Mifepristone Overcomes Tumor Resistance to Temozolomide Associated with DNA Damage Repair and Apoptosis in an Orthotopic Model of Glioblastoma

 , , and
, , and 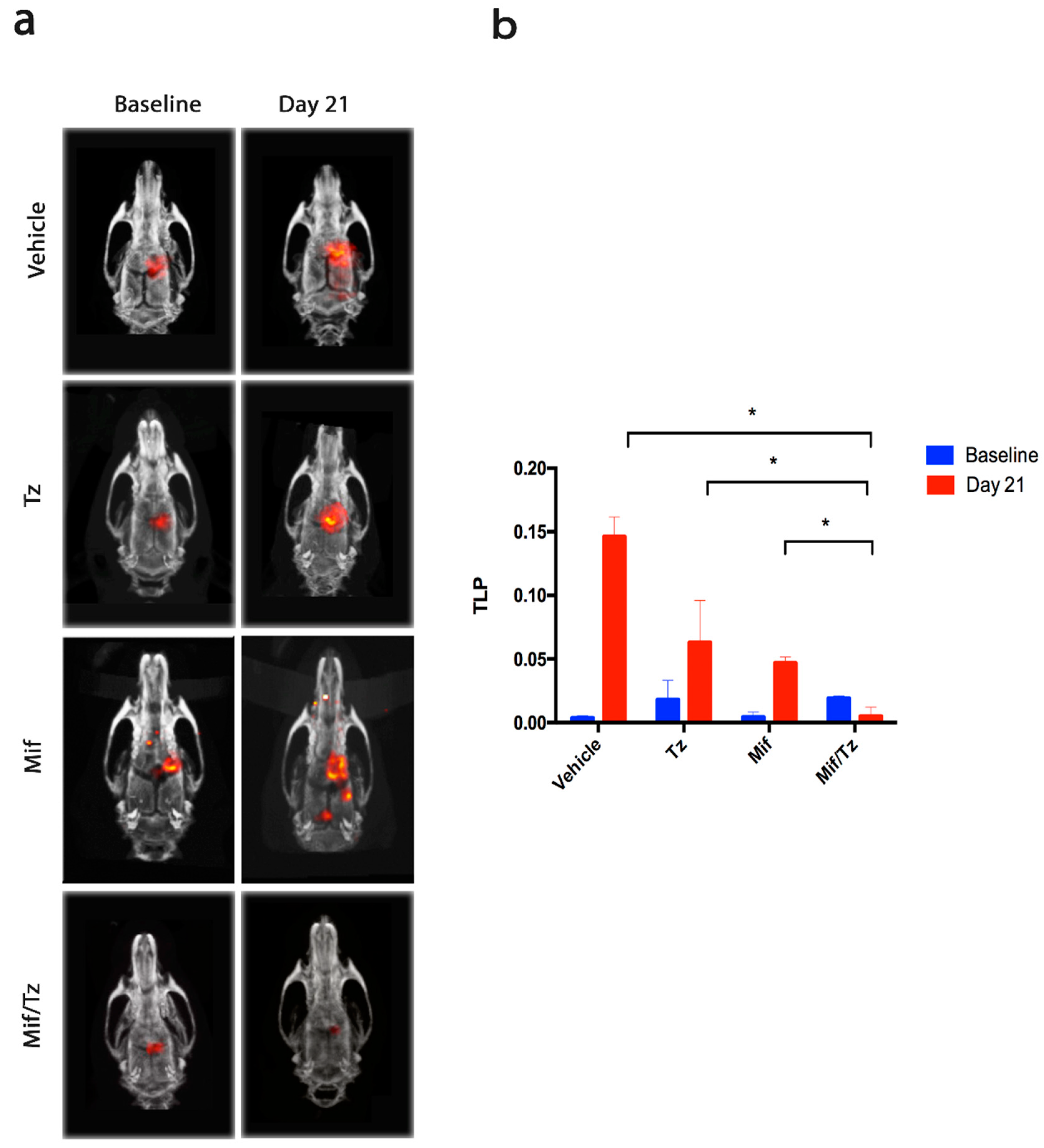{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Tumor Growth Assessed by Molecular Imaging
2.2. Determination of Body Weight and Overall Survival
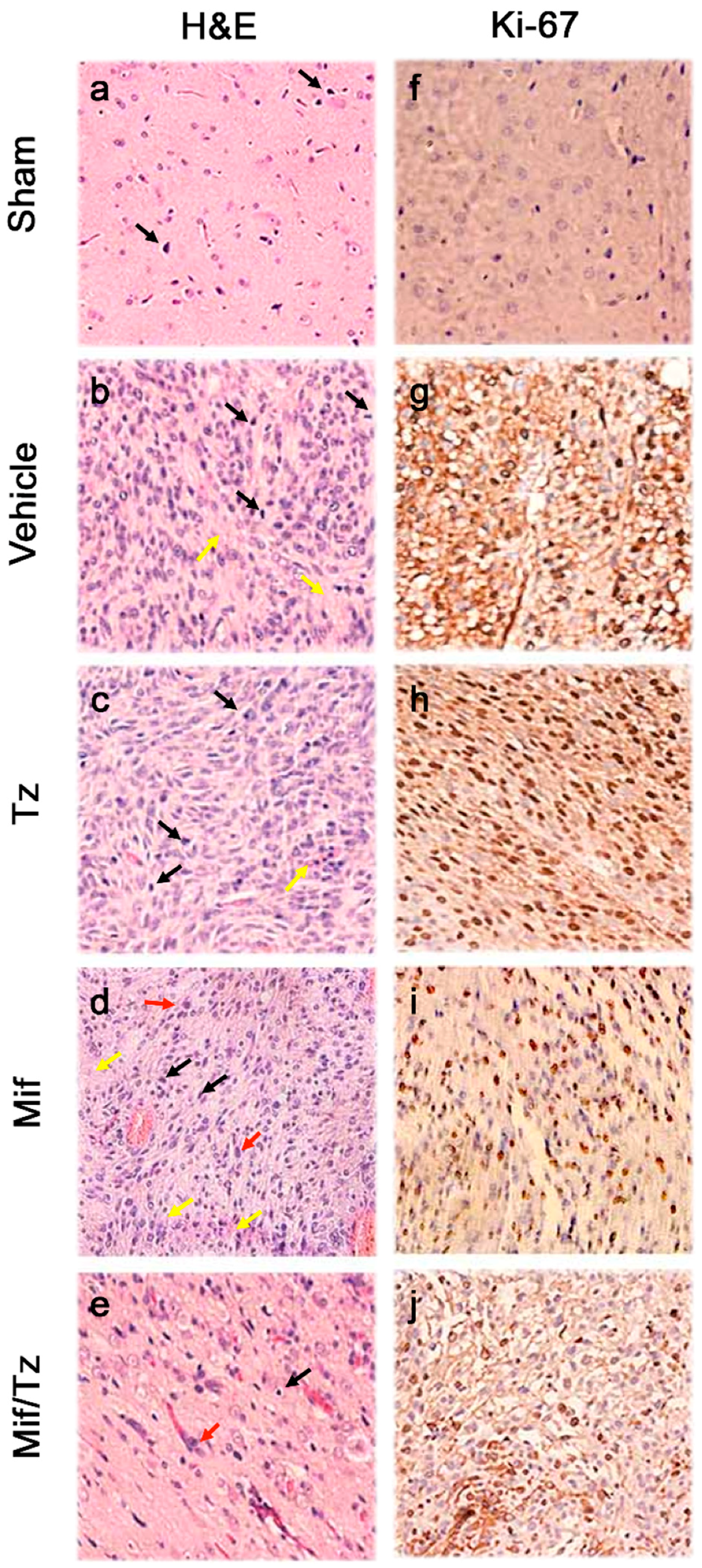
2.3. Histological and Immunohistochemical Analysis
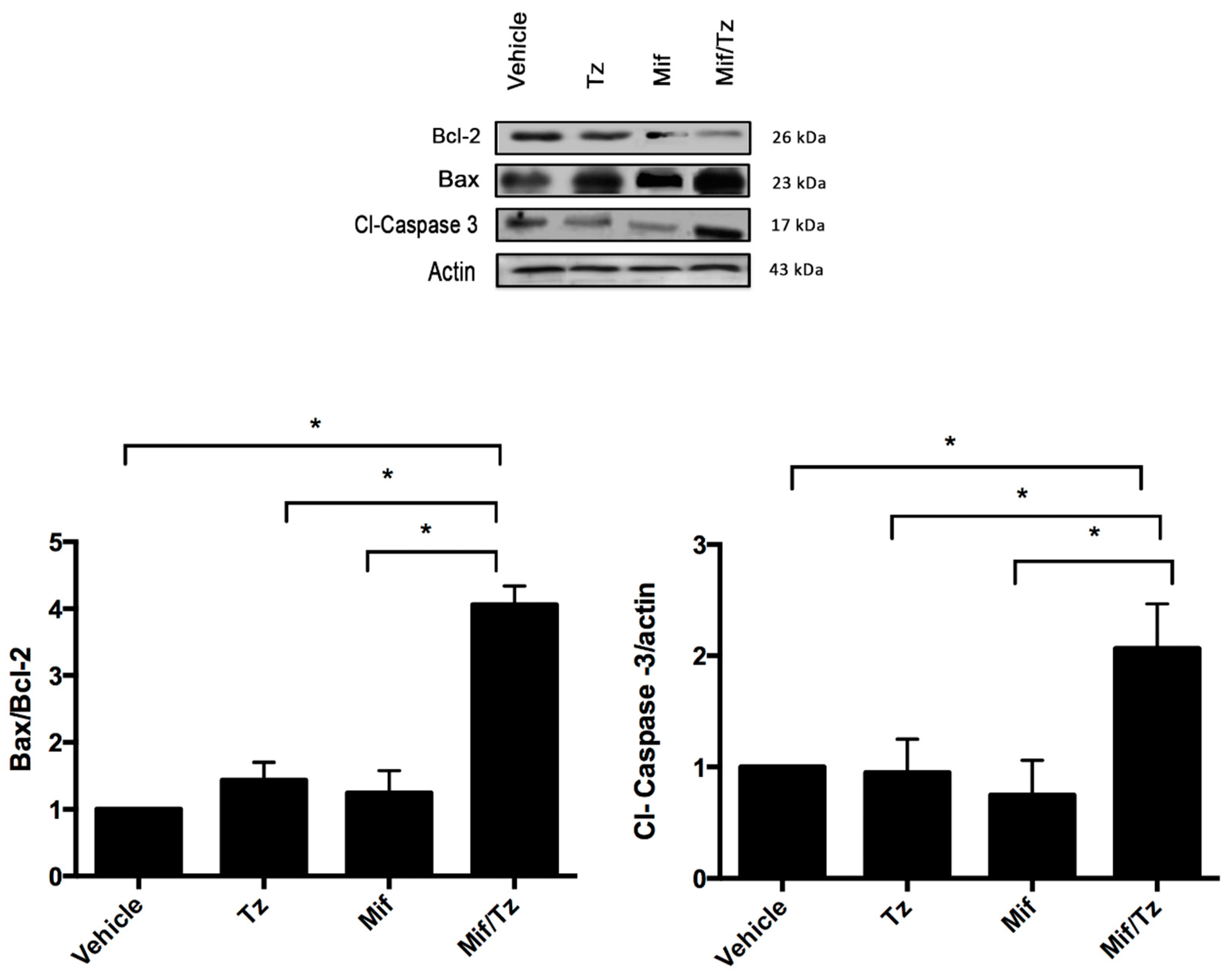
2.4. Expression of Apoptotic Proteins
2.5. Expression of MGMT
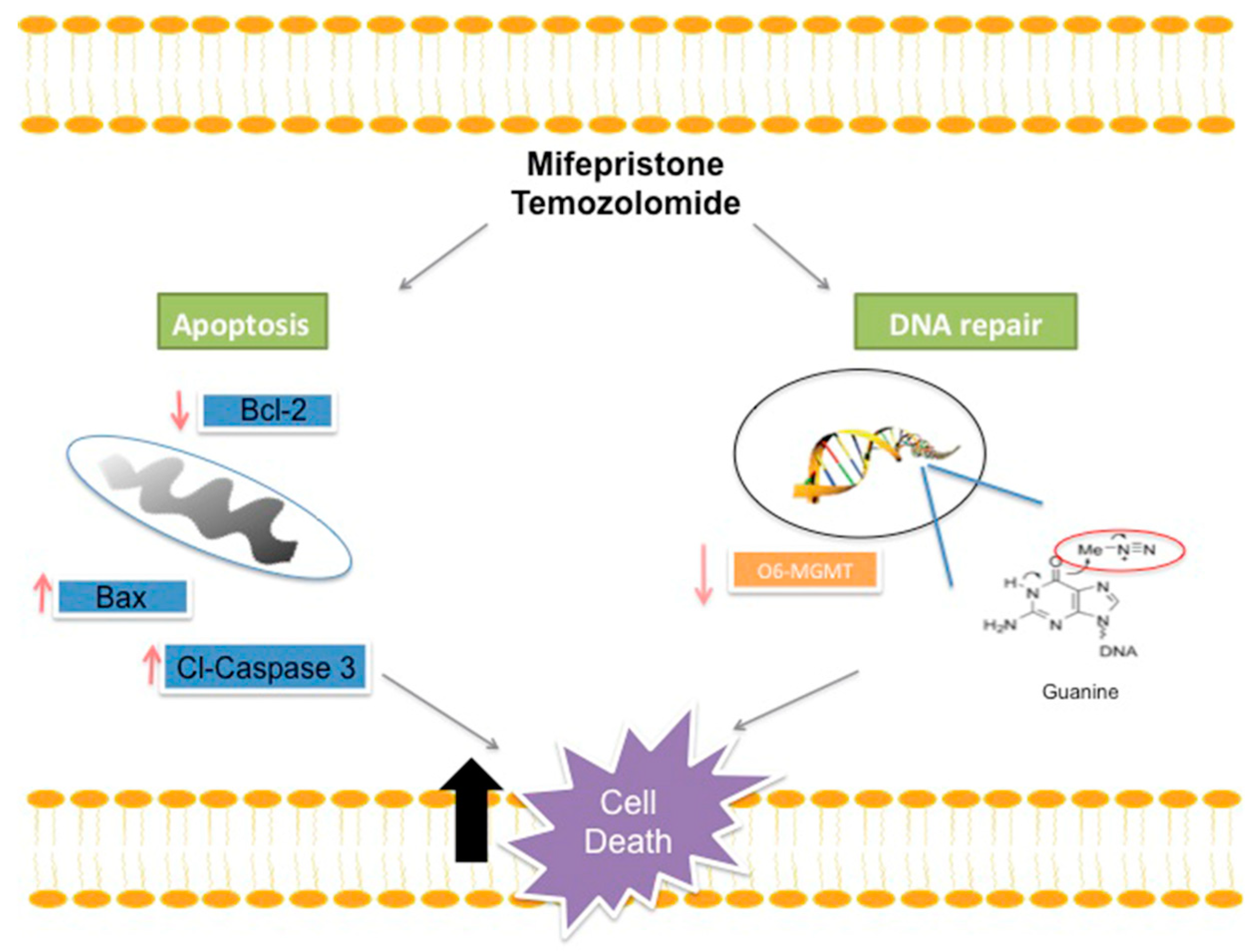
3. Discussion
4. Materials and Methods
4.1. Drugs and Reagents
4.2. Animals
4.3. Cell Culture
4.4. Tumor Cell Implantation
4.5. Treatments
4.6. Tumor Growth Assessed by Molecular Imaging
4.7. Determination of Body Weight and Overall Survival
4.8. Histology and Immunohistochemistry
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johnson, D.R.; O’Neill, B.P. Glioblastoma survival in the United States before and during the temozolomide era. J. Neurooncol. 2012, 107, 359–364. [Google Scholar] [CrossRef]
- Cardoso, F.L.; Brites, D.; Brito, M.A. Looking at the blood-brain barrier: Molecular anatomy and possible investigation approaches. Brain Res. Rev. 2010, 64, 328–363. [Google Scholar] [CrossRef]
- van Tellingen, O.; Yetkin-Arik, B.; de Gooijer, M.C.; Wesseling, P.; Wurdinger, T.; de Vries, H.E. Overcoming the blood-brain tumor barrier for effective glioblastoma treatment. Drug Resist. Updat. 2015, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, Y.P.; Weatherbee, J.L.; Wheelhouse, R.T.; Ross, A.H. Glioblastoma multiforme therapy and mechanisms of resistance. Pharmaceuticals 2013, 6, 1475–1506. [Google Scholar] [CrossRef]
- Denny, B.J.; Wheelhouse, R.T.; Stevens, M.F.; Tsang, L.L.; Slack, J.A. NMR and molecular modeling investigation of the mechanism of activation of the antitumor drug temozolomide and its interaction with DNA. Biochemistry 1994, 33, 9045–9051. [Google Scholar] [CrossRef] [PubMed]
- Kitange, G.J.; Carlson, B.L.; Mladek, A.C.; Decker, P.A.; Schroeder, M.A.; Wu, W.; Grogan, P.T.; Giannini, C.; Ballman, K.V.; Buckner, J.C.; et al. Evaluation of MGMT promoter methylation status and correlation with temozolomide response in orthotopic glioblastoma xenograft model. J. Neurooncol. 2009, 92, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.; Stupp, R.; Reifenberger, G.; Brandes, A.A.; van den Bent, M.J.; Wick, W.; Hegi, M.E. MGMT promoter methylation in malignant gliomas: Ready for personalized medicine? Nat. Rev. Neurol. 2010, 6, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Lefranc, F.; Rynkowski, M.; DeWitte, O.; Kiss, R. Present and potential future adjuvant issues in high-grade astrocytic glioma treatment. Adv. Tech. Stand Neurosurg. 2009, 34, 3–35. [Google Scholar]
- Belhadj, Z.; Zhan, C.; Ying, M.; Wei, X.; Xie, C.; Yan, Z.; Lu, W. Multifunctional targeted liposomal drug delivery for efficient glioblastoma treatment. Oncotarget 2017, 8, 66889–66900. [Google Scholar] [CrossRef] [PubMed]
- Darmon, I.; Morisse, M.C.; Coutte, A.; Blonski, M.; Le Rhun, E.; Taillandier, L.; Roufai, D.B.; Desenclos, C.; Trudel, S.; Faivre, J.C.; et al. Temozolomide and Bevacizumab Induction before Chemoradiotherapy in Patients with Bulky Glioblastoma and/or with Severe Neurological Impairment. J. Cancer 2017, 8, 1417–1424. [Google Scholar] [CrossRef]
- Toms, S.A.; Tapinos, N. Recent Advances in the Treatment of Gliomas—Comprehensive Brain Tumor Center. R. I. Med. J. 2017, 100, 43–46. [Google Scholar]
- Gonzalez-Aguero, G.; Gutierrez, A.A.; Gonzalez-Espinosa, D.; Solano, J.D.; Morales, R.; Gonzalez-Arenas, A.; Cabrera-Munoz, E.; Camacho-Arroyo, I. Progesterone effects on cell growth of U373 and D54 human astrocytoma cell lines. Endocrine 2007, 32, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Pina-Medina, A.G.; Hansberg-Pastor, V.; Gonzalez-Arenas, A.; Cerbon, M.; Camacho-Arroyo, I. Progesterone promotes cell migration, invasion and cofilin activation in human astrocytoma cells. Steroids 2016, 105, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Segovia-Mendoza, M.; Jurado, R.; Mir, R.; Medina, L.A.; Prado-Garcia, H.; Garcia-Lopez, P. Antihormonal agents as a strategy to improve the effect of chemo-radiation in cervical cancer: In vitro and in vivo study. BMC Cancer 2015, 15, 21. [Google Scholar] [CrossRef] [PubMed]
- Llaguno-Munive, M.; Medina, L.A.; Jurado, R.; Romero-Pina, M.; Garcia-Lopez, P. Mifepristone improves chemo-radiation response in glioblastoma xenografts. Cancer Cell Int. 2013, 13, e29. [Google Scholar] [CrossRef] [PubMed]
- Khawli, L.A.; Prabhu, S. Drug delivery across the blood-brain barrier. Mol. Pharm. 2013, 10, 1471–1472. [Google Scholar] [CrossRef] [PubMed]
- Im, A.; Appleman, L.J. Mifepristone: Pharmacology and clinical impact in reproductive medicine, endocrinology and oncology. Expert Opin. Pharmacother. 2010, 11, 481–488. [Google Scholar] [CrossRef]
- Spitz, I.M.; Bardin, C.W. Mifepristone (RU 486)—A modulator of progestin and glucocorticoid action. N. Engl. J. Med. 1993, 329, 404–412. [Google Scholar]
- Schwarzenberg, J.; Czernin, J.; Cloughesy, T.F.; Ellingson, B.M.; Pope, W.B.; Geist, C.; Dahlbom, M.; Silverman, D.H.; Satyamurthy, N.; Phelps, M.E.; et al. 3′-deoxy-3′-18F-fluorothymidine PET and MRI for early survival predictions in patients with recurrent malignant glioma treated with bevacizumab. J. Nucl. Med. 2012, 53, 29–36. [Google Scholar] [CrossRef]
- Moonshi, S.S.; Bejot, R.; Atcha, Z.; Vijayaragavan, V.; Bhakoo, K.K.; Goggi, J.L. A comparison of PET imaging agents for the assessment of therapy efficacy in a rodent model of glioma. Am. J. Nucl. Med. Mol. Imaging 2013, 3, 397–407. [Google Scholar]
- Tieszen, C.R.; Goyeneche, A.A.; Brandhagen, B.N.; Ortbahn, C.T.; Telleria, C.M. Antiprogestin mifepristone inhibits the growth of cancer cells of reproductive and non-reproductive origin regardless of progesterone receptor expression. BMC Cancer 2011, 11, e207. [Google Scholar] [CrossRef] [PubMed]
- Goyeneche, A.A.; Caron, R.W.; Telleria, C.M. Mifepristone inhibits ovarian cancer cell growth in vitro and in vivo. Clin. Cancer Res. 2007, 13, 3370–3379. [Google Scholar] [CrossRef] [PubMed]
- German-Castelan, L.; Manjarrez-Marmolejo, J.; Gonzalez-Arenas, A.; Gonzalez-Moran, M.G.; Camacho-Arroyo, I. Progesterone induces the growth and infiltration of human astrocytoma cells implanted in the cerebral cortex of the rat. Biomed. Res. Int. 2014, 2014, e393174. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Munoz, E.; Gonzalez-Arenas, A.; Saqui-Salces, M.; Camacho, J.; Larrea, F.; Garcia-Becerra, R.; Camacho-Arroyo, I. Regulation of progesterone receptor isoforms content in human astrocytoma cell lines. J. Steroid Biochem. Mol. Biol. 2009, 113, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Tavares, C.B.; Gomes-Braga, F.d.C.S.A.; Costa-Silva, D.R.; Escórcio-Dourado, C.S.; Borges, U.S.; Conde, A.M., Jr.; Barros-Oliveira, M.d.C.; Sousa, E.B.; Barros, L.d.R.; Martins, L.M.; et al. Expression of estrogen and progesterone receptors in astrocytomas: A literature review. Clinics 2016, 71, 481–486. [Google Scholar] [CrossRef]
- Valadez-Cosmes, P.; German-Castelan, L.; Gonzalez-Arenas, A.; Velasco-Velazquez, M.A.; Hansberg-Pastor, V.; Camacho-Arroyo, I. Expression and hormonal regulation of membrane progesterone receptors in human astrocytoma cells. J. Steroid Biochem. Mol. Biol. 2015, 154, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Cunningham, R.L.; Rybalchenko, N.; Singh, M. Progesterone increases the release of brain-derived neurotrophic factor from glia via progesterone receptor membrane component 1 (Pgrmc1)-dependent ERK5 signaling. Endocrinology 2012, 153, 4389–4400. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Q.; Wang, Z.B.; Bai, J.; Zhao, J.; Wang, Y.; Hu, K.; Du, Y.H. Effects of mifepristone on invasive and metastatic potential of human gastric adenocarcinoma cell line MKN-45 in vitro and in vivo. World J. Gastroenterol. 2004, 10, 1726–1729. [Google Scholar] [CrossRef]
- Li, A.; Felix, J.C.; Minoo, P.; Amezcua, C.A.; Jain, J.K. Effect of mifepristone on proliferation and apoptosis of Ishikawa endometrial adenocarcinoma cells. Fertil. Steril. 2005, 84, 202–211. [Google Scholar] [CrossRef]
- Hegi, M.E.; Diserens, A.C.; Gorlia, T.; Hamou, M.F.; de Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L.; et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003. [Google Scholar] [CrossRef]
- Binabaj, M.M.; Bahrami, A.; ShahidSales, S.; Joodi, M.; Joudi Mashhad, M.; Hassanian, S.M.; Anvari, K.; Avan, A. The prognostic value of MGMT promoter methylation in glioblastoma: A meta-analysis of clinical trials. J. Cell. Physiol. 2018, 233, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Paz, M.F.; Yaya-Tur, R.; Rojas-Marcos, I.; Reynes, G.; Pollan, M.; Aguirre-Cruz, L.; Garcia-Lopez, J.L.; Piquer, J.; Safont, M.J.; Balana, C.; et al. CpG island hypermethylation of the DNA repair enzyme methyltransferase predicts response to temozolomide in primary gliomas. Clin. Cancer Res. 2004, 10, 4933–4938. [Google Scholar] [CrossRef] [PubMed]
- Cabrini, G.; Fabbri, E.; Lo Nigro, C.; Dechecchi, M.C.; Gambari, R. Regulation of expression of O6-methylguanine-DNA methyltransferase and the treatment of glioblastoma (Review). Int. J. Oncol. 2015, 47, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Aasland, D.; Reich, T.R.; Tomicic, M.T.; Switzeny, O.J.; Kaina, B.; Christmann, M. Repair gene O (6)-methylguanine-DNA methyltransferase is controlled by SP1 and up-regulated by glucocorticoids, but not by temozolomide and radiation. J. Neurochem. 2018, 144, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Navo, M.A.; Smith, J.A.; Gaikwad, A.; Burke, T.; Brown, J.; Ramondetta, L.M. In vitro evaluation of the growth inhibition and apoptosis effect of mifepristone (RU486) in human Ishikawa and HEC1A endometrial cancer cell lines. Cancer Chemother Pharmacol. 2008, 62, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ye, H. Mifepristone sensitizing cisplatin for cervical adenocarcinoma HeLa cell sensitivity to chemotherapy and its mechanism. Eur. J. Gynaecol. Oncol. 2013, 34, 142–147. [Google Scholar]
- Biswas, T.; Ramana, C.V.; Srinivasan, G.; Boldogh, I.; Hazra, T.K.; Chen, Z.; Tano, K.; Thompson, E.B.; Mitra, S. Activation of human O6-methylguanine-DNA methyltransferase gene by glucocorticoid hormone. Oncogene 1999, 18, 525–532. [Google Scholar] [CrossRef]
- Ueda, S.; Mineta, T.; Nakahara, Y.; Okamoto, H.; Shiraishi, T.; Tabuchi, K. Induction of the DNA repair gene O6-methylguanine-DNA methyltransferase by dexamethasone in glioblastomas. J. Neurosurg. 2004, 101, 659–663. [Google Scholar] [CrossRef]
- Horiguchi, M.; Kim, J.; Matsunaga, N.; Kaji, H.; Egawa, T.; Makino, K.; Koyanagi, S.; Ohdo, S. Glucocorticoid-dependent expression of O (6)-methylguanine-DNA methyltransferase gene modulates dacarbazine-induced hepatotoxicity in mice. J. Pharmacol. Exp. Ther. 2010, 333, 782–787. [Google Scholar] [CrossRef]
- Check, J.H.; Wilson, C.; Cohen, R.; Sarumi, M. Evidence that Mifepristone, a progesterone receptor antagonist, can cross the blood brain barrier and provide palliative benefits for glioblastoma multiforme grade IV. Anticancer Res. 2014, 34, 2385–2388. [Google Scholar] [CrossRef]
- Check, J.H.; Check, D.; Cohen, R.; Sarumi, M. Mifepristone causing complete remission of rapidly advancing leukemia with measurement of progesterone-induced blocking factor. Anticancer Res. 2014, 34, 2413–2416. [Google Scholar] [PubMed]
- Grunberg, S.M.; Weiss, M.H.; Russell, C.A.; Spitz, I.M.; Ahmadi, J.; Sadun, A.; Sitruk-Ware, R. Long-term administration of mifepristone (RU486): Clinical tolerance during extended treatment of meningioma. Cancer Investig. 2006, 24, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 4th ed.; Academic Press: New York, NY, USA, 1998. [Google Scholar]
- Lee, S.Y. Temozolomide resistance in glioblastoma multiforme. Genes Dis. 2016, 3, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Nagane, M. Dose-dense temozolomide: Is it still promising? Neurol. Med. Chir. 2015, 55, 38–49. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Llaguno-Munive, M.; Romero-Piña, M.; Serrano-Bello, J.; Medina, L.A.; Uribe-Uribe, N.; Salazar, A.M.; Rodríguez-Dorantes, M.; Garcia-Lopez, P. Mifepristone Overcomes Tumor Resistance to Temozolomide Associated with DNA Damage Repair and Apoptosis in an Orthotopic Model of Glioblastoma. Cancers 2019, 11, 16. https://doi.org/10.3390/cancers11010016
Llaguno-Munive M, Romero-Piña M, Serrano-Bello J, Medina LA, Uribe-Uribe N, Salazar AM, Rodríguez-Dorantes M, Garcia-Lopez P. Mifepristone Overcomes Tumor Resistance to Temozolomide Associated with DNA Damage Repair and Apoptosis in an Orthotopic Model of Glioblastoma. Cancers. 2019; 11(1):16. https://doi.org/10.3390/cancers11010016
Chicago/Turabian StyleLlaguno-Munive, Monserrat, Mario Romero-Piña, Janeth Serrano-Bello, Luis A. Medina, Norma Uribe-Uribe, Ana Maria Salazar, Mauricio Rodríguez-Dorantes, and Patricia Garcia-Lopez. 2019. "Mifepristone Overcomes Tumor Resistance to Temozolomide Associated with DNA Damage Repair and Apoptosis in an Orthotopic Model of Glioblastoma" Cancers 11, no. 1: 16. https://doi.org/10.3390/cancers11010016
APA StyleLlaguno-Munive, M., Romero-Piña, M., Serrano-Bello, J., Medina, L. A., Uribe-Uribe, N., Salazar, A. M., Rodríguez-Dorantes, M., & Garcia-Lopez, P. (2019). Mifepristone Overcomes Tumor Resistance to Temozolomide Associated with DNA Damage Repair and Apoptosis in an Orthotopic Model of Glioblastoma. Cancers, 11(1), 16. https://doi.org/10.3390/cancers11010016