Human Tumor-Infiltrating Dendritic Cells: From In Situ Visualization to High-Dimensional Analyses

 ,
, 
Abstract
1. Introduction
2. DC Subpopulations and Their Functional Specificities
2.1. Plasmacytoid Dendritic Cells (pDCs) and Anti-Tumor Immunity
2.2. Conventional Dendritic Cells Subsets and Anti-Tumor Immunity
2.2.1. cDC1
2.2.2. cDC2
2.3. Other DC Populations
3. Detection of DCs in Solid Tumors
3.1. In situ Visualization of Tumor-Associated DCs
3.2. Identification of TA-DC Subsets by Flow Cytometry
4. High-Dimensional Technologies Applied to DC Subset Analyses
4.1. Limitations of Previous Techniques and Emergence of High-Dimensional Technologies
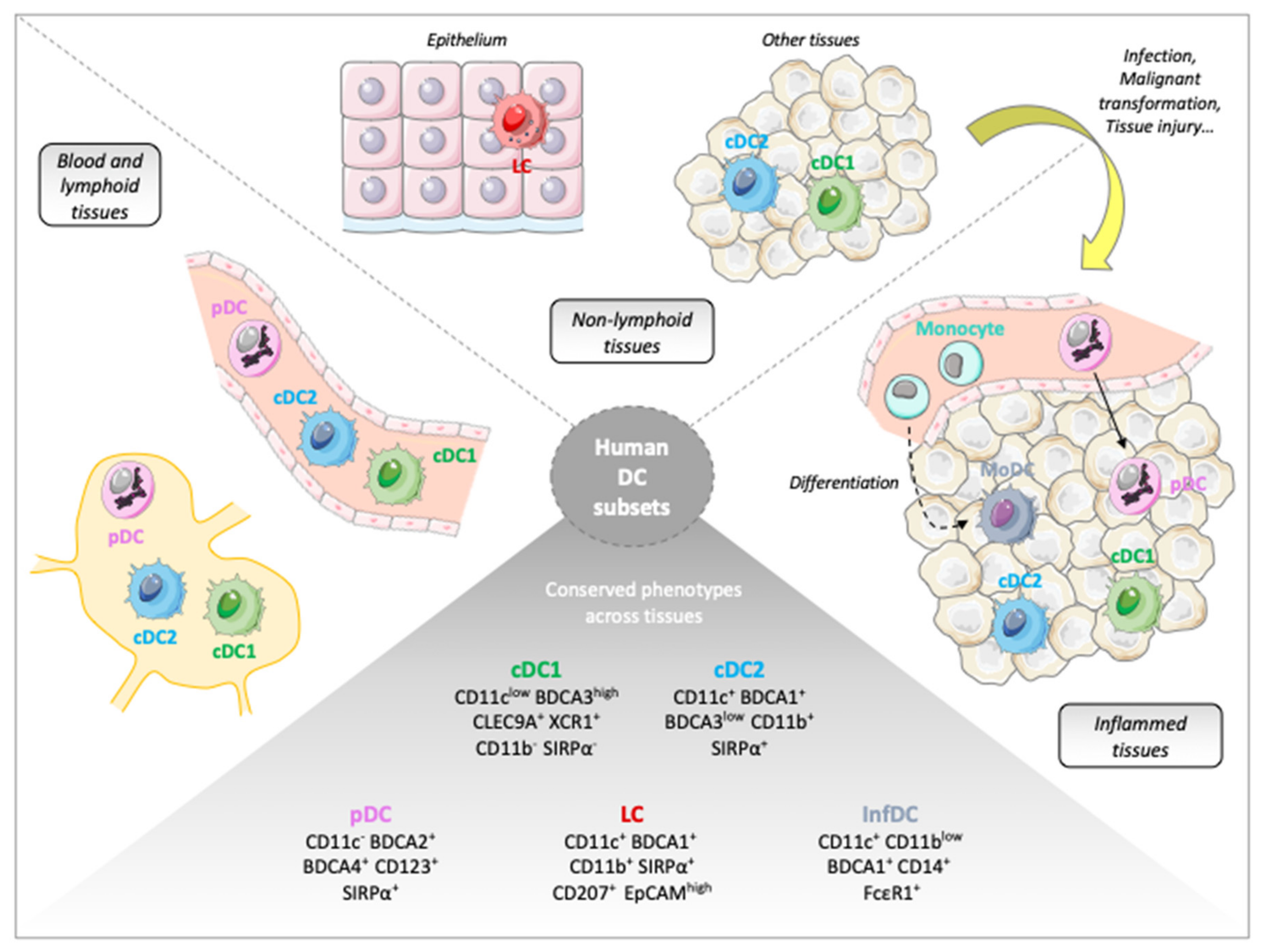
4.2. Inter-Tissue and Inter-Species Similarities
4.3. Redefining DC Classification
4.4. High-Dimensional Technologies Adapted to DC Subset Analysis in the Context of Tumors
4.5. Transcriptomic Signatures and Prognostic Impact of DCs on Cancer Patients
5. Perspectives for of TA-DC High-Dimensional Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Banchereau, J.; Briere, F.; Caux, C.; Davoust, J.; Lebecque, S.; Liu, Y.-J.; Pulendran, B.; Palucka, K. Immunobiology of Dendritic Cells. Annu. Rev. Immunol. 2000, 18, 767–811. [Google Scholar] [CrossRef] [PubMed]
- Dunn, G.P.; Old, L.J.; Schreiber, R.D. The Three Es of Cancer Immunoediting. Annu. Rev. Immunol. 2004, 22, 329360. [Google Scholar] [CrossRef]
- Chen, D.S.; Mellman, I. Oncology Meets Immunology: The Cancer-Immunity Cycle. Immunity 2013, 39, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.S.; Mellman, I. Elements of cancer immunity and the cancer–immune set point. Nature 2017, 541, 321. [Google Scholar] [CrossRef] [PubMed]
- Collin, M.; Bigley, V. Human dendritic cell subsets: An update. Immunology 2018, 154, 3–20. [Google Scholar] [CrossRef]
- Satpathy, A.T.; Wu, X.; Albring, J.C.; Murphy, K.M. Re(de)fining the dendritic cell lineage. Nat. Immunol. 2012, 13, 1145. [Google Scholar] [CrossRef] [PubMed]
- Vu Manh, T.-P.; Elhmouzi-Younes, J.; Urien, C.; Ruscanu, S.; Jouneau, L.; Bourge, M.; Moroldo, M.; Foucras, G.; Salmon, H.; Marty, H.; et al. Defining Mononuclear Phagocyte Subset Homology Across Several Distant Warm-Blooded Vertebrates Through Comparative Transcriptomics. Front. Immunol. 2015, 6, 299. [Google Scholar] [CrossRef]
- Dzionek, A.; Fuchs, A.; Schmidt, P.; Cremer, S.; Zysk, M.; Miltenyi, S.; Buck, D.; Schmitz, J. BDCA-2, BDCA-3, and BDCA-4: Three markers for distinct subsets of dendritic cells in human peripheral blood. J. Immunol. 2000, 165, 6037–6046. [Google Scholar] [CrossRef]
- MacDonald, K.P.; Munster, D.J.; Clark, G.J.; Dzionek, A.; Schmitz, J.; Hart, D.N. Characterization of human blood dendritic cell subsets. Blood 2002, 100, 45124520. [Google Scholar] [CrossRef]
- Belz, G.T.; Nutt, S.L. Transcriptional programming of the dendritic cell network. Nat. Rev. Immunol. 2012, 12, 101. [Google Scholar] [CrossRef]
- Guilliams, M.; Ginhoux, F.; Jakubzick, C.; Naik, S.H.; Onai, N.; Schraml, B.U.; Segura, E.; Tussiwand, R.; Yona, S. Dendritic cells, monocytes and macrophages: A unified nomenclature based on ontogeny. Nat. Rev. Immunol. 2014, 14, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Satpathy, A.T.; Murphy, K.M.; Wumesh, K. Transcription factor networks in dendritic cell development. Semin. Immunol. 2011, 23, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Watchmaker, P.B.; Lahl, K.; Lee, M.; Baumjohann, D.; Morton, J.; Kim, S.; Zeng, R.; Dent, A.; Ansel, M.K.; Diamond, B.; et al. Comparative transcriptional and functional profiling defines conserved programs of intestinal DC differentiation in humans and mice. Nat. Immunol. 2013, 15, ni.2768. [Google Scholar] [CrossRef] [PubMed]
- Guilliams, M.; Henri, S.; Tamoutounour, S.; Ardouin, L.; Schwartz-Cornil, I.; Dalod, M.; Malissen, B. From skin dendritic cells to a simplified classification of human and mouse dendritic cell subsets. Eur. J. Immunol. 2010, 40, 2089–2094. [Google Scholar] [CrossRef] [PubMed]
- Guilliams, M.; Dutertre, C.-A.A.; Scott, C.L.; McGovern, N.; Sichien, D.; Chakarov, S.; Gassen, S.; Chen, J.; Poidinger, M.; Prijck, S.; et al. Unsupervised High-Dimensional Analysis Aligns Dendritic Cells across Tissues and Species. Immunity 2016, 45, 669–684. [Google Scholar] [CrossRef] [PubMed]
- See, P.; Dutertre, C.-A.A.; Chen, J.; Günther, P.; McGovern, N.; Irac, S.E.; Gunawan, M.; Beyer, M.; Händler, K.; Duan, K.; et al. Mapping the human DC lineage through the integration of high-dimensional techniques. Science (N. Y.) 2017, 356, eaag3009. [Google Scholar] [CrossRef] [PubMed]
- Villani, A.-C.; Satija, R.; Reynolds, G.; Sarkizova, S.; Shekhar, K.; Fletcher, J.; Griesbeck, M.; Butler, A.; Zheng, S.; Lazo, S. Single-cell RNA-seq reveals new types of human blood dendritic cells, monocytes, and progenitors. Science 2017, 356, eaah4573. [Google Scholar] [CrossRef]
- Cella, M.; Facchetti, F.; Lanzavecchia, A.; Colonna, M. Plasmacytoid dendritic cells activated by influenza virus and CD40L drive a potent TH1 polarization. Nat. Immunol. 2000, 1, 305–310. [Google Scholar] [CrossRef]
- Kadowaki, N.; Antonenko, S.; Lau, J.; Liu, Y. Natural interferon alpha/beta-producing cells link innate and adaptive immunity. J. Exp. Med. 2000, 192, 219–226. [Google Scholar] [CrossRef]
- Villadangos, J.A.; Young, L. Antigen-Presentation Properties of Plasmacytoid Dendritic Cells. Immunity 2008, 29, 352–361. [Google Scholar] [CrossRef]
- Cella, M.; Jarrossay, D.; Facchetti, F.; Alebardi, O.; Nakajima, H.; Lanzavecchia, A.; Colonna, M. Plasmacytoid monocytes migrate to inflamed lymph nodes and produce large amounts of type I interferon. Nat. Med. 1999, 5, 919–923. [Google Scholar] [CrossRef] [PubMed]
- Siegal, F.P.; Kadowaki, N.; Shodell, M.; Fitzgerald-Bocarsly, P.A.; Shah, K.; Ho, S.; Antonenko, S.; Liu, Y.-J. The Nature of the Principal Type 1 Interferon-Producing Cells in Human Blood. Science 1999, 284, 18351837. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.; Liu, Y.-J. Regulation of TLR7/9 signaling in plasmacytoid dendritic cells. Protein Cell 2013, 4, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Yanai, H.; Negishi, H.; Asagiri, M.; Sato, M.; Mizutani, T.; Shimada, N.; Ohba, Y.; Takaoka, A.; Yoshida, N.; et al. IRF-7 is the master regulator of type-I interferon-dependent immune responses. Nature 2005, 434, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Ohba, Y.; Yanai, H.; Negishi, H.; Mizutani, T.; Takaoka, A.; Taya, C.; Taniguchi, T. Spatiotemporal regulation of MyD88–IRF-7 signalling for robust type-I interferon induction. Nature 2005, 434, 1035. [Google Scholar] [CrossRef] [PubMed]
- Guiducci, C.; Ott, G.; Chan, J.H.; Damon, E.; Calacsan, C.; Matray, T.; Lee, K.-D.; Coffman, R.L.; Barrat, F.J. Properties regulating the nature of the plasmacytoid dendritic cell response to Toll-like receptor 9 activation. J. Exp. Med. 2006, 203, 1999–2008. [Google Scholar] [CrossRef]
- Gresser, I.; Belardelli, F. Endogenous type I interferons as a defense against tumors. Cytokine Growth Factor Rev. 2002, 13, 111–118. [Google Scholar] [CrossRef]
- Zitvogel, L.; Galluzzi, L.; Kepp, O.; Smyth, M.J.; Kroemer, G. Type I interferons in anticancer immunity. Nat. Rev. Immunol. 2015, 15, nri3845. [Google Scholar] [CrossRef] [PubMed]
- Gresser, I.; Belardelli, F.; Maury, C.; Maunoury, M.; Tovey, M. Injection of mice with antibody to interferon enhances the growth of transplantable murine tumors. J. Exp. Med. 1983, 158, 2095–2107. [Google Scholar] [CrossRef]
- Deonarain, R.; Verma, A.; Porter, A.C.; Gewert, D.R.; Platanias, L.C.; Fish, E.N. Critical roles for IFN-β in lymphoid development, myelopoiesis, and tumor development: Links to tumor necrosis factor α. Proc Natl. Acad. Sci. USA 2003, 100, 13453–13458. [Google Scholar] [CrossRef]
- Dunn, G.P.; Bruce, A.T.; Sheehan, K.C.; Shankaran, V.; Uppaluri, R.; Bui, J.D.; Diamond, M.S.; Koebel, C.M.; Arthur, C.; White, J.; et al. A critical function for type I interferons in cancer immunoediting. Nat. Immunol. 2005, 6, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.S.; Kinder, M.; Matsushita, H.; Mashayekhi, M.; Dunn, G.P.; Archambault, J.M.; Lee, H.; Arthur, C.D.; White, J.; Kalinke, U.; et al. Type I interferon is selectively required by dendritic cells for immune rejection of tumors. J. Exp. Med. 2011, 208, 1989–2003. [Google Scholar] [CrossRef] [PubMed]
- Tschurtschenthaler, M.; Wang, J.; Fricke, C.; Fritz, T.M.; Niederreiter, L.; Adolph, T.E.; Sarcevic, E.; Künzel, S.; Offner, F.A.; Kalinke, U.; et al. Type I interferon signalling in the intestinal epithelium affects Paneth cells, microbial ecology and epithelial regeneration. Gut 2014, 63, 1921. [Google Scholar] [CrossRef] [PubMed]
- Katlinski, K.V.; Gui, J.; Katlinskaya, Y.V.; Ortiz, A.; Chakraborty, R.; Bhattacharya, S.; Carbone, C.J.; Beiting, D.P.; Girondo, M.A.; Peck, A.R.; et al. Inactivation of Interferon Receptor Promotes the Establishment of Immune Privileged Tumor Microenvironment. Cancer Cell 2017, 31, 194–207. [Google Scholar] [CrossRef] [PubMed]
- Fuertes, M.B.; Kacha, A.K.; Kline, J.; Woo, S.-R.R.; Kranz, D.M.; Murphy, K.M.; Gajewski, T.F. Host type I IFN signals are required for antitumor CD8+ T cell responses through CD8{alpha}+ dendritic cells. J. Exp. Med. 2011, 208, 2005–2016. [Google Scholar] [CrossRef]
- Bidwell, B.N.; Slaney, C.Y.; Withana, N.P.; Forster, S.; Cao, Y.; Loi, S.; Andrews, D.; Mikeska, T.; Mangan, N.E.; Samarajiwa, S.A.; et al. Silencing of Irf7 pathways in breast cancer cells promotes bone metastasis through immune escape. Nat. Med. 2012, 18, 1224. [Google Scholar] [CrossRef] [PubMed]
- Rautela, J.; Baschuk, N.; Slaney, C.Y.; Jayatilleke, K.M.; Xiao, K.; Bidwell, B.N.; Lucas, E.C.; Hawkins, E.D.; Lock, P.; Wong, C.S.; et al. Loss of Host Type-I IFN Signaling Accelerates Metastasis and Impairs NK-cell Antitumor Function in Multiple Models of Breast Cancer. Cancer Immunol. Res. 2015, 3, 1207–1217. [Google Scholar] [CrossRef]
- Lorenzi, S.; Mattei, F.; Sistigu, A.; Bracci, L.; Spadaro, F.; Sanchez, M.; Spada, M.; Belardelli, F.; Gabriele, L.; Schiavoni, G. Type I IFNs Control Antigen Retention and Survival of CD8α+ Dendritic Cells after Uptake of Tumor Apoptotic Cells Leading to Cross-Priming. J. Immunol. 2011, 186, 5142–5150. [Google Scholar] [CrossRef]
- Schiavoni, G.; Mattei, F.; Gabriele, L. Type I Interferons as Stimulators of DC-Mediated Cross-Priming: Impact on Anti-Tumor Response. Front. Immunol. 2013, 4, 483. [Google Scholar] [CrossRef]
- Mattei, F.; Schiavoni, G.; Belardelli, F.; Tough, D.F. IL-15 Is Expressed by Dendritic Cells in Response to Type I IFN, Double-Stranded RNA, or Lipopolysaccharide and Promotes Dendritic Cell Activation. J. Immunol. 2001, 167, 1179–1187. [Google Scholar] [CrossRef]
- Fuertes, M.B.; Woo, S.-R.; Burnett, B.; Fu, Y.-X.; Gajewski, T.F. Type I interferon response and innate immune sensing of cancer. Trends Immunol. 2013, 34, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Huntington, N.D. The unconventional expression of IL-15 and its role in NK cell homeostasis. Immunol. Cell Biol. 2014, 92, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Pace, L.; Vitale, S.; Dettori, B.; Palombi, C.; Sorsa, V.; Belardelli, F.; Proietti, E.; Doria, G. APC Activation by IFN-α Decreases Regulatory T Cell and Enhances Th Cell Functions. J. Immunol. 2010, 184, 5969–5979. [Google Scholar] [CrossRef] [PubMed]
- Sisirak, V.; Faget, J.; Gobert, M.; Goutagny, N.; Vey, N.; Treilleux, I.; Renaudineau, S.; Poyet, G.; Labidi-Galy, S.I.; Goddard-Leon, S.; et al. Impaired IFN-α production by plasmacytoid dendritic cells favors regulatory T-cell expansion that may contribute to breast cancer progression. Cancer Res. 2012, 72, 5188–5197. [Google Scholar] [CrossRef] [PubMed]
- Bacher, N.; Raker, V.; Hofmann, C.; Graulich, E.; Schwenk, M.; Baumgrass, R.; Bopp, T.; Zechner, U.; Merten, L.; Becker, C.; et al. Interferon-α Suppresses cAMP to Disarm Human Regulatory T Cells. Cancer Res. 2013, 73, 5647–5656. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hashimoto, H.; Ueda, R.; Narumi, K.; Heike, Y.; Yoshida, T.; Aoki, K. Type I IFN gene delivery suppresses regulatory T cells within tumors. Cancer Gene. Ther. 2014, 21, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Du, W.; Yan, F.; Wang, Y.; Li, H.; Cao, S.; Yu, W.; Shen, C.; Liu, J.; Ren, X. Myeloid-Derived Suppressor Cells Suppress Antitumor Immune Responses through IDO Expression and Correlate with Lymph Node Metastasis in Patients with Breast Cancer. J. Immunol. 2013, 190, 3783–3797. [Google Scholar] [CrossRef] [PubMed]
- Zoglmeier, C.; Bauer, H.; Nörenberg, D.; Wedekind, G.; Bittner, P.; Sandholzer, N.; Rapp, M.; Anz, D.; Endres, S.; Bourquin, C. CpG Blocks Immunosuppression by Myeloid-Derived Suppressor Cells in Tumor-Bearing Mice. Clin. Cancer Res. 2011, 17, 1765–1775. [Google Scholar] [CrossRef] [PubMed]
- Jongbloed, S.L.; Kassianos, A.J.; Nald, K.J.; Clark, G.J.; Ju, X.; Angel, C.E.; Chen, C.-J.J.; Dunbar, P.; Wadley, R.B.; Jeet, V.; et al. Human CD141+ (BDCA-3) + dendritic cells (DCs) represent a unique myeloid DC subset that cross-presents necrotic cell antigens. J. Exp. Med. 2010, 207, 1247–1260. [Google Scholar] [CrossRef]
- Poulin, L.; Salio, M.; Griessinger, E.; Anjos-Afonso, F.; Craciun, L.; Chen, J.-L.; Keller, A.M.; Joffre, O.; Zelenay, S.; Nye, E.; et al. Characterization of human DNGR-1+ BDCA3+ leukocytes as putative equivalents of mouse CD8α+ dendritic cells. J. Exp. Med. 2010, 207, 1261–1271. [Google Scholar] [CrossRef]
- Haniffa, M.; Shin, A.; Bigley, V.; McGovern, N.; Teo, P.; See, P.; Wasan, P.S.; Wang, X.-N.N.; Malinarich, F.; Malleret, B.; et al. Human tissues contain CD141hi cross-presenting dendritic cells with functional homology to mouse CD103+ nonlymphoid dendritic cells. Immunity 2012, 37, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Balan, S.; Ollion, V.; Colletti, N.; Chelbi, R.; Montanana-Sanchis, F.; Liu, H.; Vu Manh, T.-P.P.; Sanchez, C.; Savoret, J.; Perrot, I.; et al. Human XCR1+ dendritic cells derived in vitro from CD34+ progenitors closely resemble blood dendritic cells, including their adjuvant responsiveness, contrary to monocyte-derived dendritic cells. J. Immunol. 2014, 193, 1622–1635. [Google Scholar] [CrossRef] [PubMed]
- Lauterbach, H.; Bathke, B.; Gilles, S.; Traidl-Hoffmann, C.; Luber, C.A.; Fejer, G.; Freudenberg, M.A.; Vey, G.; Vremec, D.; Kallies, A.; et al. Mouse CD8α+ DCs and human BDCA3+ DCs are major producers of IFN-λ in response to poly IC. J. Exp. Med. 2010, 207, 2703–2717. [Google Scholar] [CrossRef] [PubMed]
- Yoshio, S.; Kanto, T.; Kuroda, S.; Matsubara, T.; Higashitani, K.; Kakita, N.; Ishida, H.; Hiramatsu, N.; Nagano, H.; Sugiyama, M.; et al. Human blood dendritic cell antigen 3 (BDCA3) + dendritic cells are a potent producer of interferon-λ in response to hepatitis C virus. Hepatology 2013, 57, 1705–1715. [Google Scholar] [CrossRef] [PubMed]
- Robbins, S.H.; Walzer, T.; Dembélé, D.; Thibault, C.; Defays, A.; Bessou, G.; Xu, H.; Vivier, E.; Sellars, M.; Pierre, P.; et al. Novel insights into the relationships between dendritic cell subsets in human and mouse revealed by genome-wide expression profiling. Genome Boil. 2008, 9, R17. [Google Scholar] [CrossRef] [PubMed]
- Galibert, L.; Diemer, G.S.; Liu, Z.; Johnson, R.S.; Smith, J.L.; Walzer, T.; Comeau, M.R.; Rauch, C.T.; Wolfson, M.F.; Sorensen, R.A.; et al. Nectin-like Protein 2 Defines a Subset of T-cell Zone Dendritic Cells and Is a Ligand for Class-I-restricted T-cell-associated Molecule. J. Biol. Chem. 2005, 280, 21955–21964. [Google Scholar] [CrossRef] [PubMed]
- Arase, N.; Takeuchi, A.; Unno, M.; Hirano, S.; Yokosuka, T.; Arase, H.; Saito, T. Heterotypic interaction of CRTAM with Necl2 induces cell adhesion on activated NK cells and CD8+ T cells. Int. Immunol. 2005, 17, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- Boles, K.S.; Barchet, W.; Diacovo, T.; Cella, M.; Colonna, M. The tumor suppressor TSLC1/NECL-2 triggers NK-cell and CD8+ T-cell responses through the cell-surface receptor CRTAM. Blood 2005, 106, 779–786. [Google Scholar] [CrossRef]
- Kennedy, J.; Vicari, A.P.; Saylor, V.; Zurawski, S.M.; Copeland, N.G.; Gilbert, D.J.; Jenkins, N.A.; Zlotnik, A. A molecular analysis of NKT cells: Identification of a class-I restricted T cell-associated molecule (CRTAM). J. Leukoc. Biol. 2000, 67, 725–734. [Google Scholar] [CrossRef]
- Bachem, A.; Güttler, S.; Hartung, E.; Ebstein, F.; Schaefer, M.; Tannert, A.; Salama, A.; Movassaghi, K.; Opitz, C.; Mages, H.W.; et al. Superior antigen cross-presentation and XCR1 expression define human CD11c+CD141+ cells as homologues of mouse CD8+ dendritic cells. J. Exp. Med. 2010, 207, 1273–1281. [Google Scholar] [CrossRef]
- Dorner, B.G.; Dorner, M.B.; Zhou, X.; Opitz, C.; Mora, A.; Güttler, S.; Hutloff, A.; Mages, H.W.; Ranke, K.; Schaefer, M.; et al. Selective expression of the chemokine receptor XCR1 on cross-presenting dendritic cells determines cooperation with CD8+ T cells. Immunity 2009, 31, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, C.; Miyamoto, R.; Hoshino, K.; Fukuda, Y.; Sasaki, I.; Saito, M.; Ishiguchi, H.; Yano, T.; Sugiyama, T.; Hemmi, H.; et al. Conservation of a chemokine system, XCR1 and its ligand, XCL1, between human and mice. Biochem. Biophys. Res. Commun. 2010, 397, 756–761. [Google Scholar] [CrossRef] [PubMed]
- Den Haan, J.; Lehar, S.; Bevan, M. CD8(+) but not CD8(-) dendritic cells cross-prime cytotoxic T cells in vivo. J. Exp. Med. 2000, 192, 1685–1696. [Google Scholar] [CrossRef] [PubMed]
- Dudziak, D.; Kamphorst, A.O.; Heidkamp, G.F.; Buchholz, V.R.; Trumpfheller, C.; Yamazaki, S.; Cheong, C.; Liu, K.; Lee, H.-W.W.; Park, C.G.; et al. Differential antigen processing by dendritic cell subsets in vivo. Science (N. Y.) 2007, 315, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Pooley, J.; Heath, W.; Shortman, K. Cutting edge: Intravenous soluble antigen is presented to CD4 T cells by CD8- dendritic cells, but cross-presented to CD8 T cells by CD8+ dendritic cells. J. Immunol. 2001, 166, 5327–5330. [Google Scholar] [CrossRef]
- Hildner, K.; Edelson, B.T.; Purtha, W.E.; Diamond, M.; Matsushita, H.; Kohyama, M.; Calderon, B.; Schraml, B.U.; Unanue, E.R.; Diamond, M.S.; et al. Batf3 Deficiency Reveals a Critical Role for CD8α+ Dendritic Cells in Cytotoxic T Cell Immunity. Science 2008, 322, 1097–1100. [Google Scholar] [CrossRef]
- Allan, R.S.; Smith, C.M.; Belz, G.T.; van Lint, A.L.; Wakim, L.M.; Heath, W.R.; Carbone, F.R. Epidermal Viral Immunity Induced by CD8α+ Dendritic Cells but Not by Langerhans Cells. Science 2003, 301, 1925–1928. [Google Scholar] [CrossRef]
- Belz, G.T.; Shortman, K.; Bevan, M.J.; Heath, W.R. CD8α+ Dendritic Cells Selectively Present MHC Class I-Restricted Noncytolytic Viral and Intracellular Bacterial Antigens in vivo. J. Immunol. 2005, 175, 196–200. [Google Scholar] [CrossRef]
- Belz, G.T.; Ith, C.; Eichner, D.; Shortman, K.; Karupiah, G.; Carbone, F.R.; Heath, W.R. Cutting edge: Conventional CD8 alpha+ dendritic cells are generally involved in priming CTL immunity to viruses. J. Immunol. 2004, 172, 1996–2000. [Google Scholar] [CrossRef]
- Schulz, O.; Diebold, S.S.; Chen, M.; Näslund, T.I.; Nolte, M.A.; Alexopoulou, L.; Azuma, Y.-T.; Flavell, R.A.; Liljeström, P.; e Sousa, C. Toll-like receptor 3 promotes cross-priming to virus-infected cells. Nature 2005, 433, 887. [Google Scholar] [CrossRef]
- Crozat, K.; Guiton, R.; Guilliams, M.; Henri, S.; Baranek, T.; Schwartz-Cornil, I.; Malissen, B.; Dalod, M. Comparative genomics as a tool to reveal functional equivalences between human and mouse dendritic cell subsets. Immunol. Rev. 2010, 234, 177–198. [Google Scholar] [CrossRef] [PubMed]
- Segura, E.; Durand, M.; Amigorena, S. Similar antigen cross-presentation capacity and phagocytic functions in all freshly isolated human lymphoid organ–resident dendritic cells. J. Exp. Med. 2013, 210, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, S.; Zelenay, S.; Sancho, D.; Hanč, P.; Kjær, S.; Feest, C.; Fletcher, G.; Durkin, C.; Postigo, A.; Skehel, M.; et al. F-actin is an evolutionarily conserved damage-associated molecular pattern recognized by DNGR-1, a receptor for dead cells. Immunity 2012, 36, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Sancho, D.; Joffre, O.P.; Keller, A.M.; Rogers, N.C.; Martínez, D.; Hernanz-Falcón, P.; Rosewell, I.; e Sousa, C. Identification of a dendritic cell receptor that couples sensing of necrosis to immunity. Nature 2009, 458, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-G.; Czabotar, P.E.; Policheni, A.N.; Caminschi, I.; San Wan, S.; Kitsoulis, S.; Tullett, K.M.; Robin, A.Y.; Brammananth, R.; van Delft, M.F.; et al. The Dendritic Cell Receptor Clec9A Binds Damaged Cells via Exposed Actin Filaments. Immunity 2012, 36, 646–657. [Google Scholar] [CrossRef] [PubMed]
- Deauvieau, F.; Ollion, V.; Doffin, A.-C.C.; Achard, C.; Fonteneau, J.-F.F.; Verronese, E.; Durand, I.; Ghittoni, R.; Marvel, J.; Dezutter-Dambuyant, C.; et al. Human natural killer cells promote cross-presentation of tumor cell-derived antigens by dendritic cells. Int. J. Cancer 2015, 136, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Broz, M.L.; Binnewies, M.; Boldajipour, B.; Nelson, A.E.; Pollack, J.L.; Erle, D.J.; Barczak, A.; Rosenblum, M.D.; Daud, A.; Barber, D.L.; et al. Dissecting the Tumor Myeloid Compartment Reveals Rare Activating Antigen-Presenting Cells Critical for T Cell Immunity. Cancer Cell 2014, 26, 638–652. [Google Scholar] [CrossRef] [PubMed]
- Roberts, E.W.; Broz, M.L.; Binnewies, M.; Headley, M.B.; Nelson, A.E.; Wolf, D.M.; Kaisho, T.; Bogunovic, D.; Bhardwaj, N.; Krummel, M.F. Critical Role for CD103+/CD141+ Dendritic Cells Bearing CCR7 for Tumor Antigen Trafficking and Priming of T Cell Immunity in Melanoma. Cancer Cell 2016, 30, 324–336. [Google Scholar] [CrossRef]
- Headley, M.B.; Bins, A.; Nip, A.; Roberts, E.W.; Looney, M.R.; Gerard, A.; Krummel, M.F. Visualization of immediate immune responses to pioneer metastatic cells in the lung. Nature 2016, 531, 513. [Google Scholar] [CrossRef]
- Laoui, D.; Keirsse, J.; Morias, Y.; Overmeire, E.; Geeraerts, X.; Elkrim, Y.; Kiss, M.; Bolli, E.; Lahmar, Q.; Sichien, D.; et al. The tumour microenvironment harbours ontogenically distinct dendritic cell populations with opposing effects on tumour immunity. Nat. Commun. 2016, 7, 13720. [Google Scholar] [CrossRef]
- Salmon, H.; Idoyaga, J.; Rahman, A.; Leboeuf, M.; Remark, R.; Jordan, S.; Casanova-Acebes, M.; Khudoynazarova, M.; Agudo, J.; Tung, N.; et al. Expansion and Activation of CD103(+) Dendritic Cell Progenitors at the Tumor Site Enhances Tumor Responses to Therapeutic PD-L1 and BRAF Inhibition. Immunity 2016, 44, 924–938. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Paulete, A.R.; Cueto, F.J.; Martínez-López, M.; Labiano, S.; Morales-Kastresana, A.; Rodríguez-Ruiz, M.E.; Jure-Kunkel, M.; Azpilikueta, A.; Aznar, M.; Quetglas, J.I.; et al. Cancer Immunotherapy with Immunomodulatory Anti-CD137 and Anti-PD-1 Monoclonal Antibodies Requires BATF3-Dependent Dendritic Cells. Cancer Dis. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Spranger, S.; Bao, R.; Gajewski, T.F. Melanoma-intrinsic β-catenin signalling prevents anti-tumour immunity. Nature 2015, 523, 231–235. [Google Scholar] [CrossRef]
- Spranger, S.; Dai, D.; Horton, B.; Gajewski, T.F. Tumor-Residing Batf3 Dendritic Cells Are Required for Effector T Cell Trafficking and Adoptive T Cell Therapy. Cancer Cell 2017, 31, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Hémont, C.; Neel, A.; Heslan, M.; Braudeau, C.; Josien, R. Human blood mDC subsets exhibit distinct TLR repertoire and responsiveness. J. Leukoc. Biol. 2013, 93, 599–609. [Google Scholar] [CrossRef]
- Jin, J.-O.; Zhang, W.; Du, J.; Yu, Q. BDCA1-Positive Dendritic Cells (DCs) Represent a Unique Human Myeloid DC Subset That Induces Innate and Adaptive Immune Responses to Staphylococcus aureus Infection. Infect. Immun. 2014, 82, 4466–4476. [Google Scholar] [CrossRef] [PubMed]
- Nizzoli, G.; Krietsch, J.; Weick, A.; Steinfelder, S.; Facciotti, F.; Gruarin, P.; Bianco, A.; Steckel, B.; Moro, M.; Crosti, M.; et al. Human CD1c+ dendritic cells secrete high levels of IL-12 and potently prime cytotoxic T-cell responses. Blood 2013, 122, 932–942. [Google Scholar] [CrossRef]
- Sittig, S.P.; Bakdash, G.; Weiden, J.; Sköld, A.E.; Tel, J.; Figdor, C.G.; de Vries, J.I.; Schreibelt, G. A Comparative Study of the T Cell Stimulatory and Polarizing Capacity of Human Primary Blood Dendritic Cell Subsets. Mediat. Inflamm. 2016, 2016, 3605643. [Google Scholar] [CrossRef]
- Dillon, S.M.; Rogers, L.M.; Howe, R.; Hostetler, L.A.; Buhrman, J.; McCarter, M.D.; Wilson, C.C. Human Intestinal Lamina Propria CD1c+ Dendritic Cells Display an Activated Phenotype at Steady State and Produce IL-23 in Response to TLR7/8 Stimulation. J. Immunol. 2010, 184, 6612–6621. [Google Scholar] [CrossRef]
- Kassianos, A.J.; Hardy, M.Y.; Ju, X.; Vijayan, D.; Ding, Y.; Vulink, A.J.; Nald, K.J.; Jongbloed, S.L.; Wadley, R.B.; Wells, C.; et al. Human CD1c (BDCA-1) + myeloid dendritic cells secrete IL-10 and display an immuno-regulatory phenotype and function in response to Escherichia coli. Eur. J. Immunol. 2012, 42, 1512–1522. [Google Scholar] [CrossRef]
- Blasio, S.; Wortel, I.M.; van Bladel, D.A.; de Vries, L.E.; Boer, T.; Worah, K.; de Haas, N.; Buschow, S.I.; de Vries, J.I.; Figdor, C.G.; et al. Human CD1c (+) DCs are critical cellular mediators of immune responses induced by immunogenic cell death. Oncoimmunology 2016, 5, e1192739. [Google Scholar] [CrossRef] [PubMed]
- Cohn, L.; Chatterjee, B.; Esselborn, F.; Smed-Sörensen, A.; Nakamura, N.; Chalouni, C.; Lee, B.-C.; Vandlen, R.; Keler, T.; Lauer, P.; et al. Antigen delivery to early endosomes eliminates the superiority of human blood BDCA3+ dendritic cells at cross presentation. J. Exp. Med. 2013, 210, 1049–1063. [Google Scholar] [CrossRef] [PubMed]
- Mittag, D.; Proietto, A.I.; Loudovaris, T.; Mannering, S.I.; Vremec, D.; Shortman, K.; Wu, L.; Harrison, L.C. Human dendritic cell subsets from spleen and blood are similar in phenotype and function but modified by donor health status. J. Immunol. 2011, 186, 6207–6217. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Nish, S.A.; Jiang, R.; Hou, L.; Licona-Limón, P.; Weinstein, J.S.; Zhao, H.; Medzhitov, R. Control of T Helper 2 Responses by Transcription Factor IRF4-Dependent Dendritic Cells. Immunity 2013, 39, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswamy, J.; Gowthaman, U.; Zhang, B.; Mattsson, J.; Szeponik, L.; Liu, D.; Wu, R.; White, T.; Calabro, S.; Xu, L.; et al. Migratory CD11b+ conventional dendritic cells induce T follicular helper cell–dependent antibody responses. Sci. Immunol. 2017, 2, eaam9169. [Google Scholar] [CrossRef]
- Binnewies, M.; Mujal, A.M.; Pollack, J.L.; Combes, A.J.; Hardison, E.A.; Barry, K.C.; Tsui, J.; Ruhland, M.K.; Kersten, K.; Abushawish, M.A.; et al. Unleashing Type-2 Dendritic Cells to Drive Protective Antitumor CD4+ T Cell Immunity. Cell 2019, 177, 556–571. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Honma, K.; Matsuyama, T.; Suzuki, K.; Toriyama, K.; Akitoyo, I.; Yamamoto, K.; Suematsu, T.; Nakamura, M.; Yui, K.; et al. Critical roles of interferon regulatory factor 4 in CD11bhighCD8α– dendritic cell development. Proc. Natl. Acad. Sci. USA 2004, 101, 8981–8986. [Google Scholar] [CrossRef]
- Tamura, T.; Tailor, P.; Yamaoka, K.; Kong, H.; Tsujimura, H.; O’Shea, J.J.; Singh, H.; Ozato, K. IFN Regulatory Factor-4 and -8 Govern Dendritic Cell Subset Development and Their Functional Diversity. J. Immunol. 2005, 174, 2573–2581. [Google Scholar] [CrossRef]
- Bajaña, S.; Roach, K.; Turner, S.; Paul, J.; Kovats, S. IRF4 Promotes Cutaneous Dendritic Cell Migration to Lymph Nodes during Homeostasis and Inflammation. J. Immunol. 2012, 189, 3368–3377. [Google Scholar] [CrossRef]
- Valladeau, J.; Ravel, O.; Dezutter-Dambuyant, C.; Moore, K.; Kleijmeer, M.; Liu, Y.; Duvert-Frances, V.; Vincent, C.; Schmitt, D.; Davoust, J.; et al. Langerin, a Novel C-Type Lectin Specific to Langerhans Cells, is an Endocytic Receptor that Induces the Formation of Birbeck Granules. Immunity 2000, 12, 71–81. [Google Scholar] [CrossRef]
- Ginhoux, F.; Tacke, F.; Angeli, V.; Bogunovic, M.; Loubeau, M.; Dai, X.-M.; Stanley, R.E.; Randolph, G.J.; Merad, M. Langerhans cells arise from monocytes in vivo. Nat. Immunol. 2006, 7, ni1307. [Google Scholar] [CrossRef] [PubMed]
- Greter, M.; Lelios, I.; Pelczar, P.; Hoeffel, G.; Price, J.; Leboeuf, M.; Kündig, T.M.; Frei, K.; Ginhoux, F.; Merad, M.; et al. Stroma-Derived Interleukin-34 Controls the Development and Maintenance of Langerhans Cells and the Maintenance of Microglia. Immunity 2012, 37, 1050–1060. [Google Scholar] [CrossRef] [PubMed]
- Klechevsky, E.; Morita, R.; Liu, M.; Cao, Y.; Coquery, S.; Thompson-Snipes, L.; Briere, F.; Chaussabel, D.; Zurawski, G.; Palucka, K.A.; et al. Functional Specializations of Human Epidermal Langerhans Cells and CD14+ Dermal Dendritic Cells. Immunity 2008, 29, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Artyomov, M.N.; Munk, A.; Gorvel, L.; Korenfeld, D.; Cella, M.; Tung, T.; Klechevsky, E. Modular expression analysis reveals functional conservation between human Langerhans cells and mouse cross-priming dendritic cells. J. Exp. Med. 2015, 212, 743–757. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Thompson-Snipes, L.; Zurawski, S.; Blanck, J.-P.P.; Cao, Y.; Clayton, S.; Gorvel, J.-P.P.; Zurawski, G.; Klechevsky, E. The differential production of cytokines by human Langerhans cells and dermal CD14(+) DCs controls CTL priming. Blood 2012, 119, 5742–5749. [Google Scholar] [CrossRef] [PubMed]
- Romano, E.; Cotari, J.W.; da Silva, R.; Betts, B.C.; Chung, D.J.; Avogadri, F.; Fink, M.J.; Angelo, E.T.; Mehrara, B.; Heller, G.; et al. Human Langerhans cells use an IL-15R-α/IL-15/pSTAT5-dependent mechanism to break T-cell tolerance against the self-differentiation tumor antigen WT1. Blood 2012, 119, 5182–5190. [Google Scholar] [CrossRef] [PubMed]
- León, B.; López-Bravo, M.; Ardavín, C. Monocyte-Derived Dendritic Cells Formed at the Infection Site Control the Induction of Protective T Helper 1 Responses against Leishmania. Immunity 2007, 26, 519–531. [Google Scholar] [CrossRef]
- León, B.; López-Bravo, M.; Ardavín, C. Monocyte-derived dendritic cells. Semin. Immunol. 2005, 17, 313–318. [Google Scholar] [CrossRef]
- Sallusto, F.; Lanzavecchia, A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J. Exp. Med. 1994, 179, 1109–1118. [Google Scholar] [CrossRef]
- Wollenberg, A.; Oppel, T.; Schottdorf, E.-M.; Günther, S.; Moderer, M.; Mommaas, M. Expression and Function of the Mannose Receptor CD206 on Epidermal Dendritic Cells in Inflammatory Skin Diseases. J. Investig. Dermatol. 2002, 118, 327–334. [Google Scholar] [CrossRef]
- Wollenberg, A.; Kraft, S.; Hanau, D.; Bieber, T. Immunomorphological and Ultrastructural Characterization of Langerhans Cells and a Novel, Inflammatory Dendritic Epidermal Cell (IDEC) Population in Lesional Skin of Atopic Eczema. J. Investig. Dermatol. 1996, 106, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Qian, B.-Z.; Li, J.; Zhang, H.; Kitamura, T.; Zhang, J.; Campion, L.R.; Kaiser, E.A.; Snyder, L.A.; Pollard, J.W. CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature 2011, 475, 222. [Google Scholar] [CrossRef] [PubMed]
- Shand, F.H.; Ueha, S.; Otsuji, M.; Koid, S.; Shichino, S.; Tsukui, T.; Kosugi-Kanaya, M.; Abe, J.; Tomura, M.; Ziogas, J.; et al. Tracking of intertissue migration reveals the origins of tumor-infiltrating monocytes. Proc. Natl. Acad. Sci. USA 2014, 111, 7771–7776. [Google Scholar] [CrossRef] [PubMed]
- Bakdash, G.; Buschow, S.I.; Gorris, M.; Halilovic, A.; Hato, S.V.; Sköld, A.E.; Schreibelt, G.; Sittig, S.P.; Torensma, R.; Boer, T.; et al. Expansion of a BDCA1+CD14+ Myeloid Cell Population in Melanoma Patients May Attenuate the Efficacy of Dendritic Cell Vaccines. Cancer Res. 2016, 76, 4332–4346. [Google Scholar] [CrossRef] [PubMed]
- Nagorsen, D.; Voigt, S.; Berg, E.; Stein, H.; Thiel, E.; Loddenkemper, C. Tumor-infiltrating macrophages and dendritic cells in human colorectal cancer: Relation to local regulatory T cells, systemic T-cell response against tumor-associated antigens and survival. J. Transl. Med. 2007, 5, 62. [Google Scholar] [CrossRef] [PubMed]
- Bell, D.; Chomarat, P.; Broyles, D.; Netto, G.; Harb, G.; Lebecque, S.; Valladeau, J.; Davoust, J.; Palucka, K.; Banchereau, J. In breast carcinoma tissue, immature dendritic cells reside within the tumor, whereas mature dendritic cells are located in peritumoral areas. J. Exp. Med. 1999, 190, 1417–1426. [Google Scholar] [CrossRef] [PubMed]
- Hillenbrand, E.; Neville, A.; Coventry, B. Immunohistochemical localization of CD1a-positive putative dendritic cells in human breast tumours. Br. J. Cancer 1999, 79, 6690150. [Google Scholar] [CrossRef]
- Tsuge, T.; Yamakawa, M.; Tsukamoto, M. Infiltrating dendritic/langerhans cells in primary breast cancer. Breast Cancer Res. Treat. 2000, 59, 141–152. [Google Scholar] [CrossRef]
- Coventry, B.; Lee, P.-L.; Gibbs, D.; Hart, D. Dendritic cell density and activation status in human breast cancer—CD1a, CMRF-44, CMRF-56 and CD-83 expression. Br. J. Cancer 2002, 86, 546. [Google Scholar] [CrossRef]
- Iwamoto, M.; Shinohara, H.; Miyamoto, A.; Okuzawa, M.; Mabuchi, H.; Nohara, T.; Gon, G.; Toyoda, M.; Tanigawa, N. Prognostic value of tumor-infiltrating dendritic cells expressing CD83 in human breast carcinomas. Int. J. Cancer 2003, 104, 92–97. [Google Scholar] [CrossRef]
- Coventry, B.; Morton, J. CD1a-positive infiltrating-dendritic cell density and 5-Year survival from human breast cancer. Br. J. Cancer 2003, 89, 6601114. [Google Scholar] [CrossRef] [PubMed]
- Treilleux, I.; Blay, J.-Y.Y.; Bendriss-Vermare, N.; Ray-Coquard, I.; Bachelot, T.; Guastalla, J.-P.P.; Bremond, A.; Goddard, S.; Pin, J.-J.J.; Barthelemy-Dubois, C.; et al. Dendritic Cell Infiltration and Prognosis of Early Stage Breast Cancer. Clin. Cancer Res. An Off. J. Am. Assoc. Cancer Res. 2004, 10, 7466–7474. [Google Scholar] [CrossRef] [PubMed]
- Martinet, L.; Filleron, T.; Guellec, S.; Rochaix, P.; Garrido, I.; Girard, J.-P. High Endothelial Venule Blood Vessels for Tumor-Infiltrating Lymphocytes are Associated wth Lymphotoxin Β–Producing Dendritic Cells in Human Breast Cancer. J. Immunol. 2013, 191, 2001–2008. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Masuda, A.; Nagata, H.; Kameoka, S.; Kikawada, Y.; Yamakawa, M.; Kasajima, T. Mature Dendritic Cells Make Clusters with T Cells in the Invasive Margin of Colorectal Carcinoma. J. Pathol. 2002, 196, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Sandel, M.H.; Dadabayev, A.R.; Menon, A.G.; Morreau, H.; Melief, C.; Offringa, R.; van der Burg, S.H.; Rhijn, C.M.; Ensink, G.N.; Tollenaar, R.; et al. Prognostic Value of Tumor-Infiltrating Dendritic Cells in Colorectal Cancer: Role of Maturation Status and Intratumoral Localization. Clin. Cancer Res. 2005, 11, 2576–2582. [Google Scholar] [CrossRef] [PubMed]
- Perrot, I.; Blanchard, D.; Freymond, N.; Isaac, S.; Guibert, B.; Pachéco, Y.; Lebecque, S. Dendritic Cells Infiltrating Human Non-Small Cell Lung Cancer are Blocked at Immature Stage. J. Immunol. 2007, 178, 2763–2769. [Google Scholar] [CrossRef]
- Dieu-Nosjean, M.-C.C.; Antoine, M.; Danel, C.; Heudes, D.; Wislez, M.; Poulot, V.; Rabbe, N.; Laurans, L.; Tartour, E.; de Chaisemartin, L.; et al. Long-Term Survival for Patients with Non-Small-Cell Lung Cancer with Intratumoral Lymphoid Structures. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2008, 26, 4410–4417. [Google Scholar] [CrossRef]
- Vermi, W.; Bonecchi, R.; Facchetti, F.; Bianchi, D.; Sozzani, S.; Festa, S.; Berenzi, A.; Cella, M.; Colonna, M. Recruitment of Immature Plasmacytoid Dendritic Cells (Plasmacytoid Monocytes) and Myeloid Dendritic Cells in Primary Cutaneous Melanomas. J. Pathol. 2003, 200, 255–268. [Google Scholar] [CrossRef]
- Movassagh, M.; Spatz, A.; Davoust, J.; Lebecque, S.; Romero, P.; Pittet, M.; Rimoldi, D.; Liénard, D.; Gugerli, O.; Ferradini, L.; et al. Selective Accumulation of Mature DC-Lamp + Dendritic Cells in Tumor Sites is Associated with Efficient T-Cell-Mediated Antitumor Response and Control of Metastatic Dissemination in Melanoma. Cancer Res. 2004, 64, 2192–2198. [Google Scholar] [CrossRef]
- Ladányi, A.; Kiss, J.; Somlai, B.; Gilde, K.; Fejos, Z.; Mohos, A.; Gaudi, I.; Tímár, J. Density of DC-LAMP (+) Mature Dendritic Cells in Combination with Activated T Lymphocytes Infiltrating Primary Cutaneous Melanoma is a Strong Independent Prognostic Factor. Cancer Immunol. Immunother. CII 2007, 56, 1459–1469. [Google Scholar] [CrossRef]
- Labidi-Galy, S.; Treilleux, I.; Goddard-Leon, S.; Combes, J.-D.; Blay, J.-Y.; Ray-Coquard, I.; Caux, C.; Bendriss-Vermare, N. Plasmacytoid dendritic cells infiltrating ovarian cancer are associated with poor prognosis. Oncoimmunology 2012, 1, 380–382. [Google Scholar] [CrossRef] [PubMed]
- Thurnher, M.; Radmayr, C.; Ramoner, R.; Ebner, S.; Böck, G.; Klocker, H.; Romani, N.; Bartsch, G. Human Renal-Cell Carcinoma Tissue Contains Dendritic Cells. Int. J. Cancer 1996, 68, 1–7. [Google Scholar] [CrossRef]
- Troy, A.J.; Summers, K.L.; Davidson, P.J.T.; Atkinson, C.H.; Hart, D.N.J. Minimal Recruitment and Activation of Dendritic Cells within Renal Cell Carcinoma. J. Urol. 1999, 161, 1737–1738. [Google Scholar] [CrossRef]
- Schwaab, T.; Schned, A.R.; Heaney, J.A.; Cole, B.F.; Atzpodien, J.; Wittke, F.; Estoff, M.S. In vivo Description of Dendritic Cells in Human Renal Cell Carcinoma. J. Urol. 1999, 162, 567–573. [Google Scholar] [CrossRef]
- Feng, J.; Chen, Y.; Shi, B.; Yan, D.; Wagn, J. Expression and Significance of Tumor Infiltrating Dendritic Cells in Renal Cell Carcinoma. Chin. J. Cancer Res. 2005, 17, 127–131. [Google Scholar] [CrossRef]
- Kerrebijn, J.D.; Balm, A.J.; Knegt, P.P.; Meeuwis, C.A.; Drexhage, H.A. Macrophage and Dendritic Cell Infiltration in Head and Neck Squamous-Cell Carcinoma; an Immunohistochemical Study. Cancer Immunol. Immunother. 1994, 38, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Goldman, S.A.; Baker, E.; Weyant, R.J.; Clarke, M.R.; Myers, J.N.; Lotze, M.T. Peritumoral CD1a-positive Dendritic Cells are Associated with Improved Survival in Patients with Tongue Carcinoma. Arch. Otolaryngol. Head Neck Surg. 1998, 124, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, E.; Wollenberg, B.; Rothenfusser, S.; Wagner, M.; Wellisch, D.; Mack, B.; Giese, T.; Gires, O.; Endres, S.; Hartmann, G. Identification and Functional Analysis of Tumor-Infiltrating Plasmacytoid Dendritic Cells in Head and Neck Cancer. Cancer Res. 2003, 63, 6478–6487. [Google Scholar]
- Li, Y.; Li, Z.; Lin, H.; Chen, X.; Li, B. Primary Cutaneous Blastic Plasmacytoid Dendritic Cell Neoplasm without Extracutaneous Manifestation: Case Report and Review of the Literature. Pathol. Res. Pract. 2011, 207, 55–59. [Google Scholar] [CrossRef]
- Ayari, C.; LaRue, H.; Hovington, H.; Decobert, M.; Harel, F.; Bergeron, A.; Têtu, B.; Lacombe, L.; Fradet, Y. Bladder Tumor Infiltrating Mature Dendritic Cells and Macrophages as Predictors of Response to Bacillus Calmette-Guérin Immunotherapy. Eur. Urol. 2009, 55, 1386–1396. [Google Scholar] [CrossRef]
- Ishigami, S.; Ueno, S.; Matsumoto, M.; Okumura, H.; Arigami, T.; Uchikado, Y.; Setoyama, T.; Arima, H.; Sasaki, K.; Kitazono, M.; et al. Prognostic Value of CD208-positive Cell Infiltration in Gastric Cancer. Cancer Immunol. Immunother. 2009, 59, 389. [Google Scholar] [CrossRef] [PubMed]
- Labidi-Galy, S.I.; Sisirak, V.; Meeus, P.; Gobert, M.; Treilleux, I.; Bajard, A.; Combes, J.-D.D.; Faget, J.; Mithieux, F.; Cassignol, A.; et al. Quantitative and functional alterations of plasmacytoid dendritic cells contribute to immune tolerance in ovarian cancer. Cancer Res. 2011, 71, 5423–5434. [Google Scholar] [CrossRef] [PubMed]
- Michea, P.; Noël, F.; Zakine, E.; Czerwinska, U.; Sirven, P.; Abouzid, O.; Goudot, C.; Scholer-Dahirel, A.; Vincent-Salomon, A.; Reyal, F.; et al. Adjustment of dendritic cells to the breast-cancer microenvironment is subset specific. Nat. Immunol. 2018, 19, 885–897. [Google Scholar] [CrossRef]
- Segura, E.; Touzot, M.; Bohineust, A.; Cappuccio, A.; Chiocchia, G.; Hosmalin, A.; Dalod, M.; Soumelis, V.; Amigorena, S. Human Inflammatory Dendritic Cells Induce Th17 Cell Differentiation. Immunity 2013, 38, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Tabarkiewicz, J.; Rybojad, P.; Jablonka, A.; Rolinski, J. CD1c+ and CD303+ Dendritic Cells in Peripheral Blood, Lymph Nodes and Tumor Tissue of Patients with Non-Small Cell Lung Cancer. Oncol. Rep. 2008, 19, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Aspord, C.; Leccia, M.-T.; Charles, J.; Plumas, J. Plasmacytoid Dendritic Cells Support Melanoma Progression by Promoting Th2 and Regulatory Immunity through OX40L and ICOSL. Cancer Immunol. Res. 2013, 1, 402–415. [Google Scholar] [CrossRef]
- Zou, W.; Machelon, V.; Coulomb-L’Hermin, A.; Borvak, J.; Nome, F.; Isaeva, T.; Wei, S.; Krzysiek, R.; Durand-Gasselin, I.; Gordon, A.; et al. Stromal-Derived Factor-1 in Human Tumors Recruits and Alters the Function of Plasmacytoid Precursor Dendritic Cells. Nat. Med. 2001, 7, 1339. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Kryczek, I.; Zou, L.; Daniel, B.; Cheng, P.; Mottram, P.; Curiel, T.; Lange, A.; Zou, W. Plasmacytoid Dendritic Cells Induce CD8+ Regulatory T Cells in Human Ovarian Carcinoma. Cancer Res. 2005, 65, 5020–5026. [Google Scholar] [CrossRef] [PubMed]
- Alcántara-Hernández, M.; Leylek, R.; Wagar, L.E.; Engleman, E.G.; Keler, T.; Marinkovich, P.M.; Davis, M.M.; Nolan, G.P.; Idoyaga, J. High-Dimensional Phenotypic Mapping of Human Dendritic Cells Reveals Interindividual Variation and Tissue Specialization. Immunity 2017, 47, 1037–1050. [Google Scholar] [CrossRef]
- Vu Manh, T.-P.P.; Dalod, M. Characterization of Dendritic Cell Subsets Through Gene Expression Analysis. Methods Mol. Biol. (Clifton N.J.) 2016, 1423, 211–243. [Google Scholar]
- Miller, J.C.; Brown, B.D.; Shay, T.; Gautier, E.L.; Jojic, V.; Cohain, A.; Pandey, G.; Leboeuf, M.; Elpek, K.G.; Helft, J.; et al. Deciphering the transcriptional network of the dendritic cell lineage. Nat. Immunol. 2012, 13, 888–899. [Google Scholar] [CrossRef] [PubMed]
- Heidkamp, G.F.; Sander, J.; Lehmann, C.H.; Heger, L.; Eissing, N.; Baranska, A.; Lühr, J.J.; Hoffmann, A.; Reimer, K.C.; Lux, A.; et al. Human lymphoid organ dendritic cell identity is predominantly dictated by ontogeny, not tissue microenvironment. Sci. Immunol. 2016, 1, eaai7677. [Google Scholar] [CrossRef] [PubMed]
- Contreras, V.; Urien, C.; Guiton, R.; Alexandre, Y.; Vu Manh, T.-P.; Andrieu, T.; Crozat, K.; Jouneau, L.; Bertho, N.; Epardaud, M.; et al. Existence of CD8α-Like Dendritic Cells with a Conserved Functional Specialization and a Common Molecular Signature in Distant Mammalian Species. J. Immunol. 2010, 185, 3313–3325. [Google Scholar] [CrossRef] [PubMed]
- Marquet, F.; Vu Manh, T.-P.; Maisonnasse, P.; Elhmouzi-Younes, J.; Urien, C.; Bouguyon, E.; Jouneau, L.; Bourge, M.; Simon, G.; Ezquerra, A.; et al. Pig Skin Includes Dendritic Cell Subsets Transcriptomically Related to Human CD1a and CD14 Dendritic Cells Presenting Different Migrating Behaviors and T Cell Activation Capacities. J. Immunol. 2014, 193, 5883–5893. [Google Scholar] [CrossRef] [PubMed]
- Vu Manh, T.; Marty, H.; Sibille, P.; Vern, L.Y.; Kaspers, B.; Dalod, M.I.; Quéré, P. Existence of conventional dendritic cells in Gallus gallus revealed by comparative gene expression profiling. J. Immunol. 2014, 192, 4510–4517. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, C.-A.; Wang, L.-F.; Ginhoux, F. Aligning bona fide dendritic cell populations across species. Cell Immunol. 2014, 291, 3–10. [Google Scholar] [CrossRef]
- Crozat, K.; Guiton, R.; Contreras, V.; Feuillet, V.; Dutertre, C.-A.A.; Ventre, E.; Vu Manh, T.-P.P.; Baranek, T.; Storset, A.K.; Marvel, J.; et al. The XC Chemokine Receptor 1 is a Conserved Selective Marker of Mammalian Cells Homologous to Mouse CD8α+ Dendritic Cells. J. Exp. Med. 2010, 207, 1283–1292. [Google Scholar] [CrossRef]
- Chevrier, S.; Levine, J.; Zanotelli, V.; Silina, K.; Schulz, D.; Bacac, M.; Ries, C.; Ailles, L.; Jewett, M.; Moch, H.; et al. An Immune Atlas of Clear Cell Renal Cell Carcinoma. Cell 2017, 169, 736–749. [Google Scholar] [CrossRef]
- Lavin, Y.; Kobayashi, S.; Leader, A.; Amir, E.-A.D.; Elefant, N.; Bigenwald, C.; Remark, R.; Sweeney, R.; Becker, C.D.; Levine, J.H.; et al. Innate Immune Landscape in Early Lung Adenocarcinoma by Paired Single-Cell Analyses. Cell 2017, 169, 750–765. [Google Scholar] [CrossRef]
- Zilionis, R.; Engblom, C.; Pfirschke, C.; Savova, V.; Zemmour, D.; Saatcioglu, H.D.; Krishnan, I.; Maroni, G.; Meyerovitz, C.V.; Kerwin, C.M.; et al. Single-Cell Transcriptomics of Human and Mouse Lung Cancers Reveals Conserved Myeloid Populations across Individuals and Species. Immunity 2019, 50, 1317–1334. [Google Scholar] [CrossRef]
- Lee, J.; Breton, G.; Oliveira, T.; Zhou, Y.; Aljoufi, A.; Puhr, S.; Cameron, M.J.; Sékaly, R.-P.; Nussenzweig, M.C.; Liu, K. Restricted dendritic cell and monocyte progenitors in human cord blood and bone marrow. J. Exp. Med. 2015, 212, 385–399. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.X.; Terry, J.M.; Belgrader, P.; Ryvkin, P.; Bent, Z.W.; Wilson, R.; Ziraldo, S.B.; Wheeler, T.D.; Rmott, G.P.; Zhu, J.; et al. Massively parallel digital transcriptional profiling of single cells. Nat. Commun. 2017, 8, ncomms14049. [Google Scholar] [CrossRef] [PubMed]
- Barnes, T.A.; Amir, E. HYPE or HOPE: The Prognostic Value of Infiltrating Immune Cells in Cancer. Br. J. Cancer 2017, 117, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Gnjatic, S.; Bronte, V.; Brunet, L.; Butler, M.O.; Disis, M.L.; Galon, J.; Hakansson, L.G.; Hanks, B.A.; Karanikas, V.; Khleif, S.N.; et al. Identifying Baseline Immune-Related Biomarkers to Predict Clinical Outcome of Immunotherapy. J. Immunother. Cancer 2017, 5, 44. [Google Scholar] [CrossRef] [PubMed]
- Jardim, J.F.; Gondak, R.; Galvis, M.M.; Pinto, C.A.; Kowalski, L.P. A Decreased Peritumoral CD1a+ Cell Number Predicts a Worse Prognosis in Oral Squamous Cell Carcinoma. Histopathology 2018, 72, 905–913. [Google Scholar] [CrossRef]
- Kindt, N.; Descamps, G.; Seminerio, I.; Bellier, J.; Lechien, J.R.; Pottier, C.; Larsimont, D.; Journé, F.; Delvenne, P.; Saussez, S. Langerhans Cell Number is a Strong and Independent Prognostic Factor for Head and Neck Squamous Cell Carcinomas. Oral. Oncol. 2016, 62, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Al-Shibli, K.; Al-Saad, S.; Donnem, T.; Persson, M.; Bremnes, R.M.; Busund, L. The Prognostic Value of Intraepithelial and Stromal Innate Immune System Cells in Non-Small Cell Lung Carcinoma. Histopathology 2009, 55, 301–312. [Google Scholar] [CrossRef]
- Koirala, P.; Roth, M.E.; Gill, J.; Piperdi, S.; Chinai, J.M.; Geller, D.S.; Hoang, B.H.; Park, A.; Fremed, M.A.; Zang, X.; et al. Immune Infiltration and PD-L1 Expression in the Tumor Microenvironment are Prognostic In Osteosarcoma. Sci. Rep. UK 2016, 6, 30093. [Google Scholar] [CrossRef]
- Lundgren, S.; Karnevi, E.; Elebro, J.; Nodin, B.; Karlsson, M.C.; Eberhard, J.; Leandersson, K.; Jirström, K. The Clinical Importance of Tumour-Infiltrating Macrophages and Dendritic Cells in Periampullary Adenocarcinoma Differs by Morphological Subtype. J. Transl. Med. 2017, 15, 152. [Google Scholar] [CrossRef]
- Alifano, M.; Mansuet-Lupo, A.; Lococo, F.; Roche, N.; Bobbio, A.; Canny, E.; Schussler, O.; Dermine, H.; Régnard, J.-F.; Burroni, B.; et al. Systemic Inflammation, Nutritional Status and Tumor Immune Microenvironment Determine Outcome of Resected Non-Small Cell Lung Cancer. PLoS ONE 2014, 9, e106914. [Google Scholar] [CrossRef]
- Goc, J.; Germain, C.; Vo-Bourgais, T.; Lupo, A.; Klein, C.; Knockaert, S.; de Chaisemartin, L.; Ouakrim, H.; Becht, E.; Alifano, M.; et al. Dendritic Cells in Tumor-Associated Tertiary Lymphoid Structures Signal a Th1 Cytotoxic Immune Contexture and License the Positive Prognostic Value of Infiltrating CD8+ T Cells. Cancer Res. 2014, 74, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Truxova, I.; Kasikova, L.; Hensler, M.; Skapa, P.; Laco, J.; Pecen, L.; Belicova, L.; Praznovec, I.; Halaska, M.J.; Brtnicky, T.; et al. Mature Dendritic Cells Correlate with Favorable Immune Infiltrate and Improved Prognosis in Ovarian Carcinoma Patients. J. Immunother. Cancer 2018, 6, 139. [Google Scholar] [CrossRef] [PubMed]
- Ishigami, S.; Natsugoe, S.; Hokita, S.; Xiangming, C.; Aridome, K.; Iwashige, H.; Tokuda, K.; Nakajo, A.; Miyazono, F.; Aikou, T. Intranodal Antitumor Immunocyte Infiltration in Node-Negative Gastric Cancers. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2000, 6, 2611–2617. [Google Scholar]
- Jensen, T.O.; Schmidt, H.; Møller, H.J.; Donskov, F.; Høyer, M.; Sjoegren, P.; Christensen, I.J.; Steiniche, T. Intratumoral Neutrophils and Plasmacytoid Dendritic Cells Indicate Poor Prognosis and are Associated with pSTAT3 Expression in AJCC Stage I/II Melanoma. Cancer 2012, 118, 2476–2485. [Google Scholar] [CrossRef] [PubMed]
- Dai, F.; Liu, L.; Che, G.; Yu, N.; Pu, Q.; Zhang, S.; Ma, J.; Ma, L.; You, Z. The Number and Microlocalization of Tumor-Associated Immune Cells are Associated with Patient’s Survival Time in Non-Small Cell Lung Cancer. BMC Cancer 2010, 10, 220. [Google Scholar] [CrossRef] [PubMed]
- Gulubova, M.V.; Ananiev, J.R.; Vlaykova, T.I.; Yovchev, Y.; Tsoneva, V.; Manolova, I.M. Role of Dendritic Cells in Progression and Clinical Outcome of Colon Cancer. Int. J. Colorectal. Dis. 2012, 27, 159–169. [Google Scholar] [CrossRef]
- Kobayashi, M.; Suzuki, K.; Yashi, M.; Yuzawa, M.; Takayashiki, N.; Morita, T. Tumor Infiltrating Dendritic Cells Predict Treatment Response to Immmunotherapy in Patients with Metastatic Renal Cell Carcinoma. Anticancer Res. 2007, 27, 1137–1141. [Google Scholar] [PubMed]
- Aso, T.; Ogawa, Y.; Naoe, M.; Fukagai, T.; Yoshida, H.; Kushima, M. Immunohistochemical Analysis of CD83, CD8 and CD4 Positive Cells in Renal Cell Carcinoma. Jpn. J. Urol. 2004, 95, 645–650. [Google Scholar] [CrossRef]
- Bailur, J.; Gueckel, B.; Pawelec, G. Prognostic Impact of High Levels of Circulating Plasmacytoid Dendritic Cells in Breast Cancer. J. Transl. Med. 2016, 14, 151. [Google Scholar] [CrossRef]
- Han, N.; Zhang, Z.; Liu, S.; Ow, A.; Ruan, M.; Yang, W.; Zhang, C. Increased Tumor-Infiltrating Plasmacytoid Dendritic Cells Predicts Poor Prognosis in Oral Squamous Cell Carcinoma. Arch. Oral. Biol. 2017, 78, 129–134. [Google Scholar] [CrossRef]
- O’Donnell, R.K.; Mick, R.; Feldman, M.; Hino, S.; Wang, Y.; Brose, M.S.; Muschel, R.J. Distribution of Dendritic Cell Subtypes in Primary Oral Squamous Cell Carcinoma is Inconsistent with a Functional Response. Cancer Lett. 2007, 255, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Barry, K.C.; Hsu, J.; Broz, M.L.; Cueto, F.J.; Binnewies, M.; Combes, A.J.; Nelson, A.E.; Loo, K.; Kumar, R.; Rosenblum, M.D.; et al. A Natural Killer-Dendritic Cell Axis Defines Checkpoint Therapy-Responsive Tumor Microenvironments. Nat. Med. 2018, 24, 1178–1191. [Google Scholar] [CrossRef] [PubMed]
- Böttcher, J.P.; Bonavita, E.; Chakravarty, P.; Blees, H.; Cabeza-Cabrerizo, M.; Sammicheli, S.; Rogers, N.C.; Sahai, E.; Zelenay, S.; Sousa, C.E. NK Cells Stimulate Recruitment of cDC1 into the Tumor Microenvironment Promoting Cancer Immune Control. Cell 2018, 172, 1022–1037. [Google Scholar] [CrossRef] [PubMed]
- Gentles, A.J.; Newman, A.M.; Liu, C.L.; Bratman, S.V.; Feng, W.; Kim, D.; Nair, V.S.; Xu, Y.; Khuong, A.; Hoang, C.D.; et al. The Prognostic Landscape of Genes and Infiltrating Immune Cells across Human Cancers. Nat. Med. 2015, 21, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Gerlini, G.; Urso, C.; Mariotti, G.; Gennaro, P.; Palli, D.; Brandani, P.; Salvadori, A.; Pimpinelli, N.; Reali, U.; Borgognoni, L. Plasmacytoid Dendritic Cells Represent a Major Dendritic Cell Subset in Sentinel Lymph Nodes of Melanoma Patients and Accumulate in Metastatic Nodes. Clin. Immunol. 2007, 125, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Bekeredjian-Ding, I.; Schäfer, M.; Hartmann, E.; Pries, R.; Parcina, M.; Schneider, P.; Giese, T.; Endres, S.; Wollenberg, B.; Hartmann, G. Tumour-Derived Prostaglandin E2 and Transforming Growth Factor-β Synergize to Inhibit Plasmacytoid Dendritic Cell-Derived Interferon-α. Immunology 2009, 128, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Le Mercier, I.; Poujol, D.; Sanlaville, A.; Sisirak, V.; Gobert, M.; Durand, I.; Dubois, B.; Treilleux, I.; Marvel, J.; Vlach, J.; et al. Tumor Promotion by Intratumoral Plasmacytoid Dendritic Cells is Reversed by TLR7 Ligand Treatment. Cancer Res. 2013, 73, 4629–4640. [Google Scholar] [CrossRef]
- Segura, E.; Kapp, E.; Gupta, N.; Wong, J.; Lim, J.; Ji, H.; Heath, W.R.; Simpson, R.; Villadangos, J.A. Differential Expression of Pathogen-Recognition Molecules between Dendritic Cell Subsets Revealed by Plasma Membrane Proteomic Analysis. Mol. Immunol. 2010, 47, 1765–1773. [Google Scholar] [CrossRef]
- Becker, L.; Liu, N.-C.; Averill, M.M.; Yuan, W.; Pamir, N.; Peng, Y.; Irwin, A.D.; Fu, X.; Bornfeldt, K.E.; Heinecke, J.W. Unique Proteomic Signatures Distinguish Macrophages and Dendritic Cells. PLoS ONE 2012, 7, e33297. [Google Scholar] [CrossRef]
- Worah, K.; Mathan, T.S.; Vu Manh, T.P.; Keerthikumar, S.; Schreibelt, G.; Tel, J.; Boer, T.; Sköld, A.E.; van Spriel, A.B.; de Vries, J.I.; et al. Proteomics of Human Dendritic Cell Subsets Reveals Subset-Specific Surface Markers and Differential Inflammasome Function. Cell Rep. 2016, 16, 2953–2966. [Google Scholar] [CrossRef]
- Korkmaz, A.; Popov, T.; Peisl, L.; Codrea, M.; Nahnsen, S.; Steimle, A.; Velic, A.; Macek, B.; von Bergen, M.; Bernhardt, J.; et al. Proteome and phosphoproteome analysis of commensally induced dendritic cell maturation states. J. Proteom. 2018, 180, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Arya, S.; Wiatrek-Moumoulidis, D.; Synowsky, S.A.; Shirran, S.L.; Botting, C.H.; Powis, S.J.; Stewart, A.J. Quantitative proteomic changes in LPS-activated monocyte-derived dendritic cells: A SWATH-MS study. Sci. Rep. 2019, 9, 4343. [Google Scholar] [CrossRef]
- Ayers, M.; Lunceford, J.; Nebozhyn, M.; Murphy, E.; Loboda, A.; Kaufman, D.R.; Albright, A.; Cheng, J.D.; Kang, S.; Shankaran, V.; et al. IFN-γ-related mRNA profile predicts clinical response to PD-1 blockade. J. Clin. Investig. 2017, 127, 2930–2940. [Google Scholar] [CrossRef]
- Higgs, B.W.; Morehouse, C.; Streicher, K.L.; Brohawn, P.; Pilataxi, F.; Gupta, A.; Ranade, K. Interferon Gamma Messenger RNA Signature in Tumor Biopsies Predicts Outcomes in Patients with Non-Small-Cell Lung Carcinoma or Urothelial Cancer Treated with Durvalumab. Clin. Cancer Res. 2018, 24, 3857–3866. [Google Scholar] [CrossRef] [PubMed]
- Danaher, P.; Warren, S.; Lu, R.; Samayoa, J.; Sullivan, A.; Pekker, I.; Wallden, B.; Marincola, F.M.; Cesano, A. Pan-cancer adaptive immune resistance as defined by the Tumor Inflammation Signature (TIS): Results from The Cancer Genome Atlas (TCGA). J. Immunother. Cancer 2018, 6, 63. [Google Scholar] [CrossRef] [PubMed]
- Matos, L.; Trufelli, D.; Matos, M.; Pinhal, M. Immunohistochemistry as an Important Tool in Biomarkers Detection and Clinical Practice. Biomark. Insights 2010, 5, BMI–S2185. [Google Scholar] [CrossRef]
- Giesen, C.; Wang, H.A.; Schapiro, D.; Zivanovic, N.; Jacobs, A.; Hattendorf, B.; Schüffler, P.J.; Grolimund, D.; Buhmann, J.M.; Brandt, S.; et al. Highly multiplexed imaging of tumor tissues with subcellular resolution by mass cytometry. Nat. Methods 2014, 11, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Angelo, M.; Bendall, S.C.; Finck, R.; Hale, M.B.; Hitzman, C.; Borowsky, A.D.; Levenson, R.M.; Lowe, J.B.; Liu, S.D.; Zhao, S.; et al. Multiplexed ion beam imaging of human breast tumors. Nat. Med. 2014, 20, nm.3488. [Google Scholar] [CrossRef]
- Keren, L.; Bosse, M.; Marquez, D.; Angoshtari, R.; Jain, S.; Varma, S.; Yang, S.-R.; Kurian, A.; Valen, D.; West, R.; et al. A Structured Tumor-Immune Microenvironment in Triple Negative Breast Cancer Revealed by Multiplexed Ion Beam Imaging. Cell 2018, 174, 1373–1387. [Google Scholar] [CrossRef]
- Wang, F.; Flanagan, J.; Su, N.; Wang, L.-C.; Bui, S.; Nielson, A.; Wu, X.; Vo, H.-T.; Ma, X.-J.; Luo, Y. RNAscope A Novel in Situ RNA Analysis Platform for Formalin-Fixed, Paraffin-Embedded Tissues. J. Mol. Diagn. 2012, 14, 22–29. [Google Scholar] [CrossRef]
- Schulz, D.; Zanotelli, V.; Fischer, J.; Schapiro, D.; Engler, S.; Lun, X.-K.; Jackson, H.; Bodenmiller, B. Simultaneous Multiplexed Imaging of mRNA and Proteins with Subcellular Resolution in Breast Cancer Tissue Samples by Mass Cytometry. Cell Syst. 2018, 6, 25–36. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Histology | Biomarkers | DC Subset | Localization | Maturation State | Reference |
|---|---|---|---|---|---|
| Breast cancer | CD1a, CD207, CD83 | DC, LC | Intratumoral | Immature | [116] |
| DC-LAMP, CD11c | cDC | Peritumoral | Mature | ||
| CD1a | LC | Intratumoral | NA | [117] | |
| CD1a | LC | Intratumoral | NA | [118] | |
| CD1a, CMRF-44, CMFR-56, CD83 | cDC, LC | Intra and peritumoral | Immature | [119] | |
| CD1a, CD83 | DC, LC | Intratumoral | Mature | [120] | |
| CD1a | LC | Intratumoral | NA | [121] | |
| CD1a, CD207 | LC | Intratumoral | Immature | [122] | |
| DC-LAMP | DC | Peritumoral | Mature | ||
| CD123 | pDC | Intratumoral | Immature | ||
| BDCA2 | pDC | Intratumoral | NA | [44] | |
| DC-LAMP | DC | Intratumoral | Mature | [123] | |
| Colorectal cancer | CD1a | LC | Intratumoral | Immature | [124] |
| CD83 | cDC | Peritumoral | Mature | ||
| CD1a | LC | Peritumoral | Immature | [125] | |
| DC-LAMP | DC | Intratumoral | Mature | ||
| CD1a, CD207, CD123, DC-LAMP | DC, LC, pDC | Intratumoral | NA or mature | [115] | |
| Lung cancer | CD11c, BDCA2, CD83, Lin- | cDC, pDC | Intratumoral | Immature | [126] |
| DC-LAMP | DC | Tertiary Lymphoid Structure | Mature | [127] | |
| Melanoma | CD1a, CD207, DC-SIGN, CD206 | cDC, LC | NA | NA | [128] |
| BDCA2 | pDC | NA | NA | ||
| CD207, CD1a | LC | NA | NA | [129] | |
| CD1a, DC-LAMP | DC, LC | Intratumoral | Immature | [130] | |
| CD1a, DC-LAMP | DC, LC | Peritumoral | Mature | ||
| CD11c, BDCA3 | cDC1 | NA | NA | [81] | |
| BDCA1 | cDC2 | NA | NA | ||
| Ovarian cancer | BDCA2 | pDC | NA | NA | [131] |
| Kidney cancer | CD80, CD83, CD86, HLA-DR, CMH-I, CD54 | cDC | NA | Mature | [132] |
| CD1a, CD80, CD86, CD83, CMRF-44 | cDC, LC | Intratumoral | Immature | [133] | |
| CD1a, CD40, CD80, CD83, CD86, HLA-DR | cDC, LC | Intratumoral | Mature and Immature | [134] | |
| CD1a | LC | Intratumoral | Immature | [135] | |
| CD83 | DC | Peritumoral | Mature | ||
| Head and Neck cancer | CD1a, HLA-DR | LC | Intratumoral | Mature | [136] |
| LC | Peritumoral | Immature | |||
| CD1a | LC | Intratumoral | Immature | [137] | |
| LC | Peritumoral | Immature | |||
| BDCA2, CD123, HLA-DR | pDC | Intratumoral | Immature | [138] | |
| CD1a | LC | Intratumoral | Immature | [139] | |
| DC-LAMP | DC | Peritumoral | Mature | ||
| Bladder cancer | CD83 | DC | Intratumoral | Mature | [140] |
| Gastric cancer | DC-LAMP | DC | Intra and peritumoral | Mature | [141] |
| Histology | Markers | DC Subset | Maturation State | Reference |
|---|---|---|---|---|
| Breast cancer | Lin− CD4+ CD11c− CD123+ BDCA2+ | pDC | Mature | [44] |
| Lin− CD4+ CD11c+ BDCA1+ | cDC2 | NA | ||
| Colorectal cancer | HLA-DR+ CD11c+ IRF8+ | cDC1 | NA | [80] |
| HLA-DR+ CD11c+ BDCA1+ | cDC2 | NA | ||
| HLA-DR+ CD11c+ CD14+ | MoDC | NA | ||
| Lung cancer | CD1c+ | cDC2/LC | NA | [145] |
| CD123+ BDCA2+ | pDC | NA | ||
| HLA-DR+ CD11c+ IRF8+ | cDC1 | NA | [80] | |
| HLA-DR+ CD11c+ BDCA1+ | cDC2 | NA | ||
| HLA-DR+ CD11c+ CD14+ | MoDC | NA | ||
| Melanoma | HLA-DR+ BDCA2+ | pDC | NA | [146] |
| HLA-DR+ CD11c+ BDCA3+ | cDC1 | NA | [77] | |
| HLA-DR+ CD11c+ BDCA1+ | cDC2 | NA | ||
| Ovarian cancer | Lin− CD4+ CD11c− | pDC | Immature | [147] |
| HLA-DR+ CD4+ CD123+ CD11c− | pDC | Immature | [148] | |
| Lin- CD4+ CD11c− CD123+ BDCA2+ | pDC | Mature | [142] | |
| Lin− CD4+ CD11c+ | cDC | NA |
| Tissues | Technologies | DC Subsets | Reference |
|---|---|---|---|
| Spleen, Liver, Lung | CyTOF | cDC1, cDC2 and pDCs | [15] |
| Blood, Skin, Spleen, Tonsil | CyTOF | cDC1, cDC2, LC (not in blood), pDCs and Axl+ DCs (not in skin) | [149] |
| Clear renal cell carcinoma | CyTOF | DCs, pDCs | [158] |
| Lung adenocarcinoma, normal lung and blood | CyTOF & scRNA-seq | cDC1, cDC2 and pDCs | [159] |
| Blood, Skin | Microarray analysis of FACS-sorted DCs | cDC1, cDC2, pDCs (blood only), BDCA1+ BDCA3+ DCs (skin only) | [51] |
| Gut | Microarray analysis of FACS-sorted DCs | CD103+ SIRP-α+ DCs, CD103− SIRP-α+ DCs, CD103+ SIRP-α− DCs | [13] |
| Blood, Spleen and Tonsil | Microarray analysis of FACS-sorted DCs | cDC1, cDC2 and pDCs | [152] |
| Blood | RNA-seq | cDC2 and MoDCs | [114] |
| Breast carcinoma | RNA-seq | cDC1-enriched cells, cDC2, pDCs and MoDCs | [143] |
| Blood | scRNA-seq of FACS-sorted DCs | cDC1, 2 clusters of cDC2, pDC and Axl+ DCs | [17] |
| Blood | scRNA-seq of FACS-sorted HLA-DR+ CD135+ cells | cDC1, 2 clusters of cDC2, pDC and Axl+ DCs | [16,96] |
| Melanoma-draining lymph nodes | scRNA-seq of HLA-DR+ cells | cDC1, 3 clusters of cDC2 and mature DCs | [96] |
| Lung adenocarcinoma | scRNA-seq | cDC1, cDC2, pDCs and mature DCs | [160] |
| Marker/Population | Prognostic Impact | Histology | Reference |
|---|---|---|---|
| CD1a | Positive | Breast | [117] |
| [120] | |||
| [121] | |||
| Head and Neck | [165] | ||
| [166] | |||
| Lung | [167] | ||
| Melanoma | [130] | ||
| Negative | Osteosarcoma | [168] | |
| Biliar tracts | [169] | ||
| Colon | [125] | ||
| None | Breast | [122] | |
| Kidney | [134] | ||
| [135] | |||
| [137] | |||
| DC-LAMP | Positive | Melanoma | [130] |
| [129] | |||
| Lung | [170] | ||
| [171] | |||
| [127] | |||
| Ovary | [172] | ||
| Breast | [123] | ||
| Negative | Colon | [125] | |
| Gastric | [173] | ||
| None | Melanoma | [174] | |
| Breast | [122] | ||
| CD83 | Positive | Breast | [120] |
| Lung | [175] | ||
| Colon | [176] | ||
| Kidney | [177] | ||
| Negative | Kidney | [135] | |
| Kidney | [134] | ||
| [178] | |||
| cDC2 (BDCA1) | Negative | Lung | [145] |
| pDC (BDCA2, CD123) | Positive | Breast | [179] |
| Negative | Breast | [122] | |
| [44] | |||
| Ovary | [131] | ||
| [142] | |||
| Head and Neck | [180] | ||
| [181] | |||
| Melanoma | [174] | ||
| [146] | |||
| None | Melanoma | [128] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hubert, M.; Gobbini, E.; Bendriss-Vermare, N.; Caux, C.; Valladeau-Guilemond, J. Human Tumor-Infiltrating Dendritic Cells: From In Situ Visualization to High-Dimensional Analyses. Cancers 2019, 11, 1082. https://doi.org/10.3390/cancers11081082
Hubert M, Gobbini E, Bendriss-Vermare N, Caux C, Valladeau-Guilemond J. Human Tumor-Infiltrating Dendritic Cells: From In Situ Visualization to High-Dimensional Analyses. Cancers. 2019; 11(8):1082. https://doi.org/10.3390/cancers11081082
Chicago/Turabian StyleHubert, Margaux, Elisa Gobbini, Nathalie Bendriss-Vermare, Christophe Caux, and Jenny Valladeau-Guilemond. 2019. "Human Tumor-Infiltrating Dendritic Cells: From In Situ Visualization to High-Dimensional Analyses" Cancers 11, no. 8: 1082. https://doi.org/10.3390/cancers11081082
APA StyleHubert, M., Gobbini, E., Bendriss-Vermare, N., Caux, C., & Valladeau-Guilemond, J. (2019). Human Tumor-Infiltrating Dendritic Cells: From In Situ Visualization to High-Dimensional Analyses. Cancers, 11(8), 1082. https://doi.org/10.3390/cancers11081082