehan, P.; Shoemaker, R.; Paull, K.; Vistica, D.; Hose, C.; Langley, J.; Cronise, P.; Vaigrowolff, A.; et al. Feasibility of a high-flux anticancer drug screen using a diverse panel of cultured human tumor-cell lines. J. Natl. Cancer Inst. 1991, 83, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Rockwell, S. In vivo-in vitro tumor-cell lines—Characteristics and limitations as models for human cancer. Br. J. Cancer 1980, 41, 118–122. [Google Scholar]
- Goodspeed, A.; Heiser, L.M.; Gray, J.W.; Costello, J.C. Tumor-Derived Cell Lines as Molecular Models of Cancer Pharmacogenomics. Mol. Cancer Res. 2016, 14, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Barretina, J.; Caponigro, G.; Stransky, N.; Venkatesan, K.; Margolin, A.A.; Kim, S.; Wilson, C.J.; Lehar, J.; Kryukov, G.V.; Sonkin, D.; et al. The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature 2012, 483, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Garnett, M.J.; Edelman, E.J.; Heidorn, S.J.; Greenman, C.D.; Dastur, A.; Lau, K.W.; Greninger, P.; Thompson, I.R.; Luo, X.; Soares, J.; et al. Systematic identification of genomic markers of drug sensitivity in cancer cells. Nature 2012, 483, U570–U587. [Google Scholar] [CrossRef] [PubMed]
- Klijn, C.; Durinck, S.; Stawiski, E.W.; Haverty, P.M.; Jiang, Z.; Liu, H.; Degenhardt, J.; Mayba, O.; Gnad, F.; Liu, J.; et al. A comprehensive transcriptional portrait of human cancer cell lines. Nat. Biotechnol. 2015, 33, 306. [Google Scholar] [CrossRef]
- Li, J.; Zhao, W.; Akbani, R.; Liu, W.; Ju, Z.; Ling, S.; Vellano, C.P.; Roebuck, P.; Yu, Q.; Eterovic, A.K.; et al. Characterization of Human Cancer Cell Lines by Reverse-phase Protein Arrays. Cancer Cell 2017, 31, 225–239. [Google Scholar] [CrossRef]
- Huang, Y.H.; Vakoc, C.R. A Biomarker Harvest from One Thousand Cancer Cell Lines. Cell 2016, 166, 536–537. [Google Scholar] [CrossRef][Green Version]
- Capes-Davis, A.; Theodosopoulos, G.; Atkin, I.; Drexler, H.G.; Kohara, A.; MacLeod, R.A.F.; Masters, J.R.; Nakamura, Y.; Reid, Y.A.; Reddel, R.R.; et al. Check your cultures! A list of cross-contaminated or misidentified cell lines. Int. J. Cancer 2010, 127, 1–8. [Google Scholar] [CrossRef]
- Coppola, L.; Cianflone, A.; Grimaldi, A.M.; Incoronato, M.; Bevilacqua, P.; Messina, F.; Baselice, S.; Soricelli, A.; Mirabelli, P.; Salvatore, M. Biobanking in health care: Evolution and future directions. J. Transl. Med. 2019, 17, 172. [Google Scholar] [CrossRef]
- Harrison, R.G. Embryonic transplantation and development of the nervous system. Anatom. Rec. 1908, 2, 385–410. [Google Scholar] [CrossRef]
- Carrel, A.; Burrows, M.T. Cultivation in vitro of malignant tumors. J. Exp. Med. 1911, 13, 571–575. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Scherer, W.F.; Syverton, J.T.; Gey, G.O. Studies on the propagation in vitro of poliomyelitis viruses. IV. Viral multiplication in a stable strain of human malignant epithelial cells (strain HeLa) derived from an epidermoid carcinoma of the cervix. J. Exp. Med. 1953, 97, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Lucey, B.P.; Nelson-Rees, W.A.; Hutchins, G.M. Henrietta Lacks, HeLa Cells, and Cell Culture Contamination. Arch. Pathol. Lab. Med. 2009, 133, 1463–1467. [Google Scholar] [PubMed]
- Masters, J.R. HeLa cells 50 years on: The good, the bad and the ugly. Nat. Rev. Cancer 2002, 2, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Pulvertaft, J.V. Cytology Of Burkitt’s Tumour (African Lymphoma). Lancet 1964, 1, 238–240. [Google Scholar] [CrossRef]
- Drexler, H.G.; MacLeod, R.A.F.; Uphoff, C.C. Leukemia cell lines: In vitro models for the study of Philadelphia chromosome-positive leukemia. Leuk. Res. 1999, 23, 207–215. [Google Scholar]
- Lanotte, M.; Martin-Thouvenin, V.; Najman, S.; Balerini, P.; Valensi, F.; Berger, R. NB4, a maturation inducible cell line with t(15;17) marker isolated from a human acute promyelocytic leukemia (M3). Blood 1991, 77, 1080–1086. [Google Scholar]
- Falini, B.; Flenghi, L.; Fagioli, M.; Lo Coco, F.; Cordone, I.; Diverio, D.; Pasqualucci, L.; Biondi, A.; Riganelli, D.; Orleth, A.; et al. Immunocytochemical diagnosis of acute promyelocytic leukemia (M3) with the monoclonal antibody PG-M3 (anti-PML). Blood 1997, 90, 4046–4053. [Google Scholar]
- O’Dwyer, M.E.; Druker, B.J. STI571: An inhibitor of the BCR-ABL tyrosine kinase for the treatment of chronic myelogenous leukaemia. Lancet Oncol. 2000, 1, 207–211. [Google Scholar] [CrossRef]
- Gonzalez-Nicolini, V.; Fussenegger, M. In vitro assays for anticancer drug discovery—A novel approach based on engineered mammalian cell lines. Anticancer Drugs 2005, 16, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, R.H. The NCI60 human tumour cell line anticancer drug screen. Nat. Rev. Cancer 2006, 6, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, C.H. Anticancer drug development at the US National Cancer Institute. Cancer Chemother. Pharmacol. 2003, 52, S29–S33. [Google Scholar] [CrossRef] [PubMed]
- Oconnor, P.M.; Jackman, J.; Bae, I.; Myers, T.G.; Fan, S.J.; Mutoh, M.; Scudiero, D.A.; Monks, A.; Sausville, E.A.; Weinstein, J.N.; et al. Characterization of the p53 tumor suppressor pathway in cell lines of the National Cancer Institute anticancer drug screen and correlations with the growth-inhibitory potency of 123 anticancer agents. Cancer Res. 1997, 57, 4285–4300. [Google Scholar]
- Suggitt, M.; Bibby, M.C. 50 years of preclinical anticancer drug screening: Empirical to target-driven approaches. Clin. Cancer Res. 2005, 11, 971–981. [Google Scholar]
- Holbeck, S.L.; Collins, J.M.; Doroshow, J.H. Analysis of Food and Drug Administration-Approved Anticancer Agents in the NCI60 Panel of Human Tumor Cell Lines. Mol. Cancer Therapeut. 2010, 9, 1451–1460. [Google Scholar] [CrossRef]
- Close, D.A.; Wang, A.X.; Kochanek, S.J.; Shun, T.; Eiseman, J.L.; Johnston, P.A. Implementation of the NCI-60 Human Tumor Cell Line Panel to Screen 2260 Cancer Drug Combinations to Generate >3 Million Data Points Used to Populate a Large Matrix of Anti-Neoplastic Agent Combinations (ALMANAC) Database. Slas Disc. 2019, 24, 242–263. [Google Scholar] [CrossRef]
- Chabner, B.A.; Roberts, T.G. Timeline—Chemotherapy and the war on cancer. Nat. Rev. Cancer 2005, 5, 65–72. [Google Scholar] [CrossRef]
- Turner, T. Development of the Polio Vaccine: A Historical Perspective of Tuskegee University’s Role in Mass Production and Distribution of HeLa Cells. J. Health Care Poor Underser. 2012, 23, 5–10. [Google Scholar] [CrossRef]
- Longo, D.L. Imatinib Changed Everything. N. Engl. J. Med. 2017, 376, 982–983. [Google Scholar] [CrossRef]
- Cicconi, L.; Lo-Coco, F. Current management of newly diagnosed acute promyelocytic leukemia. Ann. Oncol. 2016, 27, 1474–1481. [Google Scholar] [CrossRef] [PubMed]
- Otto, R.; Sers, C.; Leser, U. Robust in-silico identification of cancer cell lines based on next generation sequencing. Oncotarget 2017, 8, 34310–34320. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-omics approaches to disease. Genome Biol. 2017, 18, e83. [Google Scholar] [CrossRef] [PubMed]
- Dan, S.; Tsunoda, T.; Kitahara, O.; Yanagawa, R.; Zembutsu, H.; Katagiri, T.; Yamazaki, K.; Nakamura, Y.; Yamori, T. An integrated database of chemosensitivity to 55 anticancer drugs and gene expression profiles of 39 human cancer cell lines. Cancer Res. 2002, 62, 1139–1147. [Google Scholar] [PubMed]
- Chen, W.-H.; Lu, G.; Chen, X.; Zhao, X.-M.; Bork, P. OGEE v2: An update of the online gene essentiality database with special focus on differentially essential genes in human cancer cell lines. Nucleic Acids Res. 2017, 45, D940–D944. [Google Scholar] [CrossRef] [PubMed]
- Pauli, C.; Hopkins, B.D.; Prandi, D.; Shaw, R.; Fedrizzi, T.; Sboner, A.; Sailer, V.; Augello, M.; Puca, L.; Rosati, R.; et al. Personalized In Vitro and In Vivo Cancer Models to Guide Precision Medicine. Cancer Discov. 2017, 7, 462–477. [Google Scholar] [CrossRef] [PubMed]
- Iorio, F.; Knijnenburg, T.A.; Vis, D.J.; Bignell, G.R.; Menden, M.P.; Schubert, M.; Aben, N.; Goncalves, E.; Barthorpe, S.; Lightfoot, H.; et al. A Landscape of Pharmacogenomic Interactions in Cancer. Cell 2016, 166, 740–754. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, J.N. Drug discovery Cell lines battle cancer. Nature 2012, 483, 544–545. [Google Scholar] [CrossRef]
- Sandberg, R.; Ernberg, I. Assessment of tumor characteristic gene expression in cell lines using a tissue similarity index (TSI). Proc. Nat. Acad. Sci. USA 2005, 102, 2052–2057. [Google Scholar] [CrossRef]
- Jin, H.; Barrott, J.J.; Cable, M.G.; Monument, M.J.; Lerman, D.M.; Smith-Fry, K.; Nollner, D.; Jones, K.B. The Impact of Microenvironment on the Synovial Sarcoma Transcriptome. Cancer Microenviron. 2017, 10, 1–7. [Google Scholar] [CrossRef]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.M.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M.; Canc Genome Atlas Res, N. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Filipp, F.V. Precision medicine driven by cancer systems biology. Cancer Metastasis Rev. 2017, 36, 91–108. [Google Scholar] [CrossRef] [PubMed]
- Tsherniak, A.; Vazquez, F.; Montgomery, P.G.; Weir, B.A.; Kryukov, G.; Cowley, G.S.; Gill, S.; Harrington, W.F.; Pantel, S.; Krill-Burger, J.M.; et al. Defining a Cancer Dependency Map. Cell 2017, 170, 564. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ning, S.; Ghandi, M.; Kryukov, G.V.; Gopal, S.; Deik, A.; Souza, A.; Pierce, K.; Keskula, P.; Hernandez, D.; et al. The landscape of cancer cell line metabolism. Nat. Med. 2019, 25, 850. [Google Scholar] [CrossRef] [PubMed]
- Ghandi, M.; Huang, F.W.; Jane-Valbuena, J.; Kryukov, G.V.; Lo, C.C.; McDonald, E.R., 3rd; Barretina, J.; Gelfand, E.T.; Bielski, C.M.; Li, H.; et al. Next-generation characterization of the Cancer Cell Line Encyclopedia. Nature 2019, 569, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Van der Meer, D.; Barthorpe, S.; Yang, W.; Lightfoot, H.; Hall, C.; Gilbert, J.; Francies, H.E.; Garnett, M.J. Cell Model Passportsa hub for clinical, genetic and functional datasets of preclinical cancer models. Nucleic Acids Res. 2019, 47, 923–929. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.F.; Mardis, E.R. The emerging clinical relevance of genomics in cancer medicine. Nat. Rev. Clin. Oncol. 2018, 15, 353–365. [Google Scholar] [CrossRef]
- Gemei, M.; Mirabelli, P.; Di Noto, R.; Fortunato, G.; Del Vecchio, L. Divergent expression of CD133 in different studies on HCT-116 cell line. Int. J. Cancer 2011, 128, 998–1000. [Google Scholar] [CrossRef]
- Mariotti, E.; Gemei, M.; Mirabelli, P.; D’Alessio, F.; Di Noto, R.; Fortunato, G.; Del Vecchio, L. The percentage of CD133+ cells in human colorectal cancer cell lines is influenced by Mycoplasma hyorhinis infection. BMC Cancer 2010, 10, e120. [Google Scholar] [CrossRef]
- Nelsonrees, W.A.; Daniels, D.W.; Flandermeyer, R.R. Cross-Contamination of cells in culture. Science 1981, 212, 446–452. [Google Scholar] [CrossRef]
- Nelson-Rees, W.A. Responsibility for truth in research. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2001, 356, 849–851. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Dirks, W.G.; Matsuo, Y.; MacLeod, R.A.F. False leukemia-lymphoma cell lines: An update on over 500 cell lines. Leukemia 2003, 17, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Uphoff, C.C.; Dirks, W.G.; MacLeod, R.A.F. Mix-ups and mycoplasma: The enemies within. Leuk. Res. 2002, 26, 329–333. [Google Scholar] [CrossRef]
- MacLeod, R.A.F.; Dirks, W.G.; Matsuo, Y.; Kaufmann, M.; Milch, H.; Drexler, H.G. Widespread intraspecies cross-contamination of human tumor cell lines arising at source. Int. J. Cancer 1999, 83, 555–563. [Google Scholar] [CrossRef]
- Freshney, R.I. Database of misidentified cell lines. Int. J. Cancer 2010, 126, 302. [Google Scholar] [CrossRef] [PubMed]
- Masters, J.R.; Thomson, J.A.; Daly-Burns, B.; Reid, Y.A.; Dirks, W.G.; Packer, P.; Toji, L.H.; Ohno, T.; Tanabe, H.; Arlett, C.F.; et al. Short tandem repeat profiling provides an international reference standard for human cell lines. Proc. Nat. Acad. Sci. USA 2001, 98, 8012–8017. [Google Scholar] [CrossRef] [PubMed]
- Babic, Z.; Capes-Davis, A.; Martone, M.E.; Bairoch, A.; Ozyurt, I.B.; Gillespie, T.H.; Bandrowski, A.E. Incidences of problematic cell lines are lower in papers that use RRIDs to identify cell lines. Elife 2019, 8, e41676. [Google Scholar] [CrossRef] [PubMed]
- Robinson, L.B.; Wichelhausen, R.H. Contamination of human cell cultures by pleuropneumonialike organisms. Science 1956, 124, 1147–1148. [Google Scholar] [CrossRef] [PubMed]
- Razin, S. The Genus Mycoplasma and Related Genera (Class Mollicutes). In Prokaryotes Volume 4 Bacteria Firmicutes Cyanobacteria; Springer: New York, NY, USA, 2006; pp. 836–904. [Google Scholar]
- Kong, F.; James, G.; Gordon, S.; Zelynski, A.; Gilbert, G.L. Species-specific PCR for identification of common contaminant mollicutes in cell culture. Appl. Environ. Microbiol. 2001, 67, 3195–3200. [Google Scholar] [CrossRef]
- Uphoff, C.C.; Denkmann, S.A.; Steube, K.G.; Drexler, H.G. Detection of EBV, HBV, HCV, HIV-1, HTLV-I and -II, and SMRV in Human and Other Primate Cell Lines. J. Biomed. Biotechnol. 2010, 2010, e904767. [Google Scholar] [CrossRef]
- Uphoff, C.C.; Drexler, H.G. Prevention of mycoplasma contamination in leukemia-lymphoma cell lines. Hum. Cell 2001, 14, 244–247. [Google Scholar]
- Coecke, S.; Balls, M.; Bowe, G.; Davis, J.; Gstraunthaler, G.; Hartung, T.; Hay, R.; Merten, O.W.; Price, A.; Schechtman, L.; et al. Guidance on Good Cell Culture Practice—A report of the second ECVAM task force on Good Cell Culture Practice. Atla Altern. Lab. Animals 2005, 33, 261–287. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G. Leukemia-cell lines—In-vitro models for the study of chronic myeloid-leukemia. Leuk. Res. 1994, 18, 919. [Google Scholar] [CrossRef]
- Drexler, H.G.; Quentmeier, H.; Macleod, R.A.F.; Uphoff, C.C.; Hu, Z.B. Leukemia-cell lines—In-vitro models for the study of acute promyelocytic leukemia. Leuk. Res. 1995, 19, 681–691. [Google Scholar] [CrossRef]
- Drexler, H.G.; Uphoff, C.C.; Gaidano, G.; Carbone, A. Lymphoma cell lines: In vitro models for the study of HHV-8+ primary effusion lymphomas (body cavity-based lymphomas). Leukemia 1998, 12, 1507–1517. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Matsuo, Y. Malignant hematopoietic cell lines: In vitro models for the study of multiple myeloma and plasma cell leukemia. Leuk. Res. 2000, 24, 681–703. [Google Scholar] [CrossRef]
- Drexler, H.G.; Quentmeier, H.; MacLeod, R.A.F. Malignant hematopoietic cell lines: In vitro models for the study of MLL gene alterations. Leukemia 2004, 18, 227–232. [Google Scholar] [CrossRef]
- Drexler, H.G.; Matsuo, Y.; MacLeod, R.A.F. Malignant hematopoietic cell lines: In vitro models for the study of erythroleukemia. Leuk. Res. 2004, 28, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; MacLeod, R.A.F. Malignant hematopoietic cell lines: In vitro models for the study of Waldenstrom’s macroglobulinemia. Leuk. Res. 2008, 32, 1669–1673. [Google Scholar] [CrossRef]
- Drexler, H.G.; Dirks, W.G.; MacLeod, R.A.F. Many are called MDS cell lines: One is chosen. Leuk. Res. 2009, 33, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Ehrentraut, S.; Nagel, S.; Eberth, S.; MacLeod, R.A.F. Malignant hematopoietic cell lines: In vitro models for the study of primary mediastinal B-cell lymphomas. Leuk. Res. 2015, 39, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Eberth, S.; Nagel, S.; MacLeod, R.A.F. Malignant hematopoietic cell lines: In vitro models for double-hit B-cell lymphomas. Leukemia Lymphoma 2016, 57, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Pommerenke, C.; Eberth, S.; Nagel, S. Hodgkin lymphoma cell lines: To separate the wheat from the chaff. Biol. Chem. 2018, 399, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Macleod, R.A.F. History of leukemia-lymphoma cell lines. Hum. Cell 2010, 23, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Quentmeier, H.; Pommerenke, C.; Dirks, W.G.; Eberth, S.; Koeppel, M.; MacLeod, R.A.F.; Nagel, S.; Steube, K.; Uphoff, C.C.; Drexler, H.G. The LL-100 panel: 100 cell lines for blood cancer studies. Sci. Rep. 2019, 9, e8218. [Google Scholar] [CrossRef] [PubMed]
- Beskow, L.M. Lessons from HeLa Cells: The Ethics and Policy of Biospecimens. Ann. Rev. Genom. Hum. Genet. 2016, 17, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Uphoff, C.C. Mycoplasma contamination of cell cultures: Incidence, sources, effects, detection, elimination, prevention. Cytotechnology 2002, 39, 75–90. [Google Scholar] [CrossRef]
- Osswald, A.; Hedrich, V.; Sommergruber, W. 3D-3 Tumor Models in Drug Discovery for Analysis of Immune Cell Infiltration. Methods Mol. Biol. 2019, 1953, 151–162. [Google Scholar] [CrossRef]
- Russo, V.; Paciocco, A.; Affinito, A.; Roscigno, G.; Fiore, D.; Palma, F.; Galasso, M.; Volinia, S.; Fiorelli, A.; Esposito, C.L.; et al. Aptamer-miR-34c Conjugate Affects Cell Proliferation of Non-Small-Cell Lung Cancer Cells. Mol. Ther. Nucleic Acids 2018, 13, 334–346. [Google Scholar] [CrossRef]
- Di Noto, R.; Luciano, L.; Lo Pardo, C.; Ferrara, F.; Frigeri, F.; Mercuro, O.; Lombardi, M.L.; Pane, F.; Vacca, C.; Manzo, C.; et al. JURL-MK1 (c-kit(high)/CD30(−)/CD40(−)) and JURL-MK2 (c-kit(low)/CD30(+)/CD40(+)) cell lines: ‘two-sided’ model for investigating leukemic megakaryocytopoiesis. Leukemia 1997, 11, 1554–1564. [Google Scholar] [CrossRef]
- Di Noto, R.; Pane, F.; Camera, A.; Luciano, L.; Barone, M.; Lo Pardo, C.; Boccuni, P.; Intrieri, M.; Izzo, B.; Villa, M.R.; et al. Characterization of two novel cell lines, DERL-2 (CD56(+)/CD3(+)/TCR gamma delta(+)) and DERL-7 (CD56(+)/CD3(−)/TCR gamma delta(−)), derived from a single patient with CD56(+) non-Hodgkin’s lymphoma. Leukemia 2001, 15, 1641–1649. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Di Noto, R.; Mirabelli, P.; Mariotti, E.; Sacchetti, L.; Pane, F.; Rotoli, B.; Lo Pardo, C.; Del Vecchio, L.; Mecucci, C.; Salvatore, F. CMRL-T, a novel T-cell line showing asynchronous phenotype (CD34+/CD1a−/TCRαβ+) and dual T-cell receptor β chain. Leukemia 2006, 20, 2175–2177. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Cell Line | Year of Stabilization | Number of Publications in the Cancer Field | Benefit for Public Health Care |
|---|---|---|---|
| HeLa | 1953 | 16,843 | Development of the anti-polio vaccine |
| RAJI | 1964 | 1557 | Definition of the mechanisms of infection by Epstein-Barr virus |
| K562 | 1976 | 8001 | Development of treatment protocols for chronic myeloid leukemia |
| NB4 | 1991 | 1227 | Development of treatment protocols for acute promyelocytic leukemia |
| Resource Name | Website | Description | Reference |
|---|---|---|---|
| Cancer Cell Line Encyclopedia | https://portals.broadinstitute.org/ccle | The Cancer Cell Line Encyclopedia (CCLE) database was conceived to conduct a detailed genetic and pharmacologic characterization of a large panel of human cancer models (approximately 110 models). Gene expression, mutation, methylation, RNAseq and metabolomics data are downloadable. | [14] |
| Genomics of Drug Sensitivity in Cancer | https://www.cancerrxgene.org/ | This project aims at screening >1000 genetically characterized human cancer cell lines with a wide range of anticancer therapeutics. The sensitivity patterns of the cell lines are correlated with extensive genomic data to identify genetic features that are predictive of sensitivity. | [47] |
| MD Anderson Cell Lines Project | https://tcpaportal.org/mclp/#/ | The MD Anderson Cell Lines Project depicts the expression levels of approximately 230 key cancer-related proteins in 650 independent cell lines. This bioinformatic resource is a comprehensive resource for accessing, visualizing, and analyzing functional proteomics of cancer cell lines. | [17] |
| Project Achilles | https://depmap.org/portal/achilles/ | Project Achilles systematically identifies and catalogs gene essentiality across hundreds of genomically characterized cancer cell lines. For each cell line, a list of genes able to alter cell survival is reported as a result of RNAi and/or CRISPR-Cas9 genetic silencing or knockout of the individual gene. Additionally, these results are linked to the genetic or molecular features of the tumors to provide a “cancer dependency map”. | [53] |
| Cell Model Passports | https://cellmodelpassports.sanger.ac.uk/ | This resource provides large-scale genomic datasets for approximately 1200 cancer cell line and organoid models cataloged. For each model system, it is possible to display associated somatic nucleotide variants, gene expression, copy number variations or methylation data. Its accessibility format is also useful for noncomputational, wet laboratory scientists. | [56] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirabelli, P.; Coppola, L.; Salvatore, M. Cancer Cell Lines Are Useful Model Systems for Medical Research. Cancers 2019, 11, 1098. https://doi.org/10.3390/cancers11081098
Mirabelli P, Coppola L, Salvatore M. Cancer Cell Lines Are Useful Model Systems for Medical Research. Cancers. 2019; 11(8):1098. https://doi.org/10.3390/cancers11081098
Chicago/Turabian StyleMirabelli, Peppino, Luigi Coppola, and Marco Salvatore. 2019. "Cancer Cell Lines Are Useful Model Systems for Medical Research" Cancers 11, no. 8: 1098. https://doi.org/10.3390/cancers11081098
APA StyleMirabelli, P., Coppola, L., & Salvatore, M. (2019). Cancer Cell Lines Are Useful Model Systems for Medical Research. Cancers, 11(8), 1098. https://doi.org/10.3390/cancers11081098