Microfluidic-Based 3D Engineered Microvascular Networks and Their Applications in Vascularized Microtumor Models


Abstract
1. Introduction
2. In Vitro Vascularization Strategies
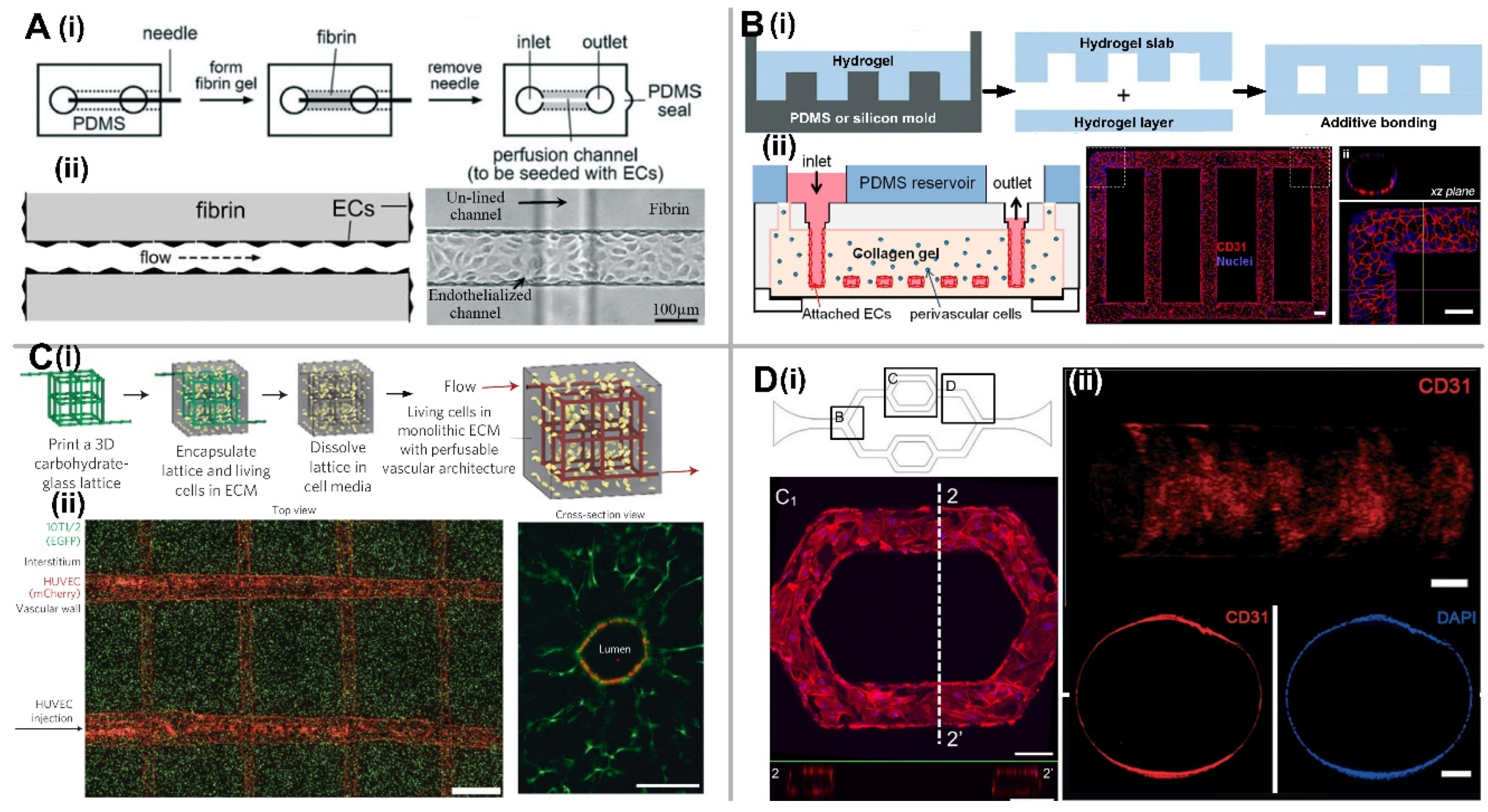
2.1. EC Lining-Based Methods
2.1.1. Microneedle-Based Removable Method
2.1.2. Micropatterned Planar Hydrogel Slab Bonding Method
2.1.3. Dissolvable Materials-Based Sacrificial Micromolding Method
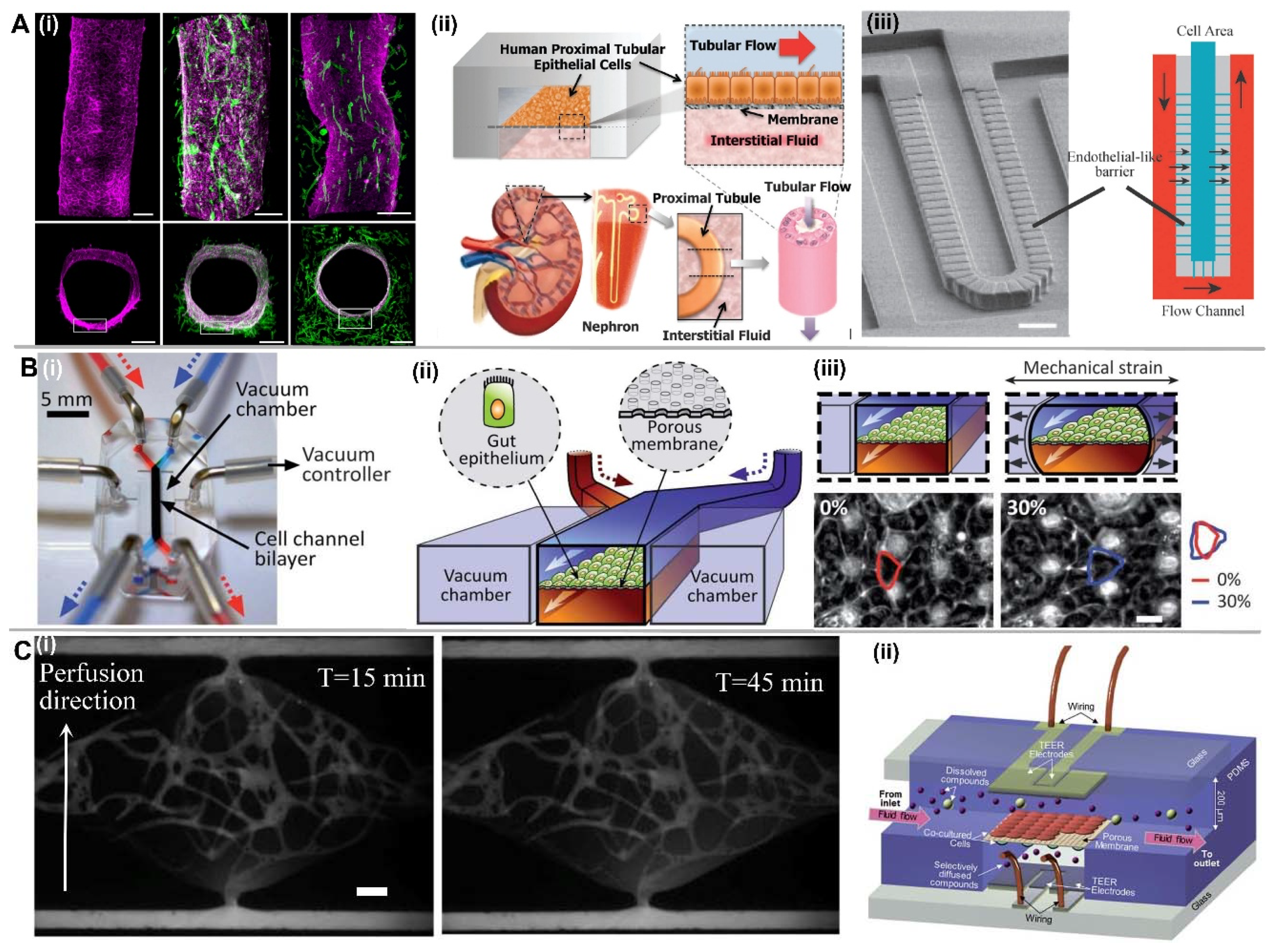
2.1.4. EC Lining inside a PDMS-Based Microfluidic Channel
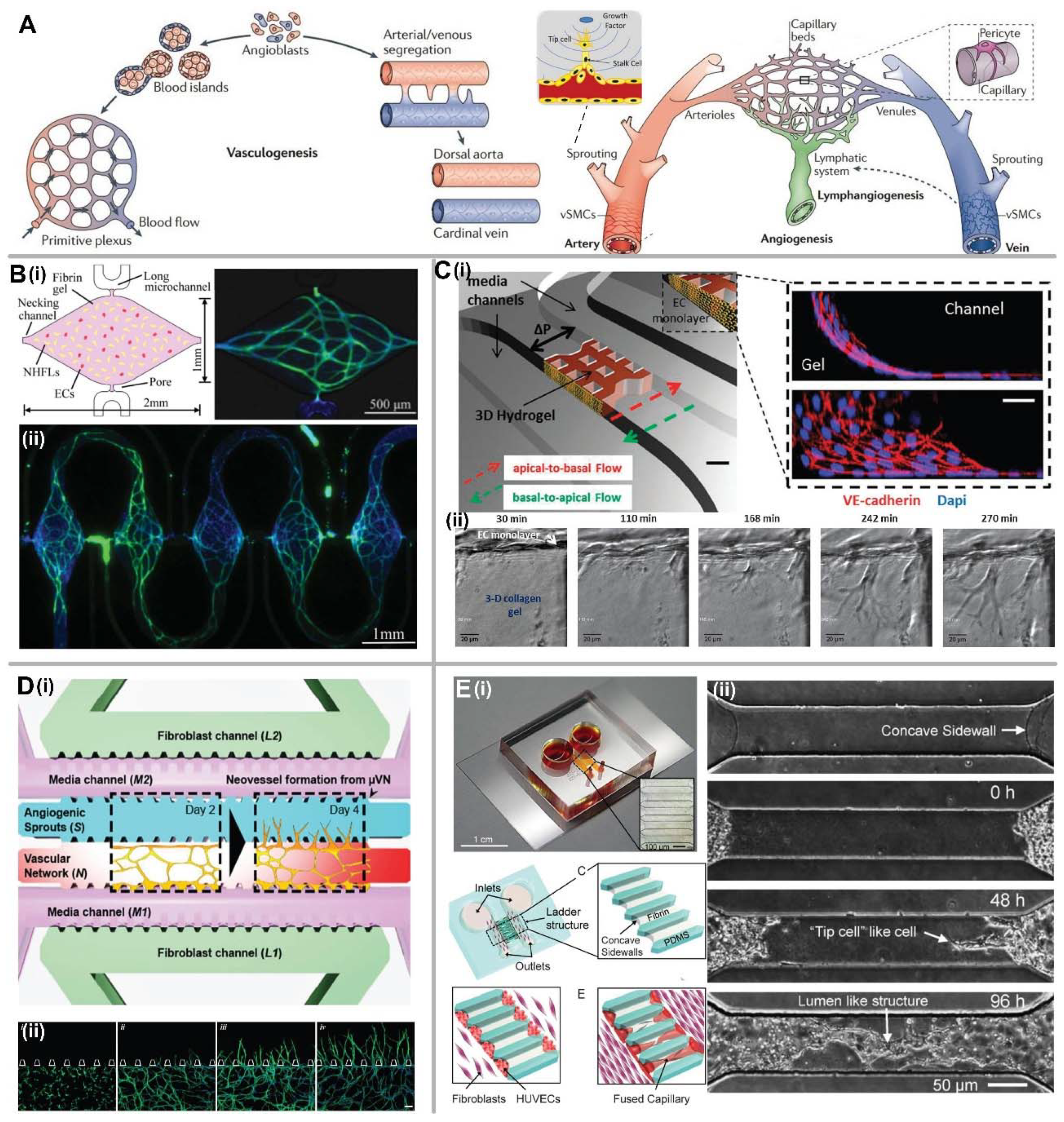
2.2. Vasculogenesis and Angiogenesis-Based Methods
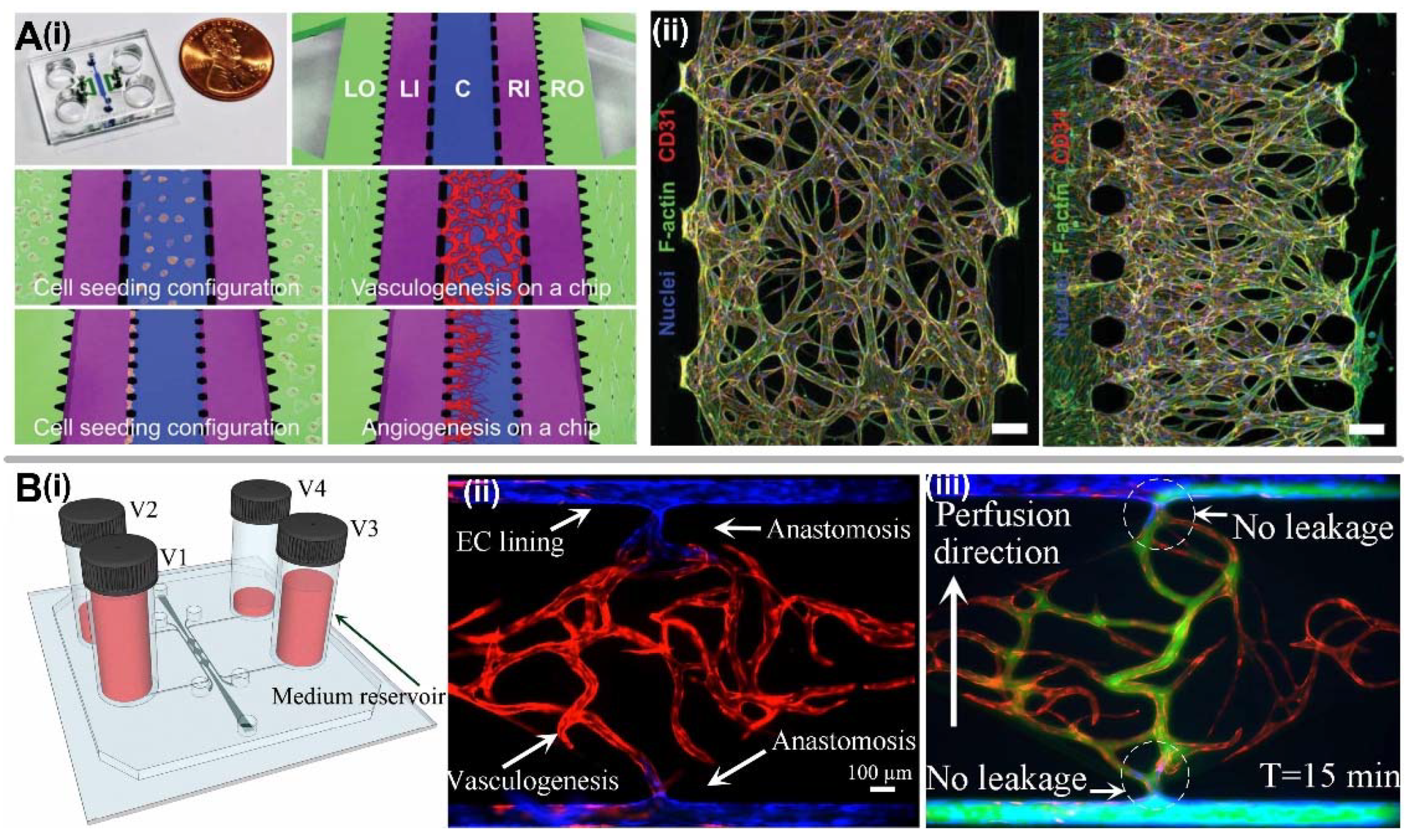
2.2.1. Vasculogenesis
2.2.2. Angiogenesis
2.3. Hybrid Methods
3. Vascular Inducing Factors
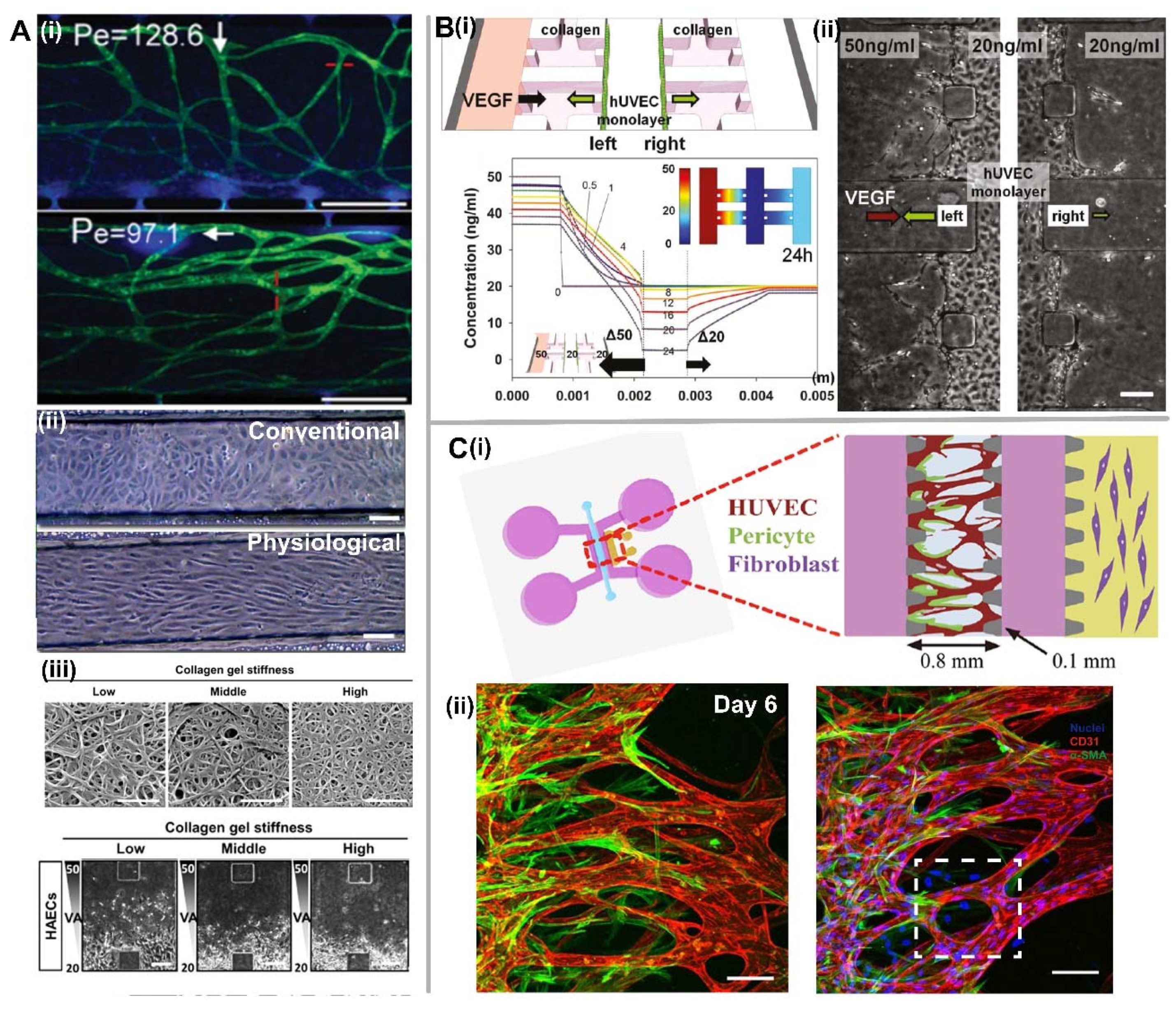
3.1. Biomechanical Factors
3.2. Extracellular (or Diffusible) Signaling Molecules
3.3. Cell Source and Cell-Cell Interaction
4. Selective Vascular Barrier
5. Application of Engineered Microvascular Networks to Cancer Biology
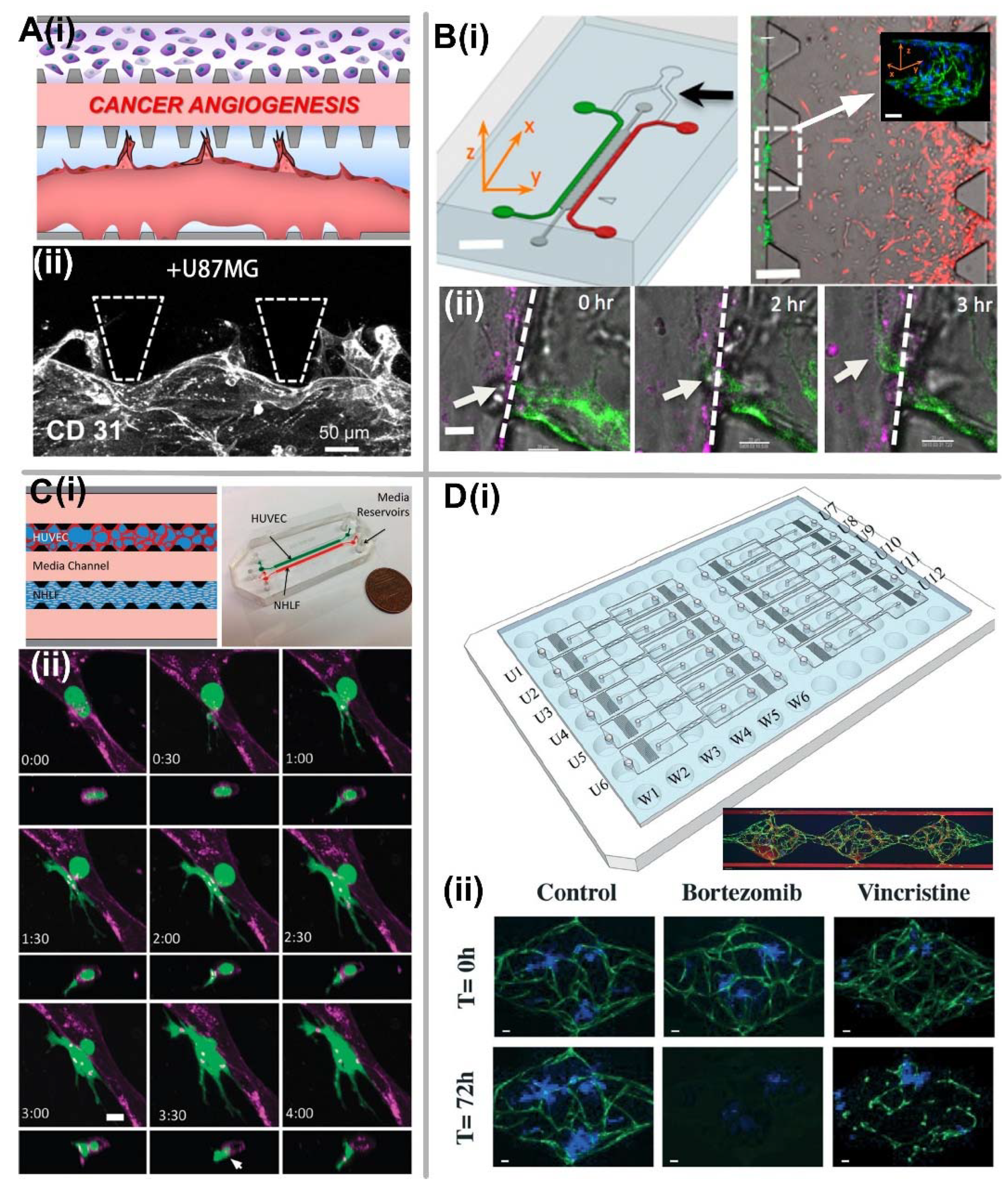
5.1. Tumor Angiogenesis
5.2. Tumor Intravasation
5.3. Tumor Extravasation
5.4. Tumor Microenvironment
5.5. Application of Vascularized Tumor-on-a-Chip
6. Conclusions and Future Perspectives
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D | Three-dimensional |
| 2D | Two-dimensional |
| ECs | Endothelial cells |
| PDMS | Polydimethylsiloxane |
| HUVECs | Human umbilical vein endothelial cells |
| PEG | Polyethylene glycol |
| SMCs | Smooth muscle cells |
| ECM | Extracellular matrix |
| EPCs | Endothelial progenitor cells |
| ECFC-ECs | Endothelial colony forming cell-derived endothelial cells |
| NHLF | Normal human lung fibroblast |
| VEGF | Vascular endothelial growth factor |
| bFGF | Basic fibroblast growth factor |
| MMPs | Metalloproteinases |
| DAPI | 4′,6-diamidino-2-phenylindole |
| FITC | Fluorescein isothiocyanate |
| NO | Nitric oxide |
| Ang-1 | Angiopoietin 1 |
| TGF-β1 | Transforming growth factor beta 1 |
| HIF | Hypoxia inducible factor |
| iPSCs | Induced pluripotent stem cells |
| MSCs | Mesenchymal stem cells |
| hESCs | Human embryonic stem cells |
| BBB | Blood-brain barrier |
| PECAM-1 | Platelet endothelial cell adhesion molecule-1 |
| VE | Vascular endothelial |
| BSA | Bovine serum albumin |
| TEER | Transendothelial electrical resistance |
| TNF-α | Tumor necrosis factor alpha |
| CTCs | Circulating tumor cells |
| TME | Tumor microenvironment |
References
- Jain, R. Molecular regulation of vessel maturation. Nat. Med. 2003, 9, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Ingber, D. Microfluidic organs-on-chips. Nat. Biotechnol. 2014, 32, 760–772. [Google Scholar] [CrossRef] [PubMed]
- Rayner, S.; Zheng, Y. Engineered microvessels for the study of human disease. J. Biomech. Eng. 2016, 138, 110801. [Google Scholar] [CrossRef] [PubMed]
- Kramer, R.; Nicolson, G. Interactions of tumor cells with vascular endothelial cell monolayers: A model for metastatic invasion. Proc. Nat. Acad. Sci. USA 1979, 76, 5704–5708. [Google Scholar] [CrossRef] [PubMed]
- Young, E.; Watson, M.; Srigunapalan, S.; Wheeler, A.; Simmons, C. Technique for real-time measurements of endothelial permeability in a microfluidic membrane chip using laser-induced fluorescence detection. Anal. Chem. 2010, 82, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, Z.; Wdzieczak-Bakala, J.; Pang, D.; Liu, J.; Chen, Y. Patterning cells and shear flow conditions: Convenient observation of endothelial cell remoulding, enhanced production of angiogenesis factors and drug response. Lab Chip 2011, 11, 4235–4240. [Google Scholar] [CrossRef] [PubMed]
- Pelton, J.C.; Wright, C.E.; Leitges, M.; Bautch, V.L. Multiple endothelial cells constitute the tip of developing blood vessels and polarize to promote lumen formation. Development 2014, 141, 4121–4126. [Google Scholar] [CrossRef] [PubMed]
- Halldorsson, S.; Lucumi, E.; Gómez-Sjöberg, R.; Fleming, R. Advantages and challenges of microfluidic cell culture in polydimethylsiloxane devices. Biosens. Bioelectron. 2015, 63, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.; Paul, A.; Vrana, N.E.; Zhao, X.; Memic, A.; Hwang, Y.S.; Dokmeci, M.R.; Khademhosseini, A. Microfluidic techniques for development of 3D vascularized tissue. Biomaterials 2014, 35, 7308–7325. [Google Scholar] [CrossRef] [PubMed]
- Tien, J.; Wong, K.H.K.; Truslow, J.G. Vascularization of microfluidic hydrogels. In Microfluidic Cell Culture Systems; Bettinger, C., Borenstein, J.T., Tao, S.L., Eds.; Elsevier: Oxford, UK, 2013; pp. 205–221. [Google Scholar]
- Chrobak, K.; Potter, D.; Tien, J. Formation of perfused, functional microvascular tubes in vitro. Microvasc. Res. 2006, 71, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Price, G.; Wong, K.; Truslow, J.; Leung, A.; Acharya, C.; Tien, J. Effect of mechanical factors on the function of engineered human blood microvessels in microfluidic collagen gels. Biomaterials 2010, 31, 6182–6189. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.; Truslow, J.; Khankhel, A.; Chan, K.; Tien, J. Artificial lymphatic drainage systems for vascularized microfluidic scaffolds. J. Biomed. Mater. Res. Part A 2012, 101A, 2181–2190. [Google Scholar] [CrossRef] [PubMed]
- Sadr, N.; Zhu, M.; Osaki, T.; Kakegawa, T.; Yang, Y.; Moretti, M.; Fukuda, J.; Khademhosseini, A. SAM-based cell transfer to photopatterned hydrogels for microengineering vascular-like structures. Biomaterials 2011, 32, 7479–7490. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Matsusaki, M.; Akashi, M. Multilayered blood capillary analogs in biodegradable hydrogels for in vitro drug permeability assays. Adv. Funct. Mater. 2012, 23, 1736–1742. [Google Scholar] [CrossRef]
- Buchanan, C.; Voigt, E.; Szot, C.; Freeman, J.; Vlachos, P.; Rylander, M. Three-dimensional microfluidic collagen hydrogels for investigating flow-mediated tumor-endothelial signaling and vascular organization. Tissue Eng. Part C 2014, 20, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Chen, J.; Craven, M.; Choi, N.; Totorica, S.; Diaz-Santana, A.; Kermani, P.; Hempstead, B.; Fischbach-Teschl, C.; Lopez, J.; et al. In vitro microvessels for the study of angiogenesis and thrombosis. Proc. Nat. Acad. Sci. USA 2012, 109, 9342–9347. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.; Stevens, K.; Yang, M.; Baker, B.; Nguyen, D.; Cohen, D.; Toro, E.; Chen, A.; Galie, P.; Yu, X.; et al. Rapid casting of patterned vascular networks for perfusable engineered three-dimensional tissues. Nat. Mater. 2012, 11, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Kolesky, D.; Truby, R.; Gladman, A.; Busbee, T.; Homan, K.; Lewis, J. 3D bioprinting of vascularized, heterogeneous cell-laden tissue constructs. Adv. Mater. 2014, 26, 3124–3130. [Google Scholar] [CrossRef] [PubMed]
- Bertassoni, L.; Cecconi, M.; Manoharan, V.; Nikkhah, M.; Hjortnaes, J.; Cristino, A.; Barabaschi, G.; Demarchi, D.; Dokmeci, M.; Yang, Y.; et al. Hydrogel bioprinted microchannel networks for vascularization of tissue engineering constructs. Lab Chip 2014, 14, 2202–2211. [Google Scholar] [CrossRef] [PubMed]
- Golden, A.; Tien, J. Fabrication of microfluidic hydrogels using molded gelatin as a sacrificial element. Lab Chip 2007, 7, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jin, Z.; Gan, B.; Lv, S.; Xie, M.; Huang, W. Engineering interconnected 3D vascular networks in hydrogels using molded sodium alginate lattice as the sacrificial template. Lab Chip 2014, 14, 2709–2716. [Google Scholar] [CrossRef] [PubMed]
- Tsang, V.; Chen, A.; Cho, L.; Jadin, K.; Sah, R.; DeLong, S.; West, J.; Bhatia, S. Fabrication of 3D hepatic tissues by additive photopatterning of cellular hydrogels. FASEB J. 2007, 21, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Whitesides, G. Soft lithography. Annu. Rev. Mater. Sci. 1998, 28, 153–184. [Google Scholar] [CrossRef]
- McGuigan, A.; Sefton, M. The influence of biomaterials on endothelial cell thrombogenicity. Biomaterials 2007, 28, 2547–2571. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Phan, D.; Sobrino, A.; George, S.; Hughes, C.; Lee, A. Engineering anastomosis between living capillary networks and endothelial cell-lined microfluidic channels. Lab Chip 2016, 16, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, S.; He, P.; Liu, Y. In vitro recapitulation of functional microvessels for the study of endothelial shear response, nitric oxide and (Ca2+)i. PLoS ONE 2015, 10, e0126797. [Google Scholar] [CrossRef] [PubMed]
- Esch, M.; Post, D.; Shuler, M.; Stokol, T. Characterization of in vitro endothelial linings grown within microfluidic channels. Tissue Eng. Part A 2011, 17, 2965–2971. [Google Scholar] [CrossRef] [PubMed]
- Bischel, L.; Young, E.; Mader, B.; Beebe, D. Tubeless microfluidic angiogenesis assay with three-dimensional endothelial-lined microvessels. Biomaterials 2013, 34, 1471–1477. [Google Scholar] [CrossRef] [PubMed]
- Bischel, L.; Lee, S.; Beebe, D. A practical method for patterning lumens through ECM hydrogels via viscous fingering patterning. J. Lab. Autom. 2012, 17, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Abdelgawad, M.; Wu, C.; Chien, W.; Geddie, W.; Jewett, M.; Sun, Y. A fast and simple method to fabricate circular microchannels in polydimethylsiloxane (PDMS). Lab Chip 2011, 11, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Fiddes, L.; Raz, N.; Srigunapalan, S.; Tumarkan, E.; Simmons, C.; Wheeler, A.; Kumacheva, E. A circular cross-section PDMS microfluidics system for replication of cardiovascular flow conditions. Biomaterials 2010, 31, 3459–3464. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, Y.; Bakht, S.; Aleman, J.; Shin, S.; Yue, K.; Sica, M.; Ribas, J.; Duchamp, M.; Ju, J.; et al. Elastomeric free-form blood vessels for interconnecting organs on chip systems. Lab Chip 2016, 16, 1579–1586. [Google Scholar] [CrossRef] [PubMed]
- Patan, S. Vasculogenesis and angiogenesis. Cancer Treat. Res. 2004, 117, 3–32. [Google Scholar] [CrossRef] [PubMed]
- Potente, M.; Makinen, T. Vascular heterogeneity and specialization in development and disease. Nat. Rev. Mol. Cell Biol. 2017, 16, 477–494. [Google Scholar] [CrossRef] [PubMed]
- Herbert, S.P.; Stainier, D.Y. Molecular control of endothelial cell behaviour during blood vessel morphogenesis. Nat. Rev. Mol. Cell Biol. 2011, 12, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Tepper, O.M.; Capla, J.M.; Galiano, R.D.; Ceradini, D.J.; Callaghan, M.J.; Kleinman, M.E.; Gurtner, G.C. Adult vasculogenesis occurs through in situ recruitment, proliferation, and tubulization of circulating bone marrow-derived cells. Blood 2005, 105, 1068–1077. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, P.; Keane, M.P. Physiological and pathological angiogenesis in the adult pulmonary circulation. Compr. Physiol. 2011, 1, 1473–1508. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.; Moya, M.; Hughes, C.; George, S.; Lee, A. A microfluidic platform for generating large-scale nearly identical human microphysiological vascularized tissue arrays. Lab Chip 2013, 13, 2990–2998. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.; Moya, M.; Abiri, P.; Hughes, C.; George, S.; Lee, A. Full range physiological mass transport control in 3D tissue cultures. Lab Chip 2013, 13, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Moya, M.; Hsu, Y.; Lee, A.; Hughes, C.; George, S. In vitro perfused human capillary networks. Tissue Eng. Part C 2013, 19, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Newman, A.C.; Nakatsu, M.N.; Chou, W.; Gershon, P.D.; Hughes, C.C. The requirement for fibroblasts in angiogenesis: Fibroblast-derived matrix proteins are essential for endothelial cell lumen formation. Mol. Biol. Cell 2011, 22, 3791–3800. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, S.; Nelson, C.; Baranski, J.; Lim, E.; Chen, C. Geometrically controlled endothelial tubulogenesis in micropatterned gels. Tissue Eng. Part A 2010, 16, 2255–2263. [Google Scholar] [CrossRef] [PubMed]
- Ghajar, C.M.; Chen, X.; Harris, J.W.; Suresh, V.; Hughes, C.C.; Jeon, N.L.; Putnam, A.J.; George, S.C. The effect of matrix density on the regulation of 3-D capillary morphogenesis. Biophys. J. 2008, 94, 1930–1941. [Google Scholar] [CrossRef] [PubMed]
- Whisler, J.; Chen, M.; Kamm, R. Control of perfusable microvascular network morphology using a multiculture microfluidic system. Tissue Eng. Part C 2014, 20, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Mendes, L.F.; Pirraco, R.P.; Szymczyk, W.; Frias, A.M.; Santos, T.C.; Reis, R.L.; Marques, A.P. Perivascular-like cells contribute to the stability of the vascular network of osteogenic tissue formed from cell sheet-based constructs. PLoS ONE 2012, 7, e41051. [Google Scholar] [CrossRef] [PubMed]
- Logsdon, E.; Finley, S.; Popel, A.; Gabhann, F. A systems biology view of blood vessel growth and remodelling. J. Cell. Mol. Med. 2013, 18, 1491–1508. [Google Scholar] [CrossRef] [PubMed]
- Vickerman, V.; Kamm, R. Mechanism of a flow-gated angiogenesis switch: Early signaling events at cell–matrix and cell–cell junctions. Integr. Biol. 2012, 4, 863. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.; Jeon, J.; Han, S.; Jung, G.; Shin, S.; Lee, S.; Sudo, R.; Kamm, R.; Chung, S. In vitro 3D collective sprouting angiogenesis under orchestrated ANG-1 and VEGF gradients. Lab Chip 2011, 11, 2175–2181. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.; Han, S.; Shin, Y.; Kwon, G.; Kamm, R.; Lee, S.; Chung, S. Sprouting angiogenesis under a chemical gradient regulated by interactions with an endothelial monolayer in a microfluidic platform. Anal. Chem. 2011, 83, 8454–8459. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chung, M.; Ahn, J.; Lee, S.; Jeon, N. Interstitial flow regulates the angiogenic response and phenotype of endothelial cells in a 3D culture model. Lab Chip 2016, 16, 4189–4199. [Google Scholar] [CrossRef] [PubMed]
- Yeon, J.; Ryu, H.; Chung, M.; Hu, Q.; Jeon, N. In vitro formation and characterization of a perfusable three-dimensional tubular capillary network in microfluidic devices. Lab Chip 2012, 12, 2815–2822. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Bazou, D.; Munn, L. Anastomosis of endothelial sprouts forms new vessels in a tissue analogue of angiogenesis. Integr. Biol. 2012, 4, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, H.; Chung, M.; Jeon, N. Engineering of functional, perfusable 3D microvascular networks on a chip. Lab Chip 2013, 13, 1489–1500. [Google Scholar] [CrossRef] [PubMed]
- Tzima, E.; Irani-Tehrani, M.; Kiosses, W.; Dejana, E.; Schultz, D.; Engelhardt, B.; Cao, G.; DeLisser, H.; Schwartz, M. A mechanosensory complex that mediates the endothelial cell response to fluid shear stress. Nature 2005, 437, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Khan, O.; Sefton, M. Endothelial cell behaviour within a microfluidic mimic of the flow channels of a modular tissue engineered construct. Biomed. Microdevices 2010, 13, 69–87. [Google Scholar] [CrossRef] [PubMed]
- Abaci, H.; Shen, Y.; Tan, S.; Gerecht, S. Recapitulating physiological and pathological shear stress and oxygen to model vasculature in health and disease. Sci. Rep. 2014, 4, 4951. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.; Stapleton, S.; Yang, M.; Cha, S.; Choi, C.; Galie, P.; Chen, C. Biomimetic model to reconstitute angiogenic sprouting morphogenesis in vitro. Proc. Nat. Acad. Sci. USA 2013, 110, 6712–6717. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Munn, L. Fluid forces control endothelial sprouting. Proc. Nat. Acad. Sci. USA 2011, 108, 15342–15347. [Google Scholar] [CrossRef] [PubMed]
- Galie, P.; Nguyen, D.; Choi, C.; Cohen, D.; Janmey, P.; Chen, C. Fluid shear stress threshold regulates angiogenic sprouting. Proc. Nat. Acad. Sci. USA 2014, 111, 7968–7973. [Google Scholar] [CrossRef] [PubMed]
- Swartz, M.; Fleury, M. Interstitial flow and its effects in soft tissues. Annu. Rev. Biomed. Eng. 2007, 9, 229–256. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.; Helm, C.; Swartz, M. Interstitial flow differentially stimulates blood and lymphatic endothelial cell morphogenesis in vitro. Microvasc. Res. 2004, 68, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Sudo, R.; Mack, P.; Wan, C.; Vickerman, V.; Kamm, R. Cell migration into scaffolds under co-culture conditions in a microfluidic platform. Lab Chip 2009, 9, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Jeong, H.; Joo, H.; Choi, S.; Park, C.; Kim, J.; Choi, J.; Cui, L.; Hong, S.; Chung, S.; et al. Intrinsic FGF2 and FGF5 promotes angiogenesis of human aortic endothelial cells in 3D microfluidic angiogenesis system. Sci. Rep. 2016, 6, 28832. [Google Scholar] [CrossRef] [PubMed]
- Laco, F.; Grant, M.; Black, R. Collagen-nanofiber hydrogel composites promote contact guidance of human lymphatic microvascular endothelial cells and directed capillary tube formation. J. Biomed. Mater. Res. A 2012, 101A, 1787–1799. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, D.; Landau, S.; Shandalov, Y.; Raindel, N.; Freiman, A.; Shor, E.; Blinder, Y.; Vandenburgh, H.H.; Mooney, D.J.; Levenberg, S. Morphogenesis of 3D vascular networks is regulated by tensile forces. Proc. Natl. Acad. Sci. USA 2016, 113, 3215–3220. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N. VEGF-A: A critical regulator of blood vessel growth. Eur. Cytokine Netw. 2009, 20, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu, M.; Sainson, R.; Aoto, J.; Taylor, K.; Aitkenhead, M.; Pérez-del-Pulgar, S.; Carpenter, P.; Hughes, C. Angiogenic sprouting and capillary lumen formation modeled by human umbilical vein endothelial cells (HUVEC) in fibrin gels: The role of fibroblasts and Angiopoietin-1. Microvasc. Res. 2003, 66, 102–112. [Google Scholar] [CrossRef]
- Gerhardt, H.; Golding, M.; Fruttiger, M.; Ruhrberg, C.; Lundkvist, A.; Abramsson, A.; Jeltsch, M.; Mitchell, C.; Alitalo, K.; Shima, D.; et al. VEGF guides angiogenic sprouting utilizing endothelial tip cell filopodia. J. Cell Biol. 2003, 161, 1163–1177. [Google Scholar] [CrossRef] [PubMed]
- Del Amo, C.; Borau, C.; Gutiérrez, R.; Asín, J.; García-Aznar, J. Quantification of angiogenic sprouting under different growth factors in a microfluidic platform. J. Biomech. 2016, 49, 1340–1346. [Google Scholar] [CrossRef] [PubMed]
- Perets, A.; Baruch, Y.; Weisbuch, F.; Shoshany, G.; Neufeld, G.; Cohen, S. Enhancing the vascularization of three-dimensional porous alginate scaffolds by incorporating controlled release basic fibroblast growth factor microspheres. J. Biomed. Mater. Res. A 2003, 65A, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Kim, C.; Aref, A.; Kamm, R.; Raghunath, M. Complementary effects of ciclopirox olamine, a prolyl hydroxylase inhibitor and sphingosine 1-phosphate on fibroblasts and endothelial cells in driving capillary sprouting. Integr. Biol. 2013, 5, 1474–1484. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Silva, E.; Manssonbroberg, A.; Grinnemo, K.; Siddiqui, A.; Dellgren, G.; Wardell, E.; Brodin, L.; Mooney, D.; Sylven, C. Angiogenic effects of sequential release of VEGF-A165 and PDGF-BB with alginate hydrogels after myocardial infarction. Cardiovasc. Res. 2007, 75, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Nillesen, S.; Geutjes, P.; Wismans, R.; Schalkwijk, J.; Daamen, W.; van Kuppevelt, T. Increased angiogenesis and blood vessel maturation in acellular collagen–heparin scaffolds containing both FGF2 and VEGF. Biomaterials 2007, 28, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.; Trappmann, B.; Stapleton, S.; Toro, E.; Chen, C. Microfluidics embedded within extracellular matrix to define vascular architectures and pattern diffusive gradients. Lab Chip 2013, 13, 3246–3252. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.; Bersini, S.; Whisler, J.; Chen, M.; Dubini, G.; Charest, J.; Moretti, M.; Kamm, R. Generation of 3D functional microvascular networks with human mesenchymal stem cells in microfluidic systems. Integr. Biol. 2014, 6, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.; Mooney, D. Effects of VEGF temporal and spatial presentation on angiogenesis. Biomaterials 2010, 31, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Krock, B.; Skuli, N.; Simon, M. Hypoxia-induced angiogenesis good and evil. Genes Cancer 2011, 2, 1117–1133. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.; Rexius-Hall, M.; Elgass, L.; Eddington, D. Oxygen control with microfluidics. Lab Chip 2014, 14, 4305–4318. [Google Scholar] [CrossRef] [PubMed]
- Oomen, P.; Skolimowski, M.; Verpoorte, E. Implementing oxygen control in chip-based cell and tissue culture systems. Lab Chip 2016, 16, 3394–3414. [Google Scholar] [CrossRef] [PubMed]
- Mehta, G.; Lee, J.; Cha, W.; Tung, Y.; Linderman, J.; Takayama, S. Hard top soft bottom microfluidic devices for cell culture and chemical analysis. Anal. Chem. 2009, 81, 3714–3722. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Cheng, Y.; Tu, M.; Chen, Y.; Peng, C.; Liao, W.; Tung, Y. A polydimethylsiloxane–polycarbonate hybrid microfluidic device capable of generating perpendicular chemical and oxygen gradients for cell culture studies. Lab Chip 2014, 14, 3762–3772. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Yang, K.; Park, H.; Cho, S.; Han, S.; Shin, Y.; Chung, S.; Lee, J. Implantable microfluidic device for the formation of three-dimensional vasculature by human endothelial progenitor cells. Biotechnol. Bioprocess Eng. 2014, 19, 379–385. [Google Scholar] [CrossRef]
- Dai, X.; Cai, S.; Ye, Q.; Jiang, J.; Yan, X.; Xiong, X.; Jiang, Q.; Wang, A.; Tan, Y. A novel in vitro angiogenesis model based on a microfluidic device. Chin. Sci. Bull. 2011, 56, 3301–3309. [Google Scholar] [CrossRef] [PubMed]
- Samuel, R.; Daheron, L.; Liao, S.; Vardam, T.; Kamoun, W.; Batista, A.; Buecker, C.; Schafer, R.; Han, X.; Au, P.; et al. Generation of functionally competent and durable engineered blood vessels from human induced pluripotent stem cells. Proc. Nat. Acad. Sci. USA 2013, 110, 12774–12779. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.; Foldes, G.; Harding, S.; Mitchell, J. Stem cell-derived endothelial cells for cardiovascular disease: A therapeutic perspective. Br. J. Clin. Pharmacol. 2013, 75, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Au, P.; Chen, T.; Shao, Y.; Daheron, L.; Bai, H.; Arzigian, M.; Fukumura, D.; Jain, R.; Scadden, D. Endothelial cells derived from human embryonic stem cells form durable blood vessels in vivo. Nat. Biotechnol. 2007, 25, 317–318. [Google Scholar] [CrossRef] [PubMed]
- Bergers, G. The role of pericytes in blood-vessel formation and maintenance. Neuro Oncol. 2005, 7, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Van der Meer, A.; Orlova, V.; ten Dijke, P.; van den Berg, A.; Mummery, C. Three-dimensional co-cultures of human endothelial cells and embryonic stem cell-derived pericytes inside a microfluidic device. Lab Chip 2013, 13, 3562–3568. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Rayatpisheh, S.; Chew, S.; Chan-Park, M. Impact of endothelial cells on 3D cultured smooth muscle cells in a biomimetic hydrogel. ACS Appl. Mater. Interfaces 2012, 4, 1378–1387. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Chung, M.; Kim, S.; Jo, D.; Kim, J.; Jeon, N. Engineering of a biomimetic pericyte-covered 3D microvascular network. PLoS ONE 2015, 10, e0133880. [Google Scholar] [CrossRef] [PubMed]
- Boyd, N.; Nunes, S.; Jokinen, J.; Krishnan, L.; Chen, Y.; Smith, K.; Stice, S.; Hoying, J. Microvascular mural cell functionality of human embryonic stem cell derived mesenchymal cells. Tissue Eng. Part A 2011, 17, 1537–1548. [Google Scholar] [CrossRef] [PubMed]
- Booth, R.; Kim, H. Characterization of a microfluidic in vitro model of the blood-brain barrier (μBBB). Lab Chip 2012, 12, 1784–1792. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.; Pensabene, V.; Markov, D.; Allwardt, V.; Neely, M.; Shi, M.; Britt, C.; Hoilett, O.; Yang, Q.; Brewer, B.; et al. Recreating blood-brain barrier physiology and structure on chip: A novel neurovascular microfluidic bioreactor. Biomicrofluidics 2015, 9, 054124. [Google Scholar] [CrossRef] [PubMed]
- Herland, A.; van der Meer, A.; FitzGerald, E.; Park, T.; Sleeboom, J.; Ingber, D. Distinct contributions of astrocytes and pericytes to neuroinflammation identified in a 3D human blood-brain barrier on a chip. PLoS ONE 2016, 11, e0150360. [Google Scholar] [CrossRef] [PubMed]
- Jang, K.; Mehr, A.; Hamilton, G.; McPartlin, L.; Chung, S.; Suh, K.; Ingber, D. Human kidney proximal tubule-on-a-chip for drug transport and nephrotoxicity assessment. Integr. Biol. 2013, 5, 1119–1129. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.; Wang, Z.; Voellinger, J.; Yeung, C.; Shen, D.; Thummel, K.; Zheng, Y.; Ligresti, G.; Eaton, D.; Muczynski, K.; et al. Innovations in preclinical biology: Ex vivo engineering of a human kidney tissue microperfusion system. Stem Cell Res. Ther. 2013, 4, S17. [Google Scholar] [CrossRef] [PubMed]
- Prodanov, L.; Jindal, R.; Bale, S.; Hegde, M.; McCarty, W.; Golberg, I.; Bhushan, A.; Yarmush, M.; Usta, O. Long-term maintenance of a microfluidic 3D human liver sinusoid. Biotechnol. Bioeng. 2015, 113, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Hung, P.; Lee, L. An artificial liver sinusoid with a microfluidic endothelial-like barrier for primary hepatocyte culture. Biotechnol. Bioeng. 2007, 97, 1340–1346. [Google Scholar] [CrossRef] [PubMed]
- Birukova, A.; Chatchavalvanich, S.; Rios, A.; Kawkitinarong, K.; Garcia, J.; Birukov, K. Differential regulation of pulmonary endothelial monolayer integrity by varying degrees of cyclic stretch. Am. J. Pathol. 2006, 168, 1749–1761. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Mireles, M.; Kwarta, B.; Gaborski, T. Use of porous membranes in tissue barrier and co-culture models. Lab Chip 2018, 18, 1671–1689. [Google Scholar] [CrossRef] [PubMed]
- Pasman, T.; Grijpma, D.; Stamatialis, D.; Poot, A. Flat and microstructured polymeric membranes in organs-on-chips. J. R. Soc. Interface 2018, 15, 20180351. [Google Scholar] [CrossRef] [PubMed]
- Huh, D.; Kim, H.J.; Fraser, J.P.; Shea, D.E.; Khan, M.; Bahinski, A.; Hamilton, G.A.; Ingber, D.E. Microfabrication of human organs-on-chips. Nat. Protoc. 2013, 8, 2135–2157. [Google Scholar] [CrossRef] [PubMed]
- Abhyankar, V.V.; Wu, M.; Koh, C.Y.; Hatch, A.V.A. reversibly sealed, easy access, modular (SEAM) microfluidic architecture to establish in vitro tissue interfaces. PLoS ONE 2016, 11, e0156341. [Google Scholar] [CrossRef] [PubMed]
- Huh, D.; Matthews, B.; Mammoto, A.; Montoya-Zavala, M.; Hsin, H.; Ingber, D. Reconstituting organ-level lung functions on a chip. Science 2010, 328, 1662–1668. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Huh, D.; Hamilton, G.; Ingber, D. Human gut-on-a-chip inhabited by microbial flora that experiences intestinal peristalsis-like motions and flow. Lab Chip 2012, 12, 2165–2174. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Lv, Y.; Zeng, M.; Fu, B. Non-invasive measurement of solute permeability in cerebral microvessels of the rat. Microvasc. Res. 2009, 77, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zeng, M.; Sun, Y.; Fu, B. Quantification of blood-brain barrier solute permeability and brain transport by multiphoton microscopy. J. Biomech. Eng. 2014, 136, 031005. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, B.; Kolli, A.; Esch, M.; Abaci, H.; Shuler, M.; Hickman, J. TEER measurement techniques for in vitro barrier model systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef] [PubMed]
- Barakat, A. Blood flow and arterial endothelial dysfunction: Mechanisms and implications. C. R. Phys. 2013, 14, 479–496. [Google Scholar] [CrossRef]
- Hulkower, K.; Herber, R. Cell migration and invasion assays as tools for drug discovery. Pharmaceutics 2011, 3, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, J.; Seidel, C.; Ebner, R.; Kunz-Schughart, L. Spheroid-based drug screen: Considerations and practical approach. Nat. Protoc. 2009, 4, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Yabushita, H.; Ohnishi, M.; Komiyama, M.; Mori, T.; Noguchi, M.; Kishida, T.; Noguchi, Y.; Sawaguchi, K.; Noguchi, M. Usefulness of collagen gel droplet embedded culture drug sensitivity testing in ovarian cancer. Oncol. Rep. 2004, 12, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.; Kobayashi, H.; Rafii, S. Instructive role of the vascular niche in promoting tumour growth and tissue repair by angiocrine factors. Nat. Rev. Cancer 2010, 10, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Chaffer, C.; Weinberg, R. A perspective on cancer cell metastasis. Science 2011, 331, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- Weis, S.M.; Cheresh, D.A. Tumor angiogenesis: Molecular pathways and therapeutic targets. Nat. Med. 2011, 17, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Chiang, S.P.; Cabrera, R.M.; Segall, J.E. Tumor cell intravasation. Am. J. Physiol. Cell Physiol. 2016, 311, C1–C14. [Google Scholar] [CrossRef] [PubMed]
- Stoletov, K.; Kato, H.; Zardouzian, E.; Kelber, J.; Yang, J.; Shattil, S.; Klemke, R. Visualizing extravasation dynamics of metastatic tumor cells. J. Cell Sci. 2010, 123, 2332–2341. [Google Scholar] [CrossRef] [PubMed]
- Fukumura, D.; Duda, D.; Munn, L.; Jain, R. Tumor microvasculature and microenvironment: Novel insights through intravital imaging in pre-clinical models. Microcirculation 2010, 17, 206–225. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Park, W.; Ryu, H.; Jeon, N. A microfluidic platform for quantitative analysis of cancer angiogenesis and intravasation. Biomicrofluidics 2014, 8, 054102. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, C.; Verbridge, S.; Vlachos, P.; Rylander, M. Flow shear stress regulates endothelial barrier function and expression of angiogenic factors in a 3D microfluidic tumor vascular model. Cell Adh. Migr. 2014, 8, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Ehsan, S.M.; Welch-Reardon, K.M.; Waterman, M.L.; Hughes, C.C.; George, S.C. A three-dimensional in vitro model of tumor cell intravasation. Integr. Biol 2014, 6, 603–610. [Google Scholar] [CrossRef] [PubMed]
- DelNero, P.; Lane, M.; Verbridge, S.S.; Kwee, B.; Kermani, P.; Hempstead, B.; Stroock, A.; Fischbach, C. 3D culture broadly regulates tumor cell hypoxia response and angiogenesis via pro-inflammatory pathways. Biomaterials 2015, 55, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Zervantonakis, I.; Hughes-Alford, S.; Charest, J.; Condeelis, J.; Gertler, F.; Kamm, R. Three-dimensional microfluidic model for tumor cell intravasation and endothelial barrier function. Proc. Nat. Acad. Sci. USA 2012, 109, 13515–13520. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.; Searson, P. Live-cell imaging of invasion and intravasation in an artificial microvessel platform. Cancer Res. 2014, 74, 4937–4945. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Whisler, J.; Jeon, J.; Kamm, R. Mechanisms of tumor cell extravasation in an in vitro microvascular network platform. Integr. Biol. 2013, 5, 1262–1271. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.; Bersini, S.; Gilardi, M.; Dubini, G.; Charest, J.; Moretti, M.; Kamm, R. Human 3D vascularized organotypic microfluidic assays to study breast cancer cell extravasation. Proc. Nat. Acad. Sci. USA 2015, 112, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Cai, B.; Lei, M.; Liu, Y.; Fu, B.M. Differential arrest and adhesion of tumor cells and microbeads in the microvasculature. Biomech. Model. Mechanobiol. 2014, 13, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Polacheck, W.J.; Charest, J.L.; Kamm, R.D. Interstitial flow influences direction of tumor cell migration through competing mechanisms. Proc. Natl. Acad. Sci. USA 2011, 108, 11115–11120. [Google Scholar] [CrossRef] [PubMed]
- Carey, S.P.; Kraning-Rush, C.M.; Williams, R.M.; Reinhart-King, C.A. Biophysical control of invasive tumor cell behavior by extracellular matrix microarchitecture. Biomaterials 2012, 33, 4157–4165. [Google Scholar] [CrossRef] [PubMed]
- Foda, H.D.; Zucker, S. Matrix metalloproteinases in cancer invasion, metastasis and angiogenesis. Drug Discov. Today 2001, 6, 478–482. [Google Scholar] [CrossRef]
- Raman, D.; Baugher, P.J.; Thu, Y.M.; Richmond, A. Role of chemokines in tumor growth. Cancer Lett. 2007, 256, 137–165. [Google Scholar] [CrossRef] [PubMed]
- Esch, E.W.; Bahinski, A.; Huh, D. Organs-on-chips at the frontiers of drug discovery. Nat. Rev. Drug Discov. 2015, 14, 248–260. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Gao, Y.; Hao, Y.; Li, E.; Wang, Y.; Zhang, J.; Wang, W.; Gao, Z.; Wang, Q. Application of a microfluidic chip-based 3d co-culture to test drug sensitivity for individualized treatment of lung cancer. Biomaterials 2013, 34, 4109–4117. [Google Scholar] [CrossRef] [PubMed]
- Sobrino, A.; Phan, D.; Datta, R.; Wang, X.; Hachey, S.; Romero-López, M.; Gratton, E.; Lee, A.; George, S.; Hughes, C. 3D microtumors in vitro supported by perfused vascular networks. Sci. Rep. 2016, 6, 31589. [Google Scholar] [CrossRef] [PubMed]
- Phan, D.; Wang, X.; Craver, B.; Sobrino, A.; Zhao, D.; Chen, J.; Lee, L.; George, S.; Lee, A.; Hughes, C. A vascularized and perfused organ-on-a-chip platform for large-scale drug screening applications. Lab Chip 2017, 17, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Jain, R. Normalization of tumor vasculature: An emerging concept in antiangiogenic therapy. Science 2005, 307, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Boussommier-Calleja, A.; Li, R.; Chen, M.B.; Wong, S.C.; Kamm, R.D. Microfluidics: A new tool for modeling cancer-immune interactions. Trends Cancer 2016, 2, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Mattei, F.; Schiavoni, G.; De Ninno, A.; Lucarini, V.; Sestili, P.; Sistigu, A.; Fragale, A.; Sanchez, M.; Spada, M.; Gerardino, A.; et al. A multidisciplinary study using in vivo tumor models and microfluidic cell-on-chip approach to explore the cross-talk between cancer and immune cells. J. Immunotoxicol. 2014, 11, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Montgomery, M.; Chamberlain, M.; Ogawa, S.; Korolj, A.; Pahnke, A.; Wells, L.; Massé, S.; Kim, J.; Reis, L.; et al. Biodegradable scaffold with built-in vasculature for organ-on-a-chip engineering and direct surgical anastomosis. Nat. Mater. 2016, 15, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Van Midwoud, P.M.; Janse, A.; Merema, M.T.; Groothuis, G.M.; Verpoorte, E. Comparison of biocompatibility and adsorption properties of different plastics for advanced microfluidic cell and tissue culture models. Anal. Chem. 2012, 84, 3938–3944. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Vessel Parameters | Effect |
|---|---|---|
| Shear stress | Barrier function | Decreasing permeability [12,55,56,57] |
| Cell orientation | Elongated and aligned along flow direction in EC lining [57] | |
| EC invasion | Attenuate invasion [58,59] | |
| Interstitial flow | Vasculogenesis | Promote capillary formation regardless of flow direction [40] |
| Angiogenesis | More active in reverse direction of interstitial flow [51] | |
| Stiff ECM | Vessel lumen | Small diameter [63] |
| Sprouting invasion | Significantly reduced [70] | |
| Fiber arrangements of ECM | Vessel alignment | Along the fiber orientation [64] |
| VEGF concentration | Endothelial sprouting | Induce sprouting at low concentration (2.5–5 ng/mL) [67] |
| VEGF gradient | Vessel morphology | Positive gradient induces sprouting, and negative gradient induces vessel dilation [59] |
| Hypoxia | Angiogenesis | Promote angiogenesis, such as tumor angiogenesis [78] |
| Co-culture with fibroblast | Perfusable vascular formation | Promote EC sprouting and lumen formation [40,52] |
| Co-culture with mural cells | Vessel stabilization | More stable [88,89,90,91] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Sun, Q.; Pei, J. Microfluidic-Based 3D Engineered Microvascular Networks and Their Applications in Vascularized Microtumor Models. Micromachines 2018, 9, 493. https://doi.org/10.3390/mi9100493
Wang X, Sun Q, Pei J. Microfluidic-Based 3D Engineered Microvascular Networks and Their Applications in Vascularized Microtumor Models. Micromachines. 2018; 9(10):493. https://doi.org/10.3390/mi9100493
Chicago/Turabian StyleWang, Xiaolin, Qiyue Sun, and Jianghua Pei. 2018. "Microfluidic-Based 3D Engineered Microvascular Networks and Their Applications in Vascularized Microtumor Models" Micromachines 9, no. 10: 493. https://doi.org/10.3390/mi9100493
APA StyleWang, X., Sun, Q., & Pei, J. (2018). Microfluidic-Based 3D Engineered Microvascular Networks and Their Applications in Vascularized Microtumor Models. Micromachines, 9(10), 493. https://doi.org/10.3390/mi9100493