Deciphering The Potential Role of Hox Genes in Pancreatic Cancer

Abstract
1. Introduction
2. Important Features of Hox Genes
3. Involvement of The Hox Genes in Pancreas Development
4. Dysregulation of Hox Gene Expression in Pancreatic Cancer
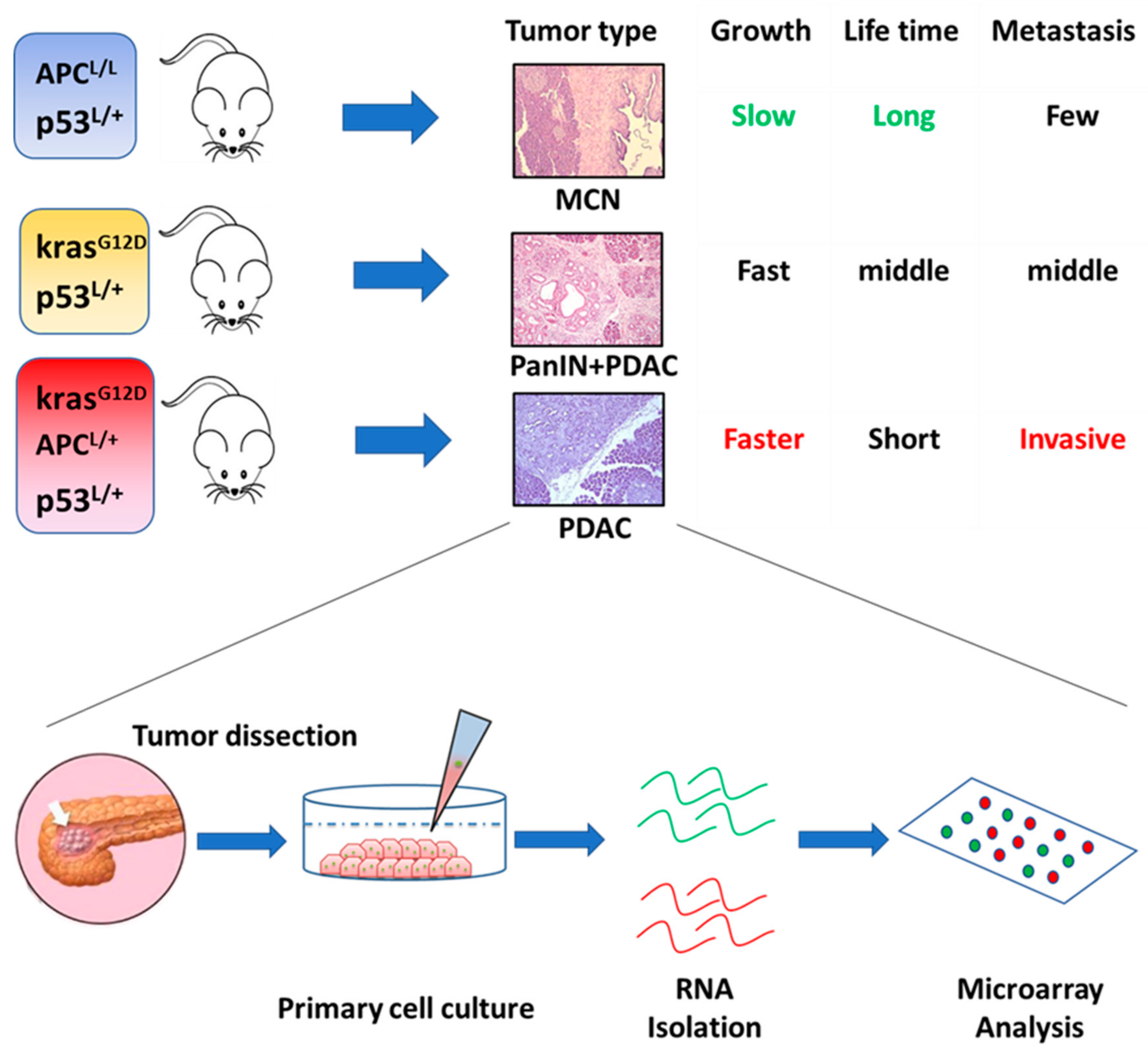
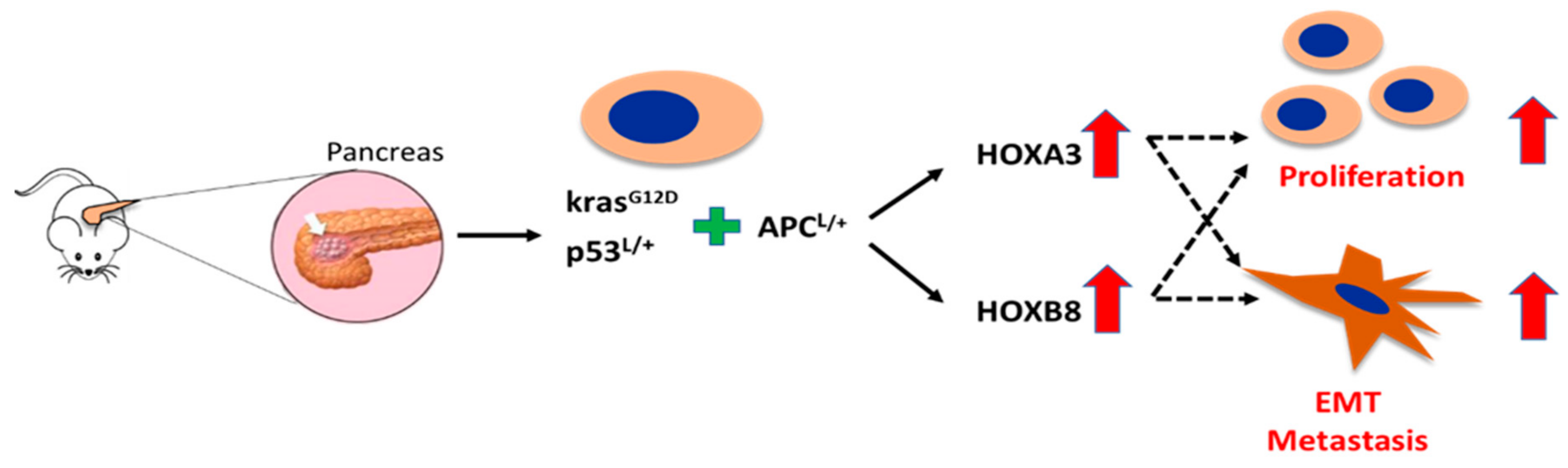
5. Novel Hox Genes in Pancreatic Tumorigenesis Identified by Genetically Engineered Mouse (GEM) Models
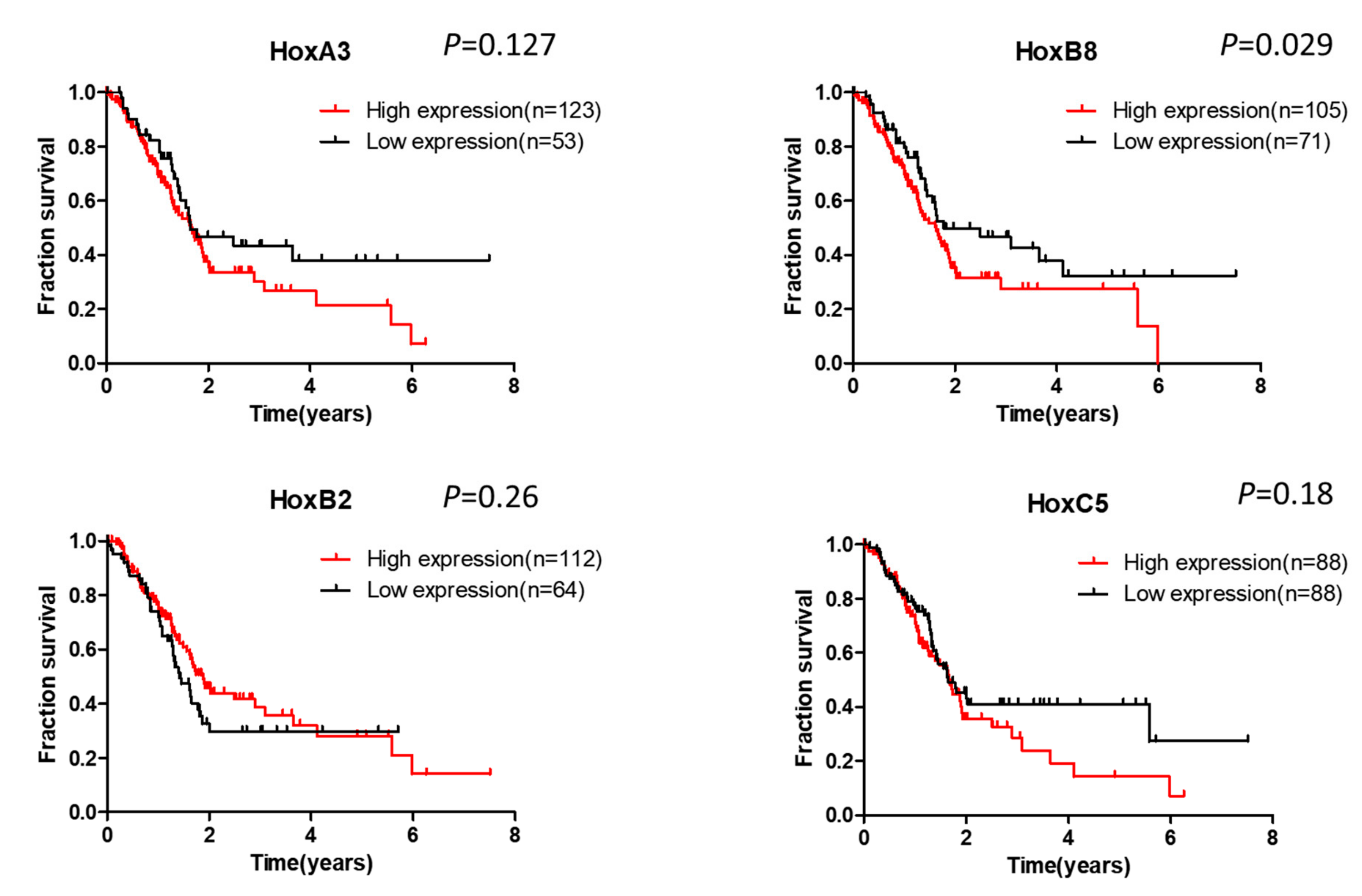
6. Clinical Significance of Hox Gene Dysregulation in Pancreatic Cancer
7. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lewis, E.B. A gene complex controlling segmentation in Drosophila. Nature 1978, 276, 565–570. [Google Scholar] [CrossRef]
- Regulski, M.; McGinnis, N.; Chadwick, R.; McGinnis, W. Developmental and molecular analysis of Deformed; a homeotic gene controlling Drosophila head development. EMBO J. 1987, 6, 767–777. [Google Scholar] [CrossRef]
- Vachon, G.; Cohen, B.; Pfeifle, C.; McGuffin, M.E.; Botas, J.; Cohen, S.M. Homeotic genes of the Bithorax complex repress limb development in the abdomen of the Drosophila embryo through the target gene Distal-less. Cell 1992, 71, 437–450. [Google Scholar] [CrossRef]
- Schneuwly, S.; Kuroiwa, A.; Baumgartner, P.; Gehring, W.J. Structural organization and sequence of the homeotic gene Antennapedia of Drosophila melanogaster. EMBO J. 1986, 5, 733–739. [Google Scholar] [CrossRef]
- Duboule, D.; Dolle, P. The structural and functional organization of the murine Hox gene family resembles that of Drosophila homeotic genes. EMBO J. 1989, 8, 1497–1505. [Google Scholar] [CrossRef]
- Graham, A.; Papalopulu, N.; Krumlauf, R. The murine and Drosophila homeobox gene complexes have common features of organization and expression. Cell 1989, 57, 367–378. [Google Scholar] [CrossRef]
- Santini, S.; Boore, J.L.; Meyer, A. Evolutionary conservation of regulatory elements in vertebrate Hox gene clusters. Genome Res. 2003, 13, 1111–1122. [Google Scholar] [CrossRef]
- Kim, C.B.; Amemiya, C.; Bailey, W.; Kawasaki, K.; Mezey, J.; Miller, W.; Minoshima, S.; Shimizu, N.; Wagner, G.; Ruddle, F. Hox cluster genomics in the horn shark, Heterodontus francisci. Proc. Natl. Acad. Sci. USA 2000, 97, 1655–1660. [Google Scholar] [CrossRef]
- Crow, K.D.; Stadler, P.F.; Lynch, V.J.; Amemiya, C.; Wagner, G.P. The “fish-specific” Hox cluster duplication is coincident with the origin of teleosts. Mol. Biol. Evol. 2006, 23, 121–136. [Google Scholar] [CrossRef]
- Scott, M.P. A rational nomenclature for vertebrate homeobox (Hox) genes. Nucleic Acids Res. 1993, 21, 1687–1688. [Google Scholar] [CrossRef][Green Version]
- Iimura, T.; Pourquie, O. Hox genes in time and space during vertebrate body formation. Dev. Growth Differ. 2007, 49, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Hoegg, S.; Meyer, A. Hox clusters as models for vertebrate genome evolution. Trends Genet. 2005, 21, 421–424. [Google Scholar] [CrossRef]
- Kawazoe, Y.; Sekimoto, T.; Araki, M.; Takagi, K.; Araki, K.; Yamamura, K. Region-specific gastrointestinal Hox code during murine embryonal gut development. Dev. Growth Differ. 2002, 44, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Larsen, B.M.; Hrycaj, S.M.; Newman, M.; Li, Y.; Wellik, D.M. Mesenchymal Hox6 function is required for mouse pancreatic endocrine cell differentiation. Development 2015, 142, 3859–3868. [Google Scholar] [CrossRef]
- Almgren, P.; Lindqvist, A.; Krus, U.; Hakaste, L.; Ottosson-Laakso, E.; Asplund, O.; Sonestedt, E.; Prasad, R.B.; Laurila, E.; Orho-Melander, M.; et al. Genetic determinants of circulating GIP and GLP-1 concentrations. JCI Insight 2017, 2. [Google Scholar] [CrossRef]
- Tehrani, Z.; Lin, S. Antagonistic interactions of hedgehog, Bmp and retinoic acid signals control zebrafish endocrine pancreas development. Development 2011, 138, 631–640. [Google Scholar] [CrossRef]
- Shi, Y. Generation of functional insulin-producing cells from human embryonic stem cells in vitro. Methods Mol. Biol. 2010, 636, 79–85. [Google Scholar] [CrossRef]
- Lomberk, G.A.; Imoto, I.; Gebelein, B.; Urrutia, R.; Cook, T.A. Conservation of the TGFbeta/Labial homeobox signaling loop in endoderm-derived cells between Drosophila and mammals. Pancreatology 2010, 10, 74–84. [Google Scholar] [CrossRef][Green Version]
- Sosa-Pineda, B. The gene Pax4 is an essential regulator of pancreatic beta-cell development. Mol. Cells 2004, 18, 289–294. [Google Scholar]
- Sosa-Pineda, B.; Chowdhury, K.; Torres, M.; Oliver, G.; Gruss, P. The Pax4 gene is essential for differentiation of insulin-producing beta cells in the mammalian pancreas. Nature 1997, 386, 399–402. [Google Scholar] [CrossRef]
- Gonzalez, S.; Binato, R.; Guida, L.; Mencalha, A.L.; Abdelhay, E. Conserved transcription factor binding sites suggest an activator basal promoter and a distal inhibitor in the galanin gene promoter in mouse ES cells. Gene 2014, 538, 228–234. [Google Scholar] [CrossRef]
- Westerman, B.A.; Murre, C.; Oudejans, C.B. The cellular Pax-Hox-helix connection. Biochim. Biophys. Acta 2003, 1629, 1–7. [Google Scholar] [CrossRef]
- Kanai, M.; Hamada, J.; Takada, M.; Asano, T.; Murakawa, K.; Takahashi, Y.; Murai, T.; Tada, M.; Miyamoto, M.; Kondo, S.; et al. Aberrant expressions of Hox genes in colorectal and hepatocellular carcinomas. Oncol. Rep. 2010, 23, 843–851. [Google Scholar]
- Cantile, M.; Pettinato, G.; Procino, A.; Feliciello, I.; Cindolo, L.; Cillo, C. In vivo expression of the whole HOX gene network in human breast cancer. Eur. J. Cancer 2003, 39, 257–264. [Google Scholar] [CrossRef]
- Miller, G.J.; Miller, H.L.; van Bokhoven, A.; Lambert, J.R.; Werahera, P.N.; Schirripa, O.; Lucia, M.S.; Nordeen, S.K. Aberrant HOXC expression accompanies the malignant phenotype in human prostate. Cancer Res. 2003, 63, 5879–5888. [Google Scholar]
- Abe, M.; Hamada, J.; Takahashi, O.; Takahashi, Y.; Tada, M.; Miyamoto, M.; Morikawa, T.; Kondo, S.; Moriuchi, T. Disordered expression of Hox genes in human non-small cell lung cancer. Oncol. Rep. 2006, 15, 797–802. [Google Scholar] [CrossRef]
- Costa, B.M.; Smith, J.S.; Chen, Y.; Chen, J.; Phillips, H.S.; Aldape, K.D.; Zardo, G.; Nigro, J.; James, C.D.; Fridlyand, J.; et al. Reversing HOXA9 oncogene activation by PI3K inhibition: Epigenetic mechanism and prognostic significance in human glioblastoma. Cancer Res. 2010, 70, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Liu, J.; Yoshida, H.; Rosen, D.; Naora, H. Lineage infidelity of epithelial ovarian cancers is controlled by Hox genes that specify regional identity in the reproductive tract. Nat. Med. 2005, 11, 531–537. [Google Scholar] [CrossRef]
- Argiropoulos, B.; Humphries, R.K. Hox genes in hematopoiesis and leukemogenesis. Oncogene 2007, 26, 6766–6776. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Chen, H.; Parker, B.; Rubin, E.; Zhu, T.; Lee, J.S.; Argani, P.; Sukumar, S. HOXB7, a homeodomain protein, is overexpressed in breast cancer and confers epithelial-mesenchymal transition. Cancer Res. 2006, 66, 9527–9534. [Google Scholar] [CrossRef]
- Zhuang, L.; Li, W.H.; Li, K.; Mao, Y.; Gao, C.L.; Zhang, C. HoxB7 Promotes Growth and Metastasis of Lung Adenocarcinoma Cells Through Regulation of the Tgf-Beta/Smad3 Signaling. J. Biol. Regul. Homeost. Agents 2015, 29, 601–608. [Google Scholar] [PubMed]
- Yamashita, T.; Tazawa, S.; Yawei, Z.; Katayama, H.; Kato, Y.; Nishiwaki, K.; Yokohama, Y.; Ishikawa, M. Suppression of invasive characteristics by antisense introduction of overexpressed Hox genes in ovarian cancer cells. Int. J. Oncol. 2006, 28, 931–938. [Google Scholar] [CrossRef][Green Version]
- Nguyen Kovochich, A.; Arensman, M.; Lay, A.R.; Rao, N.P.; Donahue, T.; Li, X.; French, S.W.; Dawson, D.W. HOXB7 promotes invasion and predicts survival in pancreatic adenocarcinoma. Cancer 2013, 119, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Weiss, F.U.; Marques, I.J.; Woltering, J.M.; Vlecken, D.H.; Aghdassi, A.; Partecke, L.I.; Heidecke, C.D.; Lerch, M.M.; Bagowski, C.P. Retinoic acid receptor antagonists inhibit miR-10a expression and block metastatic behavior of pancreatic cancer. Gastroenterology 2009, 137, 2136–2145. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.L.; Wu, Y.; Cai, C.Y.; Tang, Z.G. Overexpression of homeobox B-13 correlates with angiogenesis, aberrant expression of EMT markers, aggressive characteristics and poor prognosis in pancreatic carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 6919–6927. [Google Scholar] [PubMed]
- Segara, D.; Biankin, A.V.; Kench, J.G.; Langusch, C.C.; Dawson, A.C.; Skalicky, D.A.; Gotley, D.C.; Coleman, M.J.; Sutherland, R.L.; Henshall, S.M. Expression of HOXB2, a retinoic acid signaling target in pancreatic cancer and pancreatic intraepithelial neoplasia. Clin. Cancer Res. 2005, 11, 3587–3596. [Google Scholar] [CrossRef] [PubMed]
- Asuthkar, S.; Stepanova, V.; Lebedeva, T.; Holterman, A.L.; Estes, N.; Cines, D.B.; Rao, J.S.; Gondi, C.S. Multifunctional roles of urokinase plasminogen activator (uPA) in cancer stemness and chemoresistance of pancreatic cancer. Mol. Biol. Cell 2013, 24, 2620–2632. [Google Scholar] [CrossRef]
- Fu, Z.; Chen, C.; Zhou, Q.; Wang, Y.; Zhao, Y.; Zhao, X.; Li, W.; Zheng, S.; Ye, H.; Wang, L.; et al. LncRNA HOTTIP modulates cancer stem cell properties in human pancreatic cancer by regulating HOXA9. Cancer Lett. 2017, 410, 68–81. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, X.; Zhou, Y.; Liu, Y.; Zhou, Q.; Ye, H.; Wang, Y.; Zeng, J.; Song, Y.; Gao, W.; et al. The long non-coding RNA HOTTIP promotes progression and gemcitabine resistance by regulating HOXA13 in pancreatic cancer. J. Transl. Med. 2015, 13, 84. [Google Scholar] [CrossRef]
- Makiyama, K.; Hamada, J.; Takada, M.; Murakawa, K.; Takahashi, Y.; Tada, M.; Tamoto, E.; Shindo, G.; Matsunaga, A.; Teramoto, K.; et al. Aberrant expression of Hox genes in human invasive breast carcinoma. Oncol. Rep. 2005, 13, 673–679. [Google Scholar] [CrossRef][Green Version]
- Maeda, K.; Hamada, J.; Takahashi, Y.; Tada, M.; Yamamoto, Y.; Sugihara, T.; Moriuchi, T. Altered expressions of Hox genes in human cutaneous malignant melanoma. Int. J. Cancer 2005, 114, 436–441. [Google Scholar] [CrossRef]
- Lopez, R.; Garrido, E.; Vazquez, G.; Pina, P.; Perez, C.; Alvarado, I.; Salcedo, M. A subgroup of HOX Abd-B gene is differentially expressed in cervical cancer. Int. J. Gynecol. Cancer 2006, 16, 1289–1296. [Google Scholar] [CrossRef]
- Abdel-Fattah, R.; Xiao, A.; Bomgardner, D.; Pease, C.S.; Lopes, M.B.; Hussaini, I.M. Differential expression of Hox genes in neoplastic and non-neoplastic human astrocytes. J. Pathol. 2006, 209, 15–24. [Google Scholar] [CrossRef]
- Cantile, M.; Franco, R.; Tschan, A.; Baumhoer, D.; Zlobec, I.; Schiavo, G.; Forte, I.; Bihl, M.; Liguori, G.; Botti, G.; et al. HOX D13 expression across 79 tumor tissue types. Int. J. Cancer 2009, 125, 1532–1541. [Google Scholar] [CrossRef]
- Ohuchida, K.; Mizumoto, K.; Lin, C.; Yamaguchi, H.; Ohtsuka, T.; Sato, N.; Toma, H.; Nakamura, M.; Nagai, E.; Hashizume, M.; et al. MicroRNA-10a is overexpressed in human pancreatic cancer and involved in its invasiveness partially via suppression of the HOXA1 gene. Ann. Surg. Oncol. 2012, 19, 2394–2402. [Google Scholar] [CrossRef]
- Almoguera, C.; Shibata, D.; Forrester, K.; Martin, J.; Arnheim, N.; Perucho, M. Most human carcinomas of the exocrine pancreas contain mutant c-K-ras genes. Cell 1988, 53, 549–554. [Google Scholar] [CrossRef]
- Biankin, A.V.; Waddell, N.; Kassahn, K.S.; Gingras, M.C.; Muthuswamy, L.B.; Johns, A.L.; Miller, D.K.; Wilson, P.J.; Patch, A.M.; Wu, J.; et al. Pancreatic cancer genomes reveal aberrations in axon guidance pathway genes. Nature 2012, 491, 399–405. [Google Scholar] [CrossRef]
- Morton, J.P.; Timpson, P.; Karim, S.A.; Ridgway, R.A.; Athineos, D.; Doyle, B.; Jamieson, N.B.; Oien, K.A.; Lowy, A.M.; Brunton, V.G.; et al. Mutant p53 drives metastasis and overcomes growth arrest/senescence in pancreatic cancer. Proc. Natl. Acad. Sci. USA 2010, 107, 246–251. [Google Scholar] [CrossRef]
- Bardeesy, N.; Aguirre, A.J.; Chu, G.C.; Cheng, K.H.; Lopez, L.V.; Hezel, A.F.; Feng, B.; Brennan, C.; Weissleder, R.; Mahmood, U.; et al. Both p16(Ink4a) and the p19(Arf)-p53 pathway constrain progression of pancreatic adenocarcinoma in the mouse. Proc. Natl. Acad. Sci. USA 2006, 103, 5947–5952. [Google Scholar] [CrossRef]
- Bardeesy, N.; Cheng, K.H.; Berger, J.H.; Chu, G.C.; Pahler, J.; Olson, P.; Hezel, A.F.; Horner, J.; Lauwers, G.Y.; Hanahan, D.; et al. Smad4 is dispensable for normal pancreas development yet critical in progression and tumor biology of pancreas cancer. Genes Dev. 2006, 20, 3130–3146. [Google Scholar] [CrossRef]
- Kuo, T.L.; Weng, C.C.; Kuo, K.K.; Chen, C.Y.; Wu, D.C.; Hung, W.C.; Cheng, K.H. APC haploinsufficiency coupled with p53 loss sufficiently induces mucinous cystic neoplasms and invasive pancreatic carcinoma in mice. Oncogene 2016, 35, 2223–2234. [Google Scholar] [CrossRef]
- Kuo, T.L.; Cheng, K.H.; Shan, Y.S.; Chen, L.T.; Hung, W.C. Beta-catenin-activated autocrine PDGF/Src signaling is a therapeutic target in pancreatic cancer. Theranostics 2019, 9, 324–336. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, G.; Ding, L.; Jiang, T.; Shao, S.; Gao, Y.; Lu, Y. HOXA3 promotes tumor growth of human colon cancer through activating EGFR/Ras/Raf/MEK/ERK signaling pathway. J. Cell. Biochem. 2018, 119, 2864–2874. [Google Scholar] [CrossRef]
- Jiang, L.; Wu, Z.; Meng, X.; Chu, X.; Huang, H.; Xu, C. LncRNA HOXA-AS2 Facilitates Tumorigenesis and Progression of Papillary Thyroid Cancer by Modulating the miR-15a-5p/HOXA3 Axis. Hum. Gene Ther. 2019. [Google Scholar] [CrossRef]
- Vider, B.Z.; Zimber, A.; Hirsch, D.; Estlein, D.; Chastre, E.; Prevot, S.; Gespach, C.; Yaniv, A.; Gazit, A. Human colorectal carcinogenesis is associated with deregulation of homeobox gene expression. Biochem. Biophys. Res. Commun. 1997, 232, 742–748. [Google Scholar] [CrossRef]
- Fujino, T.; Yamazaki, Y.; Largaespada, D.A.; Jenkins, N.A.; Copeland, N.G.; Hirokawa, K.; Nakamura, T. Inhibition of myeloid differentiation by Hoxa9, Hoxb8, and Meis homeobox genes. Exp. Hematol. 2001, 29, 856–863. [Google Scholar] [CrossRef]
- Wang, T.; Lin, F.; Sun, X.; Jiang, L.; Mao, R.; Zhou, S.; Shang, W.; Bi, R.; Lu, F.; Li, S. HOXB8 enhances the proliferation and metastasis of colorectal cancer cells by promoting EMT via STAT3 activation. Cancer Cell Int. 2019, 19, 3. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.J.; Zhou, M.; Chen, M.M.; Qu, C.Y. HOXB8 promotes tumor metastasis and the epithelial-mesenchymal transition via ZEB2 targets in gastric cancer. J. Cancer Res. Clin. Oncol. 2017, 143, 385–397. [Google Scholar] [CrossRef]
- Yuan, Y.; Sun, S.; Jiao, N.; Shu, Y.; Zhang, Y. Upregulation of HOXA10 Protein Expression Predicts Poor Prognosis for Colorectal Cancer. Genet. Test. Mol. Biomark. 2018, 22, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, T.T.; Zhai, G.Q.; Guo, X.Y.; Qin, Y.; Gan, T.Q.; Zhang, Y.; Chen, G.; Mo, W.J.; Feng, Z.B. Evaluation of the HOXA11 level in patients with lung squamous cancer and insights into potential molecular pathways via bioinformatics analysis. World J. Surg. Oncol. 2018, 16, 109. [Google Scholar] [CrossRef] [PubMed]
- Johng, D.; Torga, G.; Ewing, C.M.; Jin, K.; Norris, J.D.; McDonnell, D.P.; Isaacs, W.B. HOXB13 interaction with MEIS1 modifies proliferation and gene expression in prostate cancer. Prostate 2019, 79, 414–424. [Google Scholar] [CrossRef]
- Shen, Z.H.; Zhao, K.M.; Du, T. HOXA10 promotes nasopharyngeal carcinoma cell proliferation and invasion via inducing the expression of ZIC2. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 945–952. [Google Scholar]
- Chu, M.C.; Selam, F.B.; Taylor, H.S. HOXA10 regulates p53 expression and matrigel invasion in human breast cancer cells. Cancer Biol. Ther. 2004, 3, 568–572. [Google Scholar] [CrossRef]
- Sentani, K.; Oue, N.; Naito, Y.; Sakamoto, N.; Anami, K.; Oo, H.Z.; Uraoka, N.; Aoyagi, K.; Sasaki, H.; Yasui, W. Upregulation of HOXA10 in gastric cancer with the intestinal mucin phenotype: Reduction during tumor progression and favorable prognosis. Carcinogenesis 2012, 33, 1081–1088. [Google Scholar] [CrossRef]
- Zhang, L.; Wan, Y.; Jiang, Y.; Ma, J.; Liu, J.; Tang, W.; Wang, X.; Cheng, W. Upregulation HOXA10 homeobox gene in endometrial cancer: Role in cell cycle regulation. Med. Oncol. 2014, 31, 52. [Google Scholar] [CrossRef]
- Han, Y.; Lu, S.; Wen, Y.G.; Yu, F.D.; Zhu, X.W.; Qiu, G.Q.; Tang, H.M.; Peng, Z.H.; Zhou, C.Z. Overexpression of HOXA10 promotes gastric cancer cells proliferation and HOXA10(+)/CD44(+) is potential prognostic biomarker for gastric cancer. Eur. J. Cell Biol. 2015, 94, 642–652. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| KPC/PA53 | ||
|---|---|---|
| Gene Name | Fold Change (log2) | p-Value |
| HoxB2 | 3.508 | <0.001 |
| HoxC5 | 2.940 | 0.007 |
| HoxB8 | 2.402 | <0.001 |
| HoxA3 | 2.185 | <0.001 |
| HoxB9 | 2.098 | <0.001 |
| HoxA1 | 1.867 | <0.001 |
| HoxB3 | 1.249 | 0.038 |
| HoxB13 | −2.188 | 0.028 |
| HoxA10 | −2.605 | <0.001 |
| HoxA11 | −4.501 | <0.001 |
| KPA/KPC | ||
|---|---|---|
| Gene Name | Fold Change (log2) | p-Value |
| HoxA3 | 5.297 | <0.001 |
| HoxB4 | 1.939 | <0.001 |
| HoxB8 | 1.421 | <0.001 |
| HoxA2 | 1.077 | 0.031 |
| HoxA11 | −1.278 | 0.014 |
| HoxA10 | −2.519 | <0.001 |
| HoxB13 | −3.178 | <0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuo, T.-L.; Cheng, K.-H.; Chen, L.-T.; Hung, W.-C. Deciphering The Potential Role of Hox Genes in Pancreatic Cancer. Cancers 2019, 11, 734. https://doi.org/10.3390/cancers11050734
Kuo T-L, Cheng K-H, Chen L-T, Hung W-C. Deciphering The Potential Role of Hox Genes in Pancreatic Cancer. Cancers. 2019; 11(5):734. https://doi.org/10.3390/cancers11050734
Chicago/Turabian StyleKuo, Tzu-Lei, Kuang-Hung Cheng, Li-Tzong Chen, and Wen-Chun Hung. 2019. "Deciphering The Potential Role of Hox Genes in Pancreatic Cancer" Cancers 11, no. 5: 734. https://doi.org/10.3390/cancers11050734
APA StyleKuo, T.-L., Cheng, K.-H., Chen, L.-T., & Hung, W.-C. (2019). Deciphering The Potential Role of Hox Genes in Pancreatic Cancer. Cancers, 11(5), 734. https://doi.org/10.3390/cancers11050734