

Natural Organic Materials Based Memristors and Transistors for Artificial Synaptic Devices in Sustainable Neuromorphic Computing Systems


Abstract
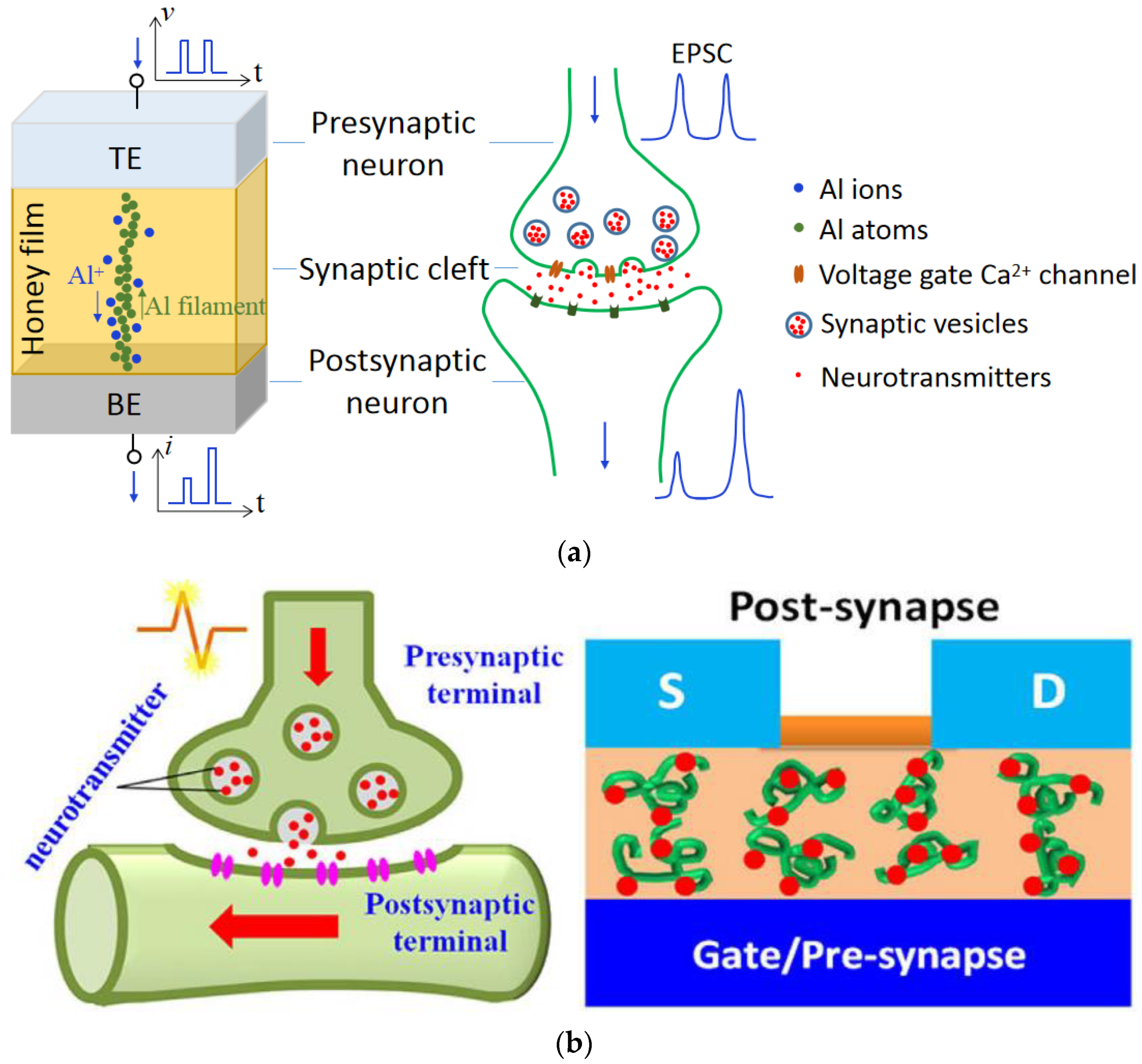
1. Introduction
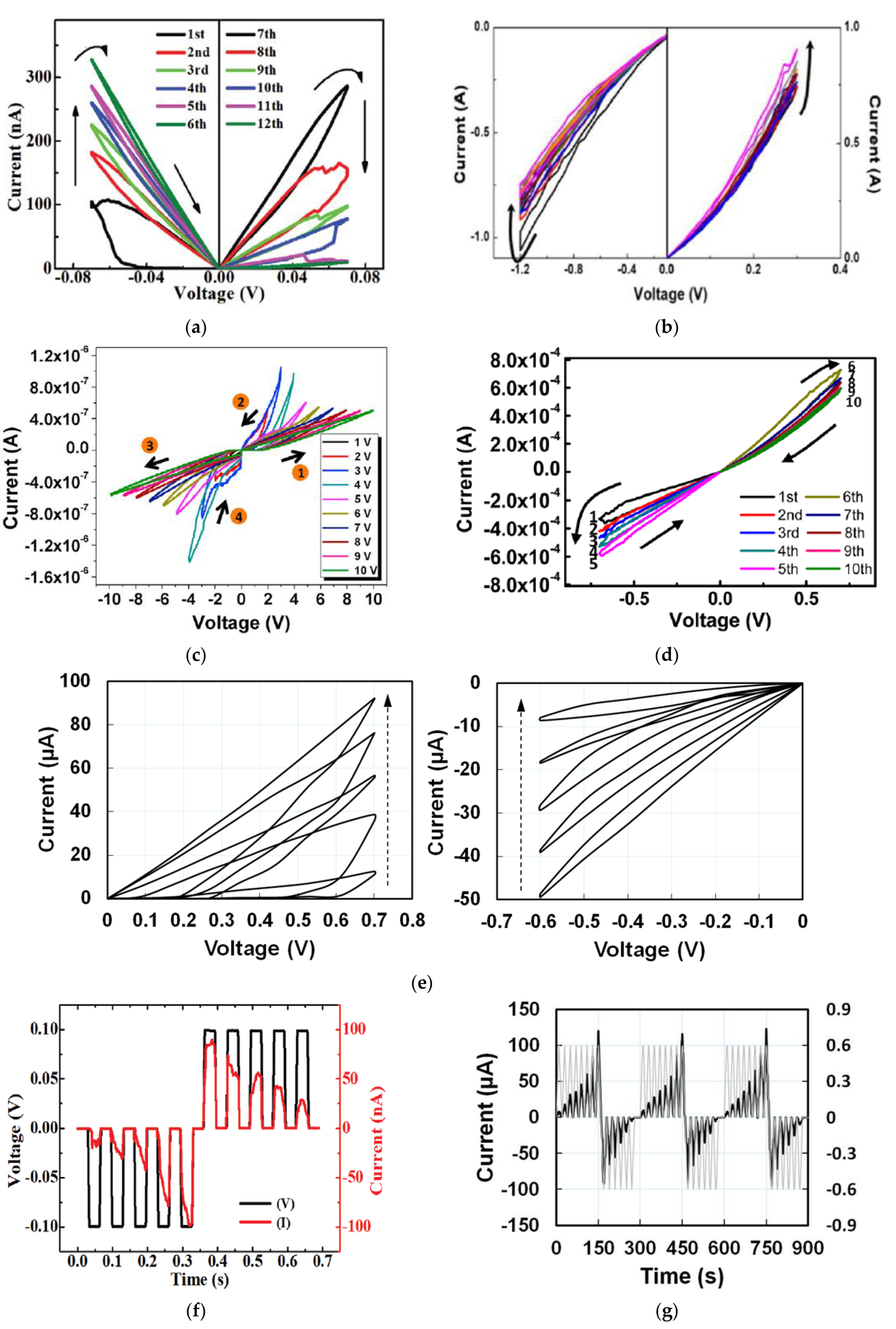
2. Analog Memristive Behaviors
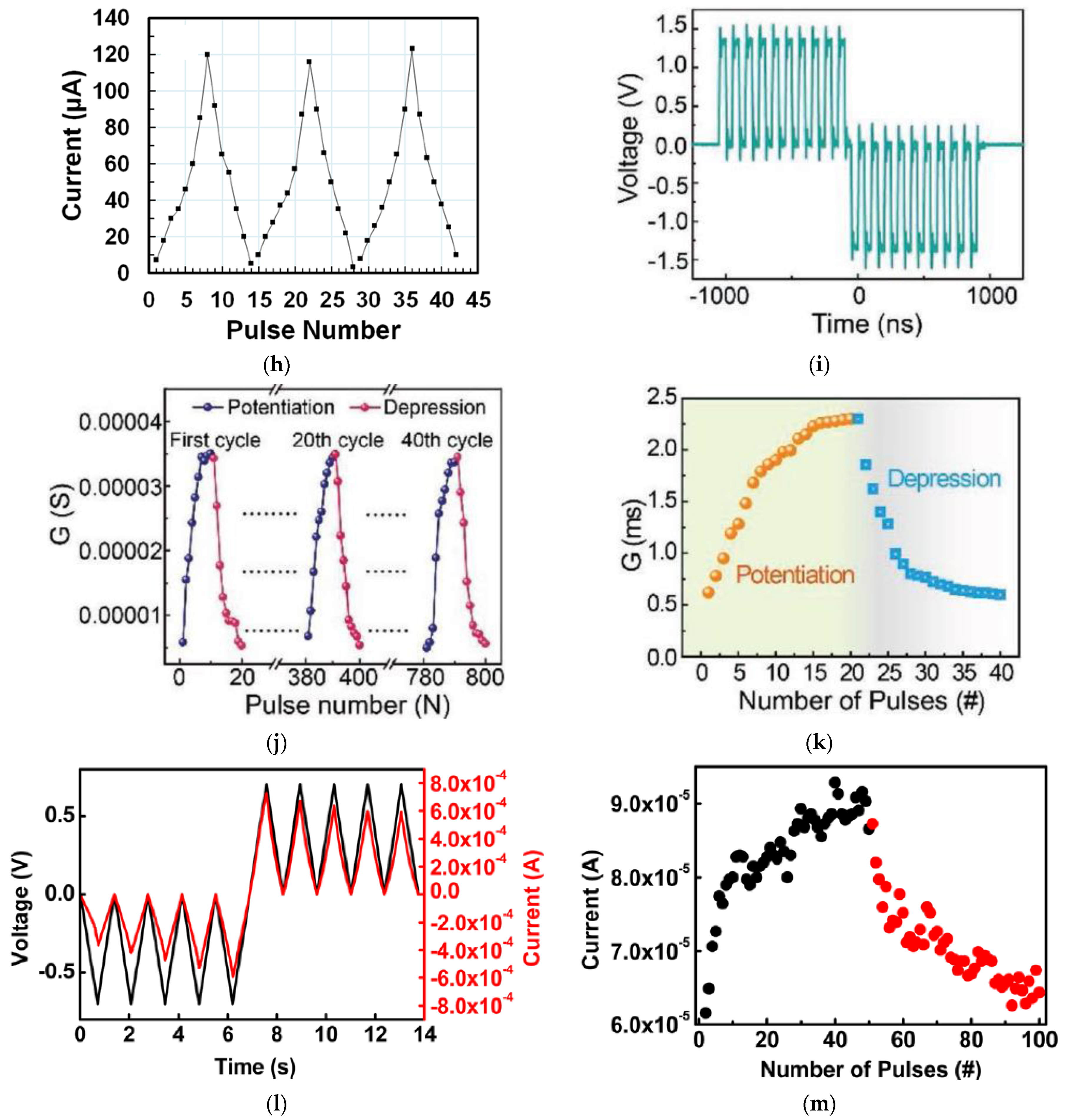
3. Synaptic Plasticity
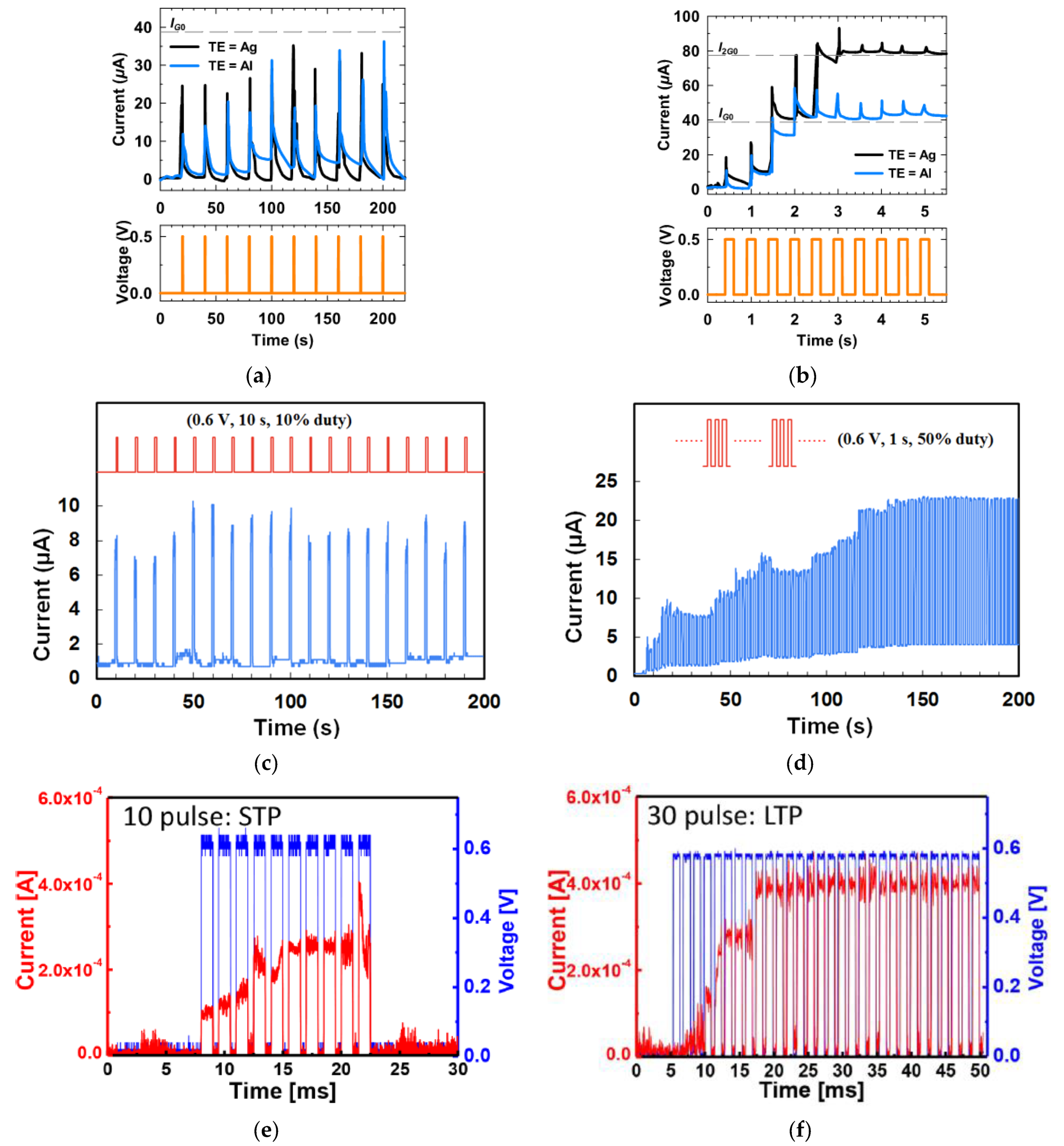
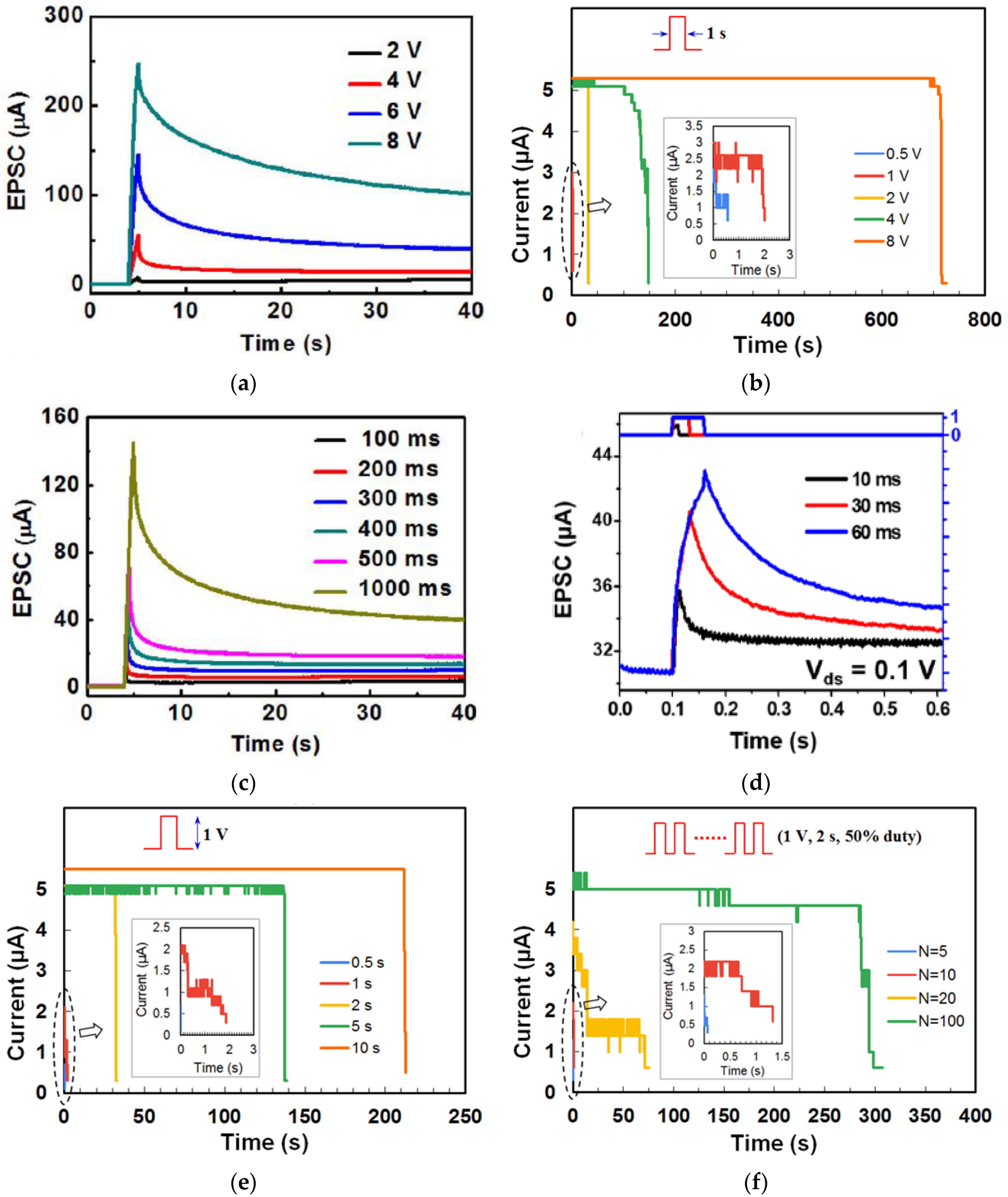
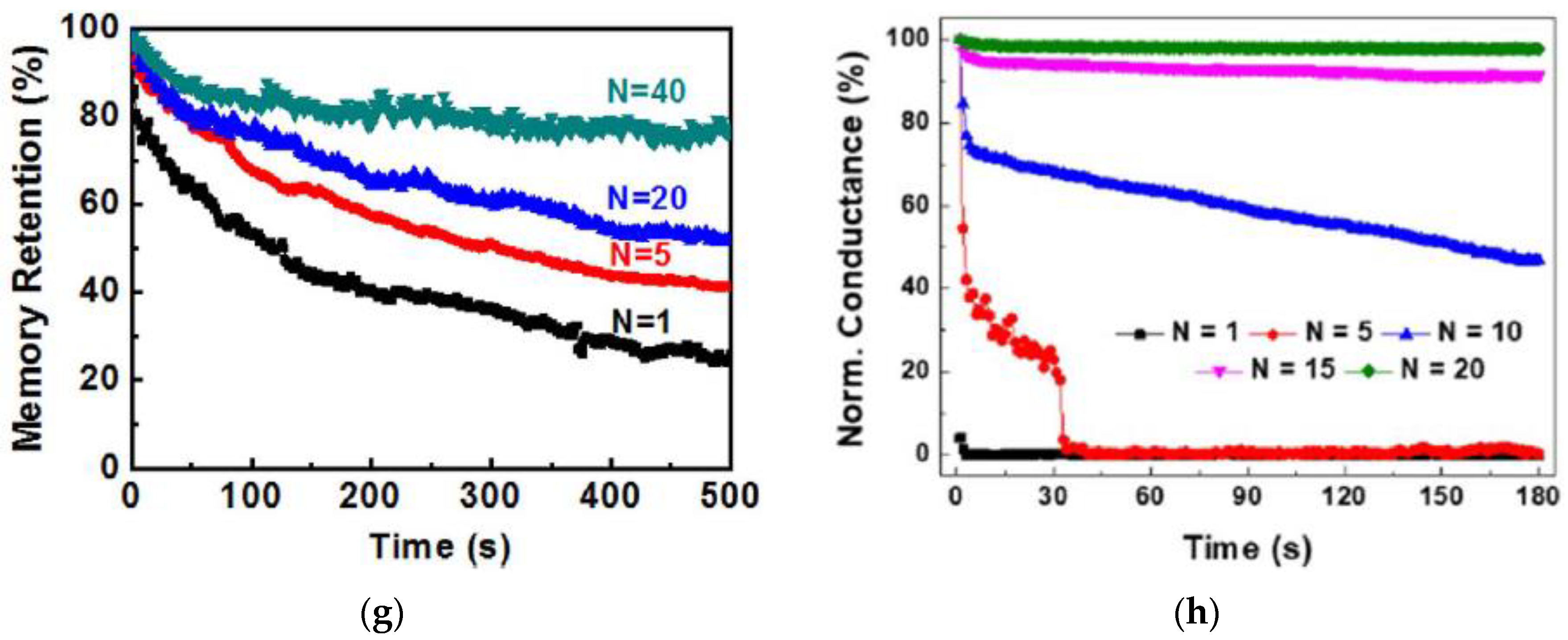
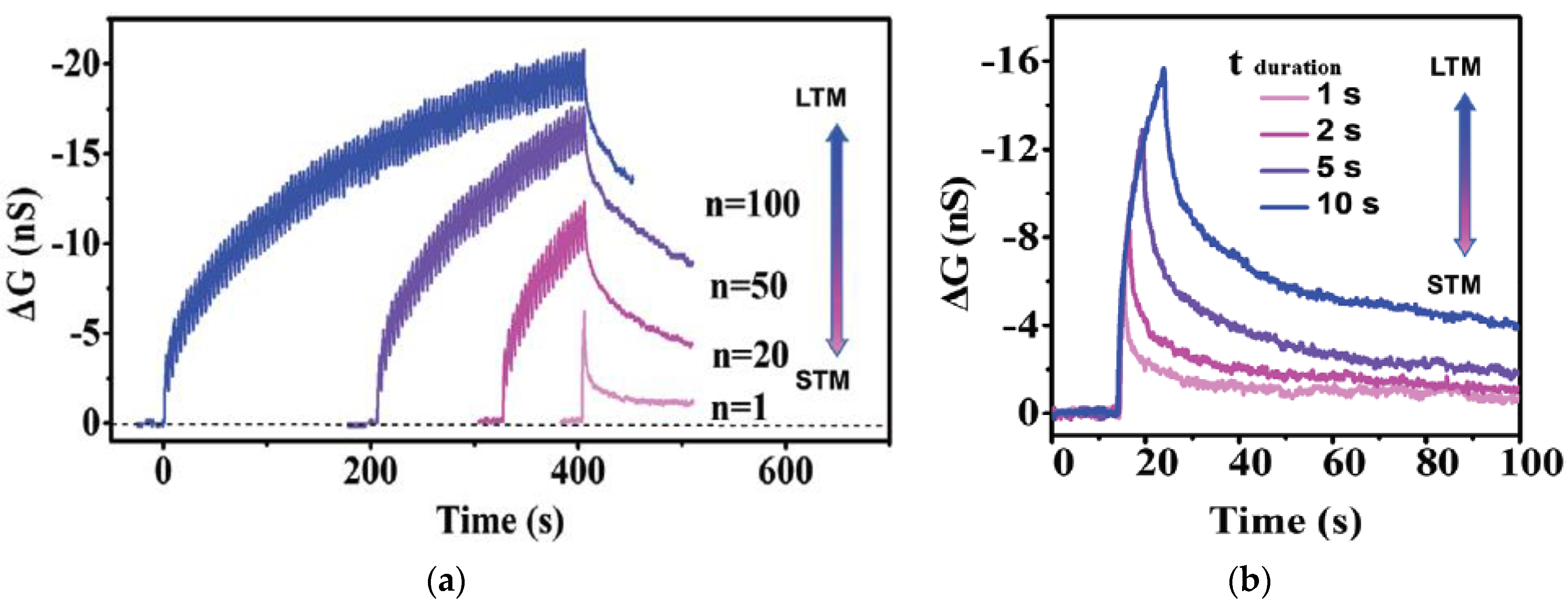
3.1. Short Term Memory and Long Term Memory
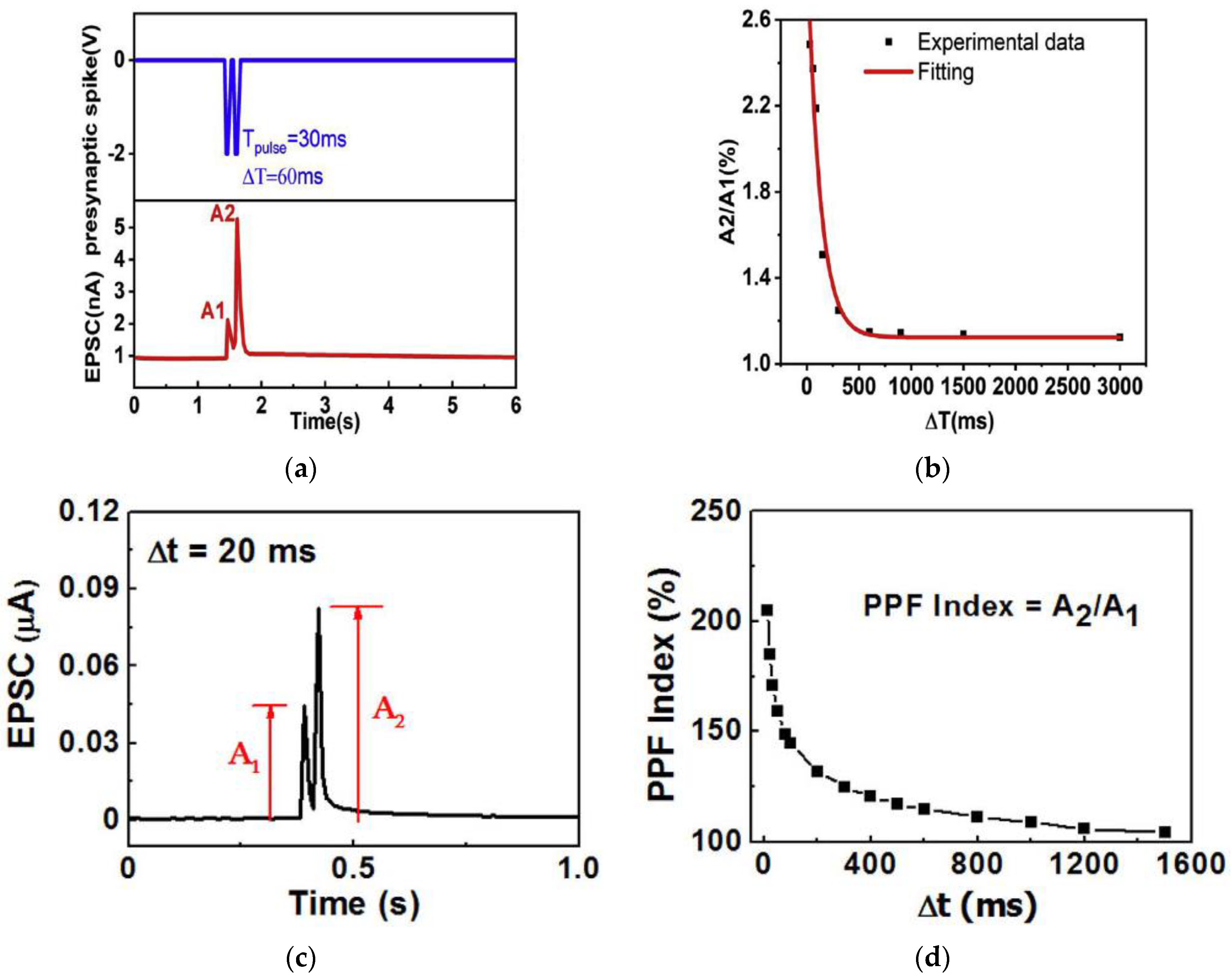
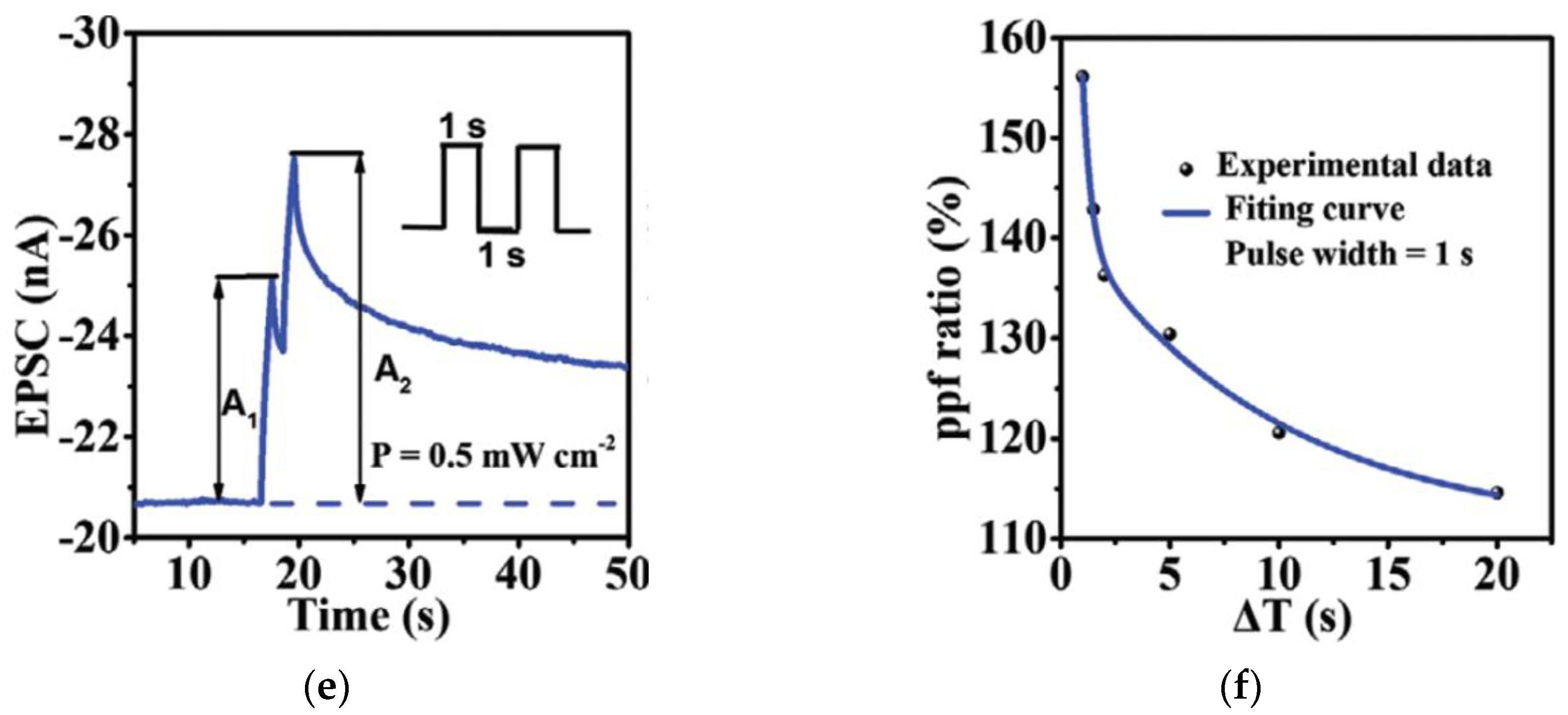
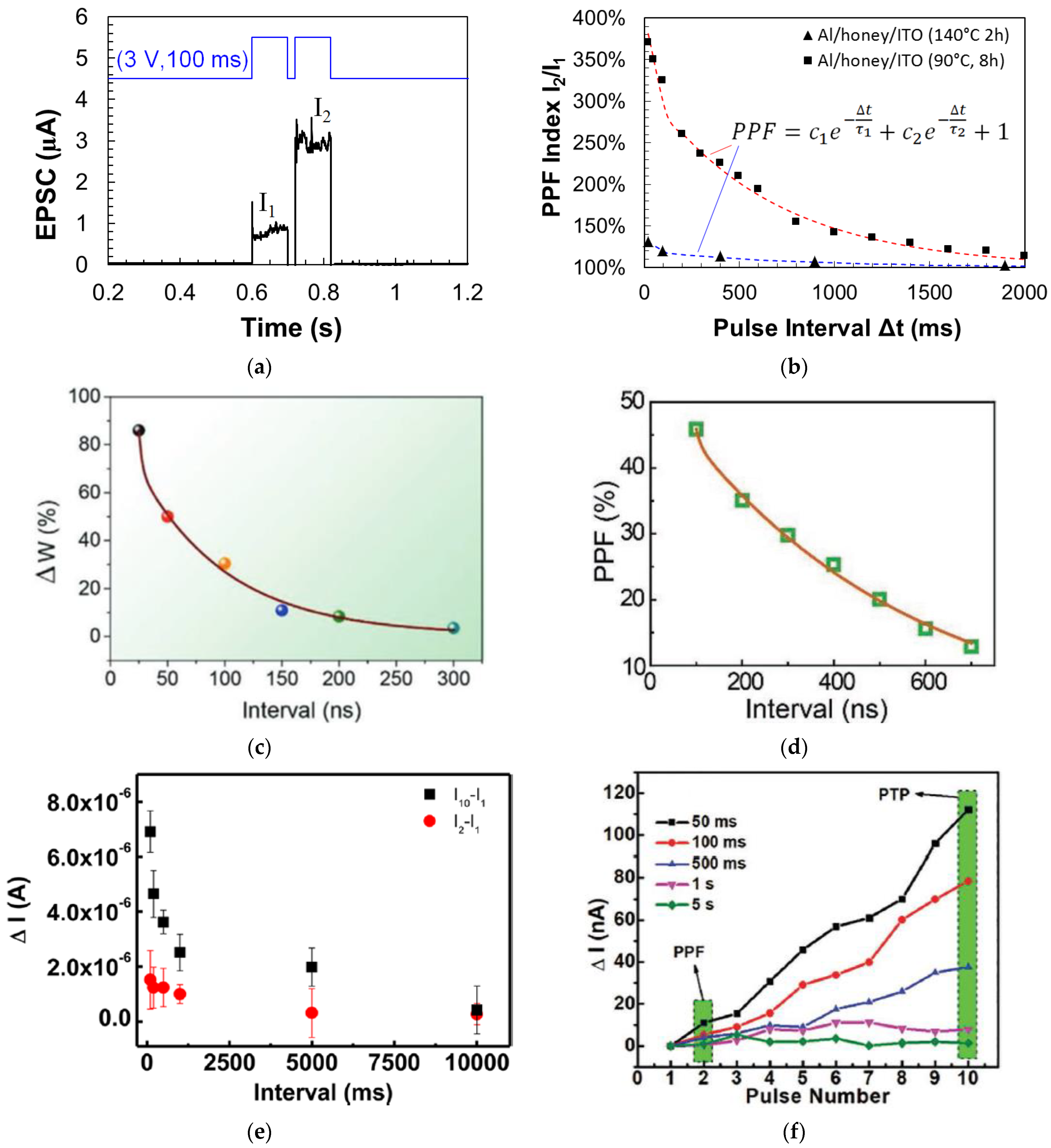
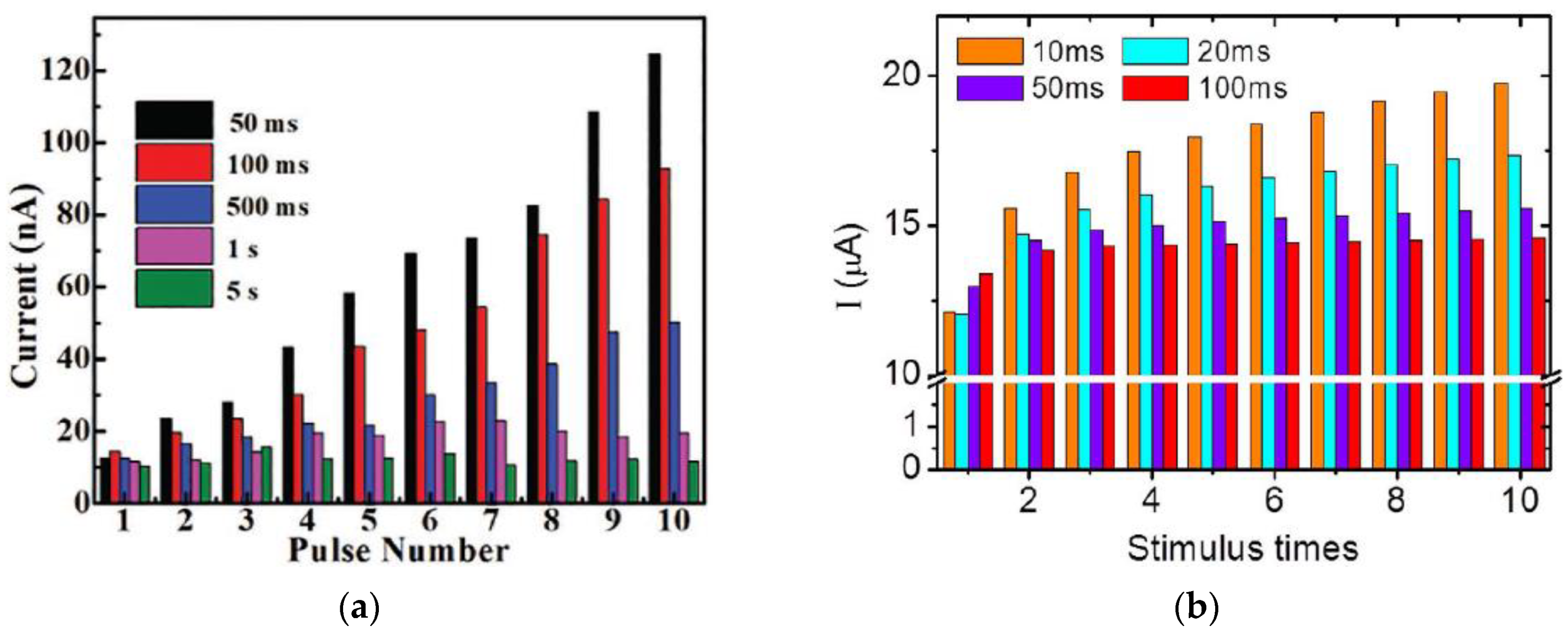
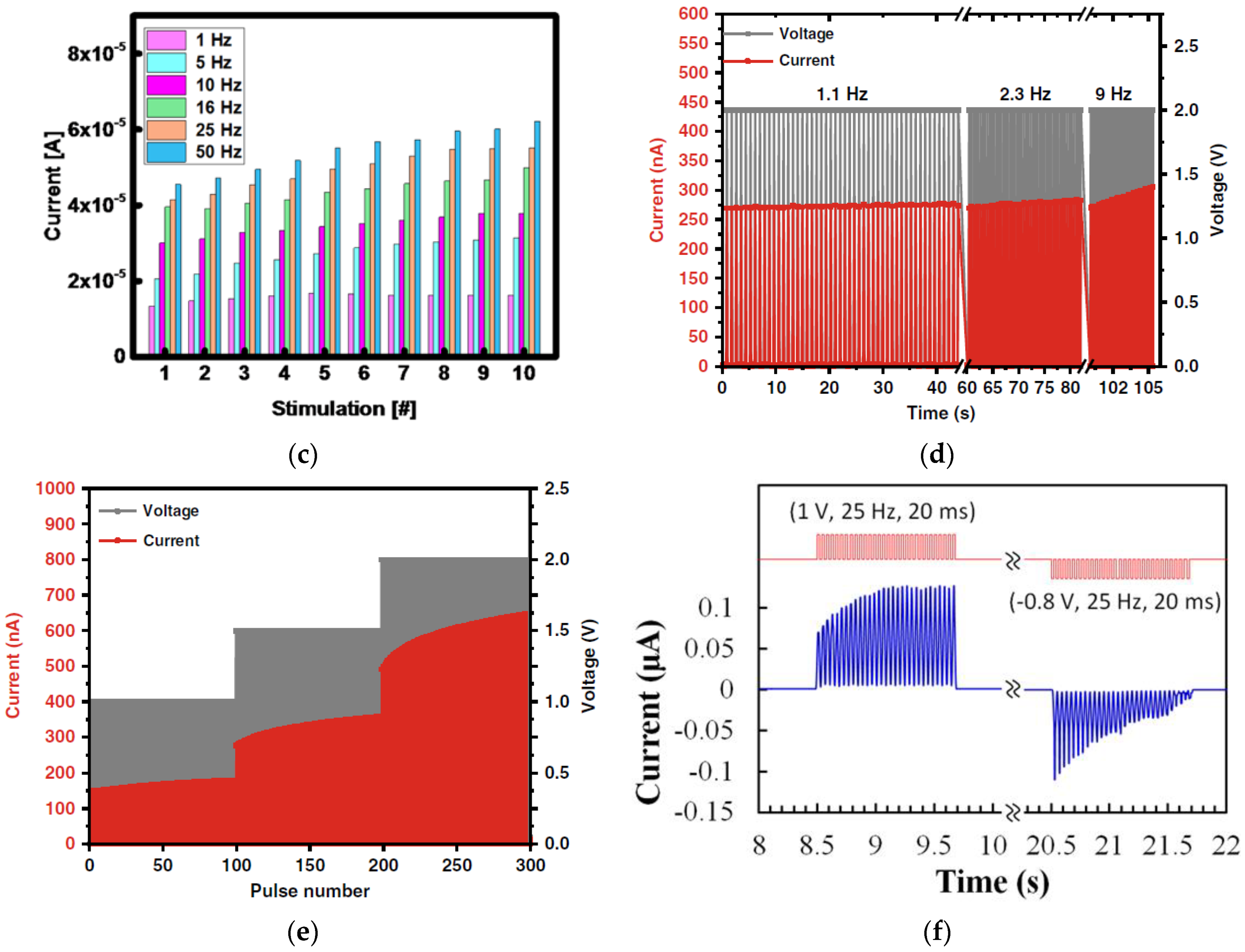
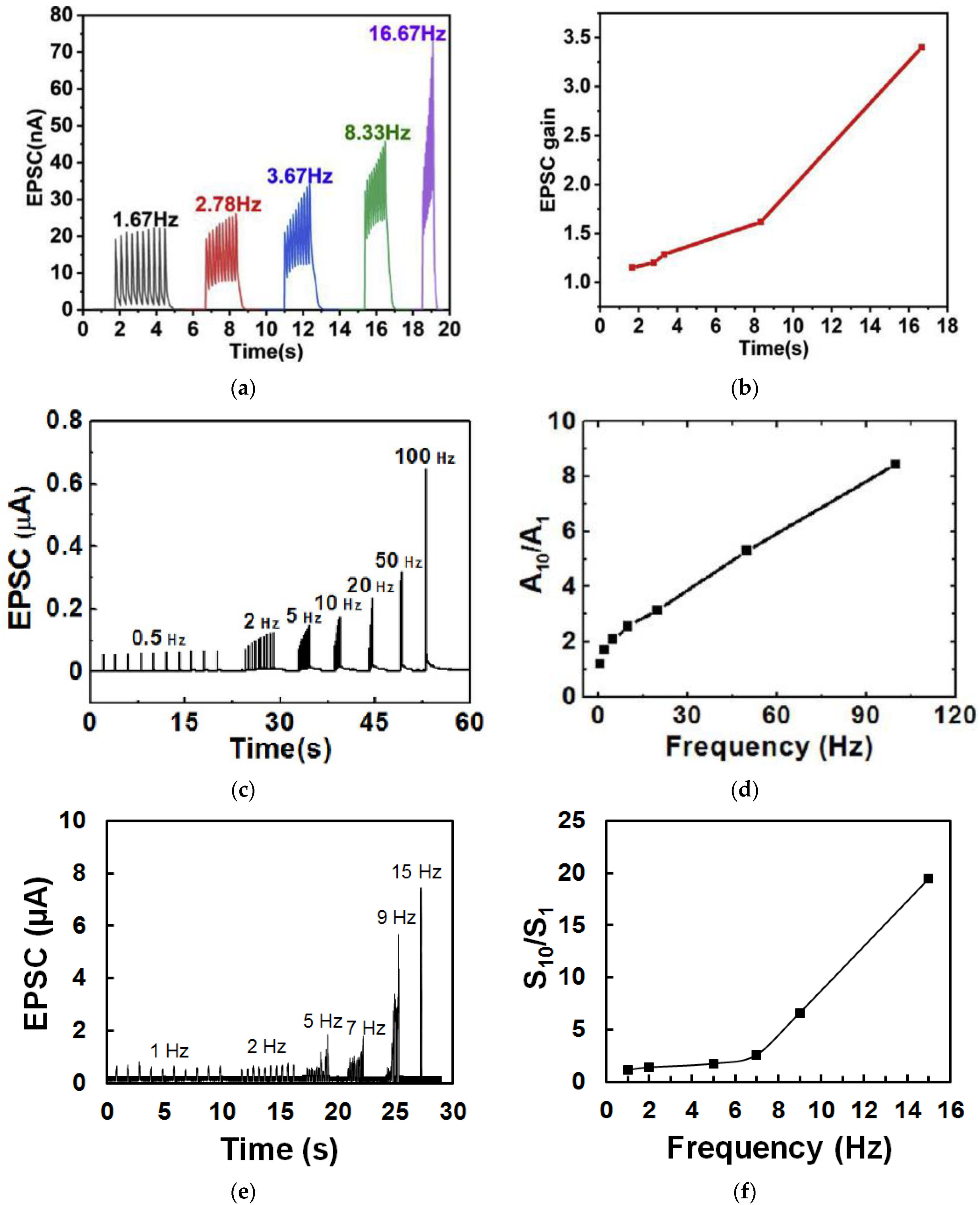
3.2. Paired Pulse Facilitation
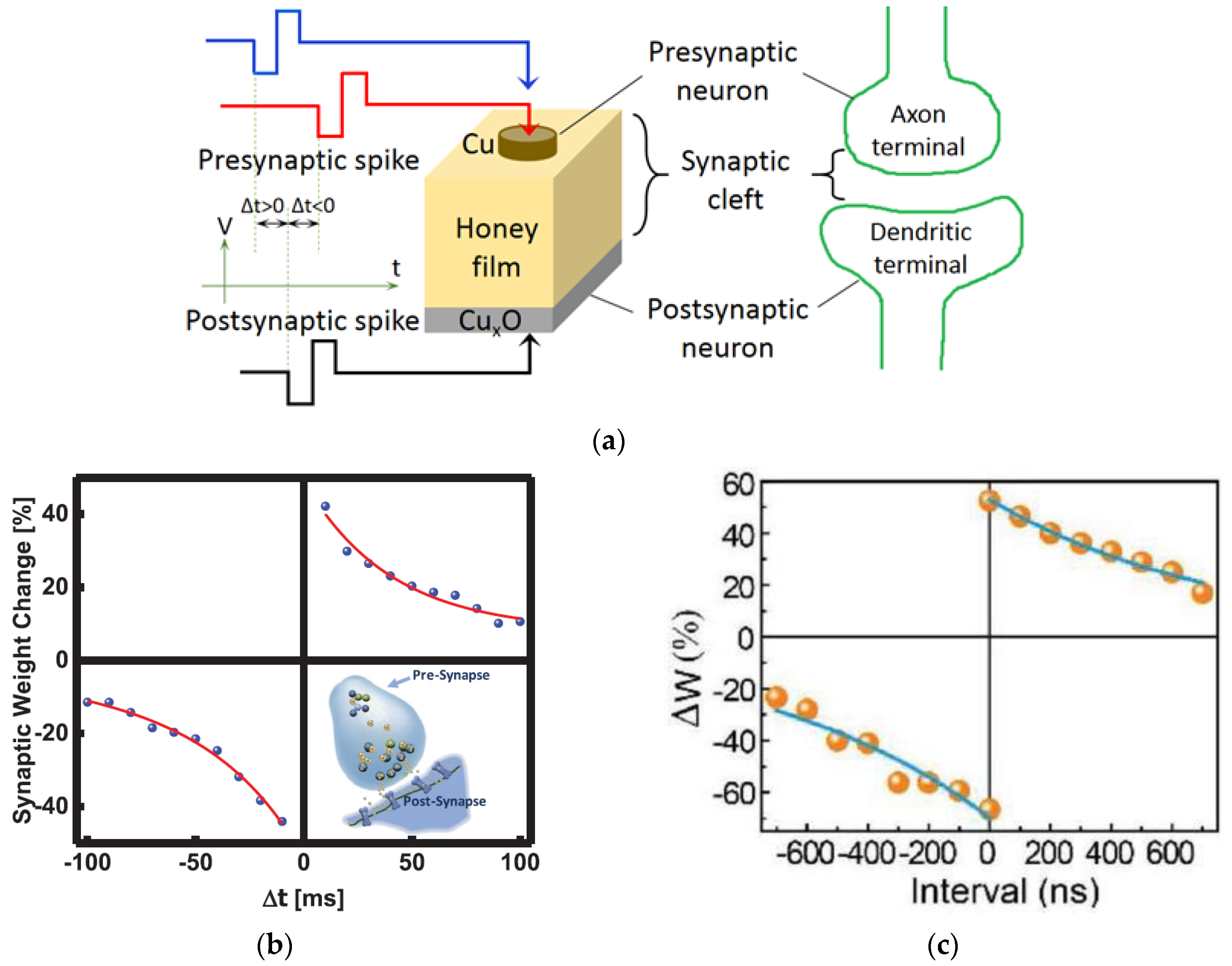
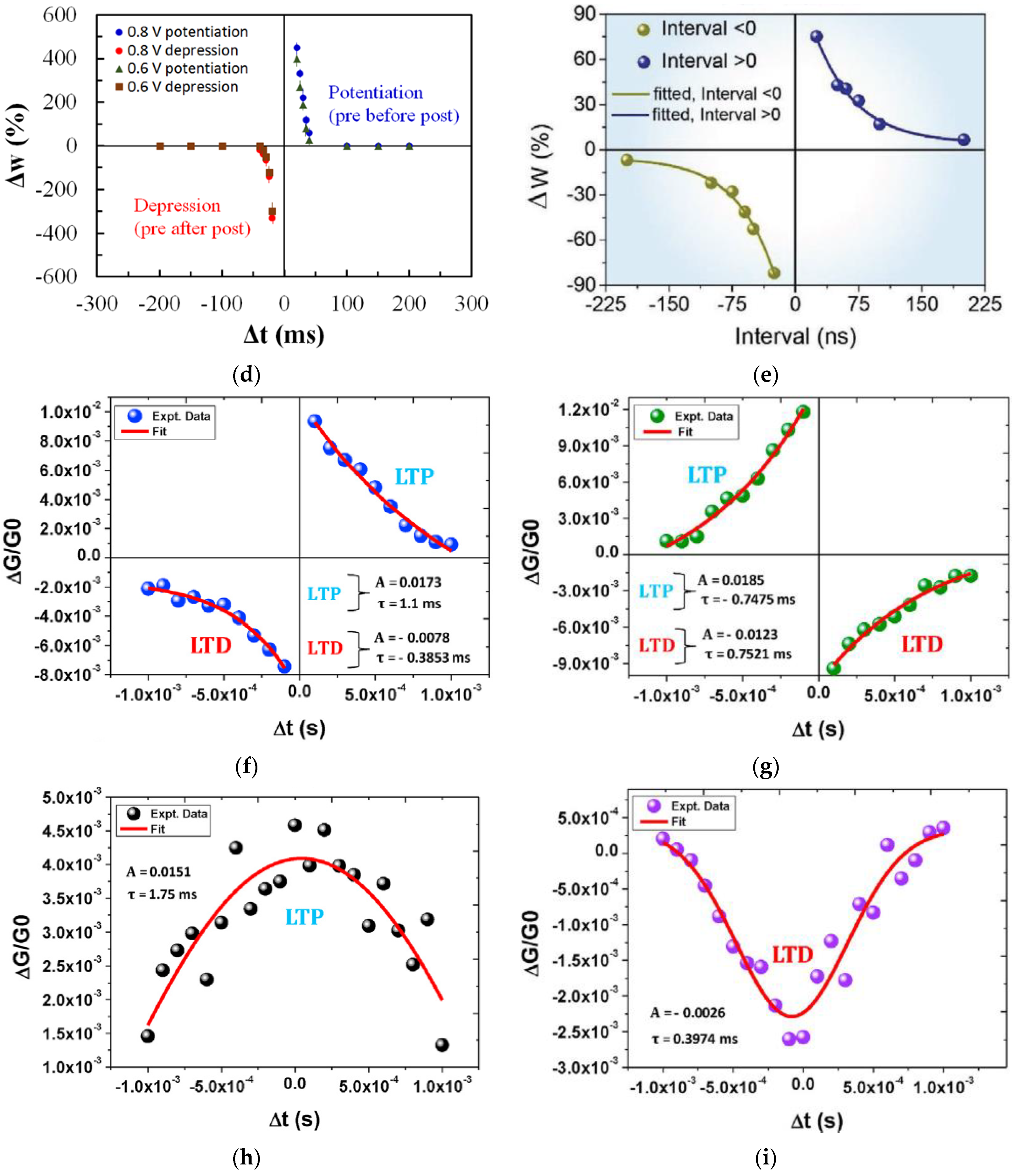
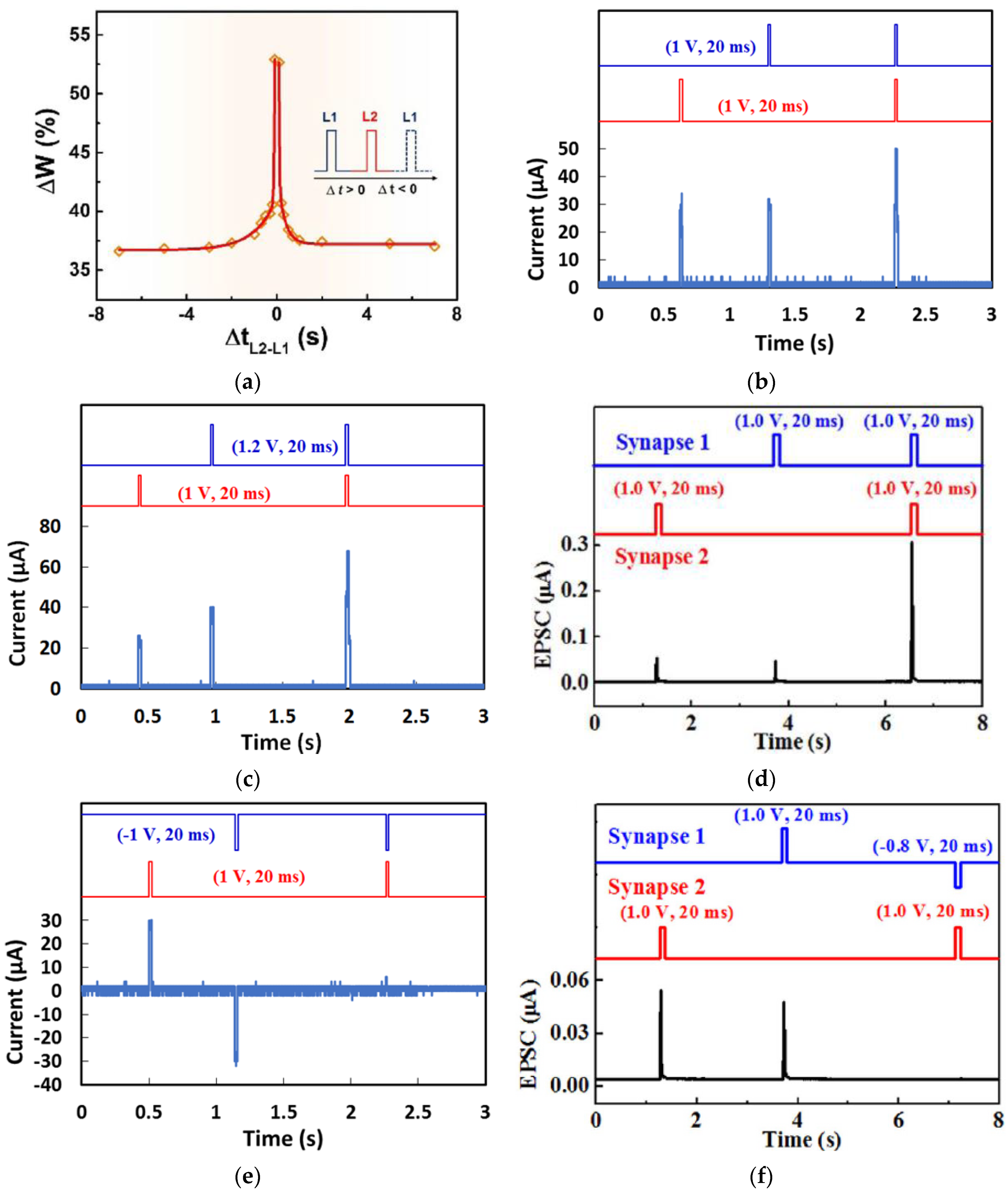
3.3. Spike-Timing-Dependent Plasticity
3.4. Spike-Rate-Dependent Plasticity
3.5. Dynamic Filtering
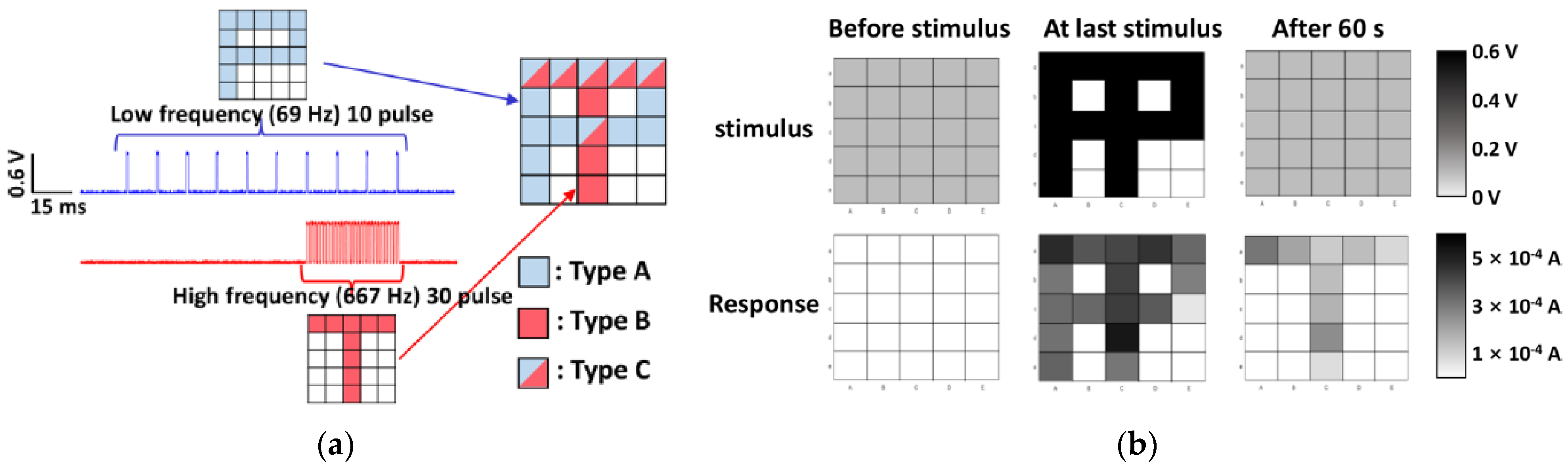
3.6. Spatial Summation
4. Device Fabrication
5. Mechanisms
6. Future Prospects
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Calimera, A.; Macii, E.; Poncino, M. The Human Brain Project and neuromorphic computing. Funct. Neurol. 2013, 28, 191–196. [Google Scholar] [CrossRef]
- Wang, J.; Mao, S.; Zhu, S.; Hou, W.; Yang, F.; Sun, B. Biomemristors-based synaptic devices for artificial intelligence applications. Org. Electron. 2022, 106, 106540. [Google Scholar] [CrossRef]
- Lu, Q.; Sun, F.; Liu, L.; Li, L.; Wang, Y.; Hao, M.; Wang, Z.; Wang, S.; Zhang, T. Biological receptor-inspired flexible artificial synapse based on ionic dynamics. Microsyst. Nanoeng. 2020, 6, 84. [Google Scholar] [CrossRef]
- Fu, W.-H.; Li, J.; Jiang, D.-L.; Yang, Y.-H.; Chen, Q.; Zhu, W.-Q.; Zhang, J.-H. Proton conducting C3N4/Chitosan composite electrolytes based InZnO thin film transistor for artificial synapse. Org. Electron. 2020, 85, 105870. [Google Scholar] [CrossRef]
- Feng, P.; Du, P.; Wan, C.; Shi, Y.; Wan, Q. Proton Conducting Graphene Oxide/Chitosan Composite Electrolytes as Gate Dielectrics for New-Concept Devices. Sci. Rep. 2016, 6, srep34065. [Google Scholar] [CrossRef]
- Min, S.-Y.; Cho, W.-J. Memristive Switching Characteristics in Biomaterial Chitosan-Based Solid Polymer Electrolyte for Artificial Synapse. Int. J. Mol. Sci. 2021, 22, 773. [Google Scholar] [CrossRef]
- Lim, Z.X.; Sreenivasan, S.; Wong, Y.H.; Zhao, F.; Cheong, K.Y. Effects of Electrode Materials on Charge Conduction Mechanisms of Memory Device Based on Natural Aloe Vera. MRS Adv. 2016, 1, 2513–2518. [Google Scholar] [CrossRef]
- Lim, Z.X.; Sreenivasan, S.; Wong, Y.H.; Zhao, F.; Cheong, K.Y. Filamentary Conduction in Aloe Vera Film for Memory Application. Procedia Eng. 2017, 184, 655–662. [Google Scholar] [CrossRef]
- Lim, Z.X.; Tayeb, I.A.; Hamid, Z.A.A.; Ain, M.F.; Hashim, A.M.; Abdullah, J.M.; Zhao, F.; Cheong, K.Y. Switching Dynamics and Conductance Quantization of $Aloe$ Polysaccharides-Based Device. IEEE Trans. Electron Devices 2019, 66, 3110–3117. [Google Scholar] [CrossRef]
- Lim, Z.X.; Tayeb, I.A.; Hamid, Z.A.A.; Ain, M.F.; Hashim, A.M.; Abdullah, J.M.; Sivkov, A.A.; Zhao, F.; Cheong, K.Y. Artificial Synaptic Behavior of Aloe Polysaccharides-Based Device with Au as Top Electrode. MRS Adv. 2020, 5, 693–698. [Google Scholar] [CrossRef]
- Tayeb, I.A.; Zhao, F.; Abdullah, J.M.; Cheong, K.Y. Resistive switching behaviour in a polymannose film for multistate non-volatile memory application. J. Mater. Chem. C 2020, 9, 1437–1450. [Google Scholar] [CrossRef]
- Cheong, K.Y.; Tayeb, I.A.; Zhao, F.; Abdullah, J.M. Review on resistive switching mechanisms of bio-organic thin film for non-volatile memory application. Nanotechnol. Rev. 2021, 10, 680–709. [Google Scholar] [CrossRef]
- Yap, P.L.; Cheong, K.Y.; Lee, H.L.; Zhao, F. Effects of drying temperature on preparation of pectin polysaccharide thin film for resistive switching memory. J. Mater. Sci. Mater. Electron. 2022, 33, 19805–19826. [Google Scholar] [CrossRef]
- Kim, Y.; Park, C.H.; An, J.S.; Choi, S.-H.; Kim, T.W. Biocompatible artificial synapses based on a zein active layer obtained from maize for neuromorphic computing. Sci. Rep. 2021, 11, 20633. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.; Li, E.; Yan, Y.; Liu, Y.; Zhong, J.; Lv, D.; Ke, Y.; Chen, H.; Guo, T. Gelatin-hydrogel based organic synaptic transistor. Org. Electron. 2019, 75, 105409. [Google Scholar] [CrossRef]
- Sivkov, A.A.; Xing, Y.; Cheong, K.Y.; Zeng, X.; Zhao, F. Investigation of honey thin film as a resistive switching material for nonvolatile memories. Mater. Lett. 2020, 271, 127796. [Google Scholar] [CrossRef]
- Sueoka, B.; Cheong, K.Y.; Zhao, F. Natural biomaterial honey-based resistive switching device for artificial synapse in neuromorphic systems. Appl. Phys. Lett. 2022, 120, 083301. [Google Scholar] [CrossRef]
- Sueoka, B.; Zhao, F. Memristive synaptic device based on a natural organic material—Honey for spiking neural network in biodegradable neuromorphic systems. J. Phys. D Appl. Phys. 2022, 55, 225105. [Google Scholar] [CrossRef]
- Sueoka, B.; Cheong, K.Y.; Zhao, F. Study of synaptic properties of honey thin film for neuromorphic systems. Mater. Lett. 2021, 308, 131169. [Google Scholar] [CrossRef]
- Sueoka, B.; Tanim, M.H.; Williams, L.; Xiao, Z.; Seah, Y.Z.; Cheong, K.Y.; Zhao, F. A synaptic memristor based on natural organic honey with neural facilitation. Org. Electron. 2022, 109, 106622. [Google Scholar] [CrossRef]
- Tanim, M.H.; Sueoka, B.; Xiao, Z.; Cheong, K.Y.; Zhao, F. Study of carbon nanotube embedded honey as a resistive switching material. Nanotechnology 2022, 33, 495705. [Google Scholar] [CrossRef] [PubMed]
- Sueoka, B.; Vicenciodelmoral, A.Y.; Tanim, M.H.; Zhao, X.; Zhao, F. Correlation of natural honey-based RRAM processing and switching properties by experimental study and machine learning. Solid-State Electron. 2022, 197, 108463. [Google Scholar] [CrossRef]
- Wu, W.; Han, S.-T.; Venkatesh, S.; Sun, Q.; Peng, H.; Zhou, Y.; Yeung, C.; Li, R.K.; Roy, V. Biodegradable skin-inspired nonvolatile resistive switching memory based on gold nanoparticles embedded alkali lignin. Org. Electron. 2018, 59, 382–388. [Google Scholar] [CrossRef]
- Park, Y.; Lee, J.-S. Artificial Synapses with Short- and Long-Term Memory for Spiking Neural Networks Based on Renewable Materials. ACS Nano 2017, 11, 8962–8969. [Google Scholar] [CrossRef] [PubMed]
- Raeis-Hosseini, N.; Park, Y.; Lee, J.-S. Flexible Artificial Synaptic Devices Based on Collagen from Fish Protein with Spike-Timing-Dependent Plasticity. Adv. Funct. Mater. 2018, 28, 1800553. [Google Scholar] [CrossRef]
- Desai, T.R.; Dongale, T.D.; Patil, S.R.; Tiwari, A.P.; Pawar, P.K.; Kamat, R.K.; Kim, T.G. Synaptic learning functionalities of inverse biomemristive device based on trypsin for artificial intelligence application. J. Mater. Res. Technol. 2021, 11, 1100–1110. [Google Scholar] [CrossRef]
- Kim, M.-K.; Lee, J.-S. Short-Term Plasticity and Long-Term Potentiation in Artificial Biosynapses with Diffusive Dynamics. ACS Nano 2018, 12, 1680–1687. [Google Scholar] [CrossRef]
- Ge, J.; Li, D.; Huang, C.; Zhao, X.; Qin, J.; Liu, H.; Ye, W.; Xu, W.; Liu, Z.; Pan, S. Memristive synapses with high reproducibility for flexible neuromorphic networks based on biological nanocomposites. Nanoscale 2019, 12, 720–730. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, X.; Wang, S.; Zhang, C.; Sun, H.; Xu, F.; Tong, Y.; Tang, Q.; Liu, Y. Exploration of the proton conduction behavior in natural neutral polysaccharides for biodegradable organic synaptic transistors. J. Mater. Chem. C 2020, 8, 16542–16550. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, X.; Zhang, C.; Tong, Y.; Hu, J.; Zhang, H.; Yang, M.; Ye, X.; Wang, S.; Sun, Z.; et al. Ultraflexible, Degradable Organic Synaptic Transistors Based on Natural Polysaccharides for Neuromorphic Applications. Adv. Funct. Mater. 2020, 30, 2006271. [Google Scholar] [CrossRef]
- Hu, W.; Jiang, J.; Xie, D.; Wang, S.; Bi, K.; Duan, H.; Yang, J.; He, J. Transient security transistors self-supported on biodegradable natural-polymer membranes for brain-inspired neuromorphic applications. Nanoscale 2018, 10, 14893–14901. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Wang, Y.; Zhang, J.; Zhao, Y.; Xiao, F.; Liu, D.; Wang, T.; Huang, J. Wood-Derived Nanopaper Dielectrics for Organic Synaptic Transistors. ACS Appl. Mater. Interfaces 2018, 10, 39983–39991. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, E.; Yan, Y.; Yang, Q.; Cao, S.; Zhong, J.; Chen, H.; Guo, T. Flexible metal oxide synaptic transistors using biomass-based hydrogel as gate dielectric. J. Phys. D Appl. Phys. 2019, 52, 484002. [Google Scholar] [CrossRef]
- Kim, H.-S.; Park, H.; Cho, W.-J. Biocompatible Casein Electrolyte-Based Electric-Double-Layer for Artificial Synaptic Transistors. Nanomaterials 2022, 12, 2596. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Zhu, L.Q.; Qi, H.F.; Ren, Z.Y.; Wang, F.; Xiao, H. Brain-inspired biodegradable pectin based proton conductor gated electronic synapse. Org. Electron. 2020, 82, 105782. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, C.; Zhao, X.; Tong, Y.; Tang, Q.; Liu, Y. Ultrasensitive and Degradable Ultraflexible Synaptic Transistors Based on Natural Pectin. ACS Appl. Electron. Mater. 2021, 4, 316–325. [Google Scholar] [CrossRef]
- Yang, B.; Wang, Y.; Hua, Z.; Zhang, J.; Li, L.; Hao, D.; Guo, P.; Xiong, L.; Huang, J. Low-power consumption light-stimulated synaptic transistors based on natural carotene and organic semiconductors. Chem. Commun. 2021, 57, 8300–8303. [Google Scholar] [CrossRef]
- Lubrano, C.; Bruno, U.; Ausilio, C.; Santoro, F. Supported Lipid Bilayers Coupled to Organic Neuromorphic Devices Modulate Short-Term Plasticity in Biomimetic Synapses. Adv. Mater. 2022, 34, 2110194. [Google Scholar] [CrossRef]
- Zhang, C.; Xu, F.; Zhao, X.; Zhang, M.; Han, W.; Yu, H.; Wang, S.; Yang, Y.; Tong, Y.; Tang, Q.; et al. Natural polyelectrolyte-based ultraflexible photoelectric synaptic transistors for hemispherical high-sensitive neuromorphic imaging system. Nano Energy 2022, 95, 107001. [Google Scholar] [CrossRef]
- Sung, S.; Park, J.H.; Wu, C.; Kim, T.W. Biosynaptic devices based on chicken egg albumen:graphene quantum dot nanocomposites. Sci. Rep. 2020, 10, 1255. [Google Scholar] [CrossRef]
- Yan, X.; Li, X.; Zhou, Z.; Zhao, J.; Wang, H.; Wang, J.; Zhang, L.; Ren, D.; Zhang, X.; Chen, J.; et al. Flexible Transparent Organic Artificial Synapse Based on the Tungsten/Egg Albumen/Indium Tin Oxide/Polyethylene Terephthalate Memristor. ACS Appl. Mater. Interfaces 2019, 11, 18654–18661. [Google Scholar] [CrossRef] [PubMed]
- Irimia-Vladu, M. “Green” electronics: Biodegradable and biocompatible materials and devices for sustainable future. Chem. Soc. Rev. 2013, 43, 588–610. [Google Scholar] [CrossRef]
- Wu, G.; Feng, P.; Wan, X.; Zhu, L.; Shi, Y.; Wan, Q. Artificial Synaptic Devices Based on Natural Chicken Albumen Coupled Electric-Double-Layer Transistors. Sci. Rep. 2016, 6, 23578. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-K.; Park, Y.; Kim, I.-J.; Lee, J.-S. Emerging Materials for Neuromorphic Devices and Systems. Iscience 2020, 23, 101846. [Google Scholar] [CrossRef]
- Motanis, H.; Seay, M.J.; Buonomano, D.V. Short-Term Synaptic Plasticity as a Mechanism for Sensory Timing. Trends Neurosci. 2018, 41, 701–711. [Google Scholar] [CrossRef]
- Citri, A.; Malenka, R.C. Synaptic Plasticity: Multiple Forms, Functions, and Mechanisms. Neuropsychopharmacology 2008, 33, 18–41. [Google Scholar] [CrossRef] [PubMed]
- Lisman, J. Glutamatergic synapses are structurally and biochemically complex because of multiple plasticity processes: Long-term potentiation, long-term depression, short-term potentiation and scaling. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160260. [Google Scholar] [CrossRef]
- Vitureira, N.; Goda, Y. The interplay between Hebbian and homeostatic synaptic plasticity. J. Cell Biol. 2013, 203, 175–186. [Google Scholar] [CrossRef]
- Ren, Z.Y.; Zhu, L.Q.; Guo, Y.B.; Long, T.Y.; Yu, F.; Xiao, H.; Lu, H.L. Threshold-Tunable, Spike-Rate-Dependent Plasticity Originating from Interfacial Proton Gating for Pattern Learning and Memory. ACS Appl. Mater. Interfaces 2020, 12, 7833–7839. [Google Scholar] [CrossRef]
- Bi, G.-Q.; Poo, M.-M.; Caporale, N.; Dan, Y.; Martin, S.J.; Grimwood, P.D.; Morris, R.G.M.; Hawkins, R.D.; Kandel, E.R.; Siegelbaum, S.A.; et al. Synaptic Modification by Correlated Activity: Hebb’s Postulate Revisited. Annu. Rev. Neurosci. 2001, 24, 139–166. [Google Scholar] [CrossRef]
- Sjöström, P.J.; Rancz, E.A.; Roth, A.; Häusser, M. Dendritic Excitability and Synaptic Plasticity. Physiol. Rev. 2008, 88, 769–840. [Google Scholar] [CrossRef]
- Panwar, N.; Rajendran, B.; Ganguly, U. Arbitrary Spike Time Dependent Plasticity (STDP) in Memristor by Analog Waveform Engineering. IEEE Electron Device Lett. 2017, 38, 740–743. [Google Scholar] [CrossRef]
- Hebb, D.O. The Organization of Behavior: A Neuropsychological Theory; Wiley: New York, NY, USA, 1949. [Google Scholar]
- Hussain, T.; Abbas, H.; Youn, C.; Lee, H.; Boynazarov, T.; Ku, B.; Jeon, Y.; Han, H.; Lee, J.H.; Choi, C.; et al. Cellulose Nanocrystal Based Bio-Memristor as a Green Artificial Synaptic Device for Neuromorphic Computing Applications. Adv. Mater. Technol. 2021, 7, 2100744. [Google Scholar] [CrossRef]
- Shi, C.; Wang, J.; Sushko, M.L.; Qiu, W.; Yan, X.; Liu, X.Y. Silk Flexible Electronics: From Bombyx mori Silk Ag Nanoclusters Hybrid Materials to Mesoscopic Memristors and Synaptic Emulators. Adv. Funct. Mater. 2019, 29, 1904777. [Google Scholar] [CrossRef]
- Qin, W.; Kang, B.H.; Kim, H.J. Flexible Artificial Synapses with a Biocompatible Maltose–Ascorbic Acid Electrolyte Gate for Neuromorphic Computing. ACS Appl. Mater. Interfaces 2021, 13, 34597–34604. [Google Scholar] [CrossRef] [PubMed]
- Ou, Q.; Yang, B.; Zhang, J.; Liu, D.; Chen, T.; Wang, X.; Hao, D.; Lu, Y.; Huang, J. Degradable Photonic Synaptic Transistors Based on Natural Biomaterials and Carbon Nanotubes. Small 2021, 17, e2007241. [Google Scholar] [CrossRef]
- Lamprecht, R.; LeDoux, J. Structural plasticity and memory. Nat. Rev. Neurosci. 2004, 5, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Lan, J.; Wang, J.; Zhang, S.; Lin, Y.; Zhu, S.; Stegmann, A.E.; Yu, R.; Yan, X.; Liu, X.Y. Flexible and Insoluble Artificial Synapses Based on Chemical Cross-Linked Wool Keratin. Adv. Funct. Mater. 2020, 30, 2002882. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, D.; Shi, Q.; Ben Yang, B.; Guo, P.; Fang, L.; Dai, S.; Xiong, L.; Huang, J. Bioinspired organic optoelectronic synaptic transistors based on cellulose nanopaper and natural chlorophyll-a for neuromorphic systems. npj Flex. Electron. 2022, 6, 30. [Google Scholar] [CrossRef]
- López, J.C. A fresh look at paired-pulse facilitation. Nat. Rev. Neurosci. 2001, 2, 307. [Google Scholar] [CrossRef]
- Mazur, T.; Zawal, P.; Szaciłowski, K. Synaptic plasticity, metaplasticity and memory effects in hybrid organic–inorganic bismuth-based materials. Nanoscale 2018, 11, 1080–1090. [Google Scholar] [CrossRef] [PubMed]
- Shouval, H.Z.; Wang, S.S.-H.; Wittenberg, G.M. Spike timing dependent plasticity: A consequence of more fundamental learning rules. Front. Comput. Neurosci. 2010, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Ma, W.; Chang, T.; Sheridan, P.; Lu, W.D. Biorealistic Implementation of Synaptic Functions with Oxide Memristors through Internal Ionic Dynamics. Adv. Funct. Mater. 2015, 25, 4290–4299. [Google Scholar] [CrossRef]
- Kandel, E.R.; Schwartz, J.H. Principles of Neural Science, 2nd ed.; Elsevier: New York, NY, USA, 1985. [Google Scholar]
- Abbott, L.F.; Regehr, W.G. Synaptic computation. Nature 2004, 431, 796–803. [Google Scholar] [CrossRef]
- Ding, G.; Yang, B.; Zhou, K.; Zhang, C.; Wang, Y.; Yang, J.; Han, S.; Zhai, Y.; Roy, V.A.L.; Zhou, Y. Synaptic Plasticity and Filtering Emulated in Metal–Organic Frameworks Nanosheets Based Transistors. Adv. Electron. Mater. 2019, 6, 1900978. [Google Scholar] [CrossRef]
- Yue, K.; Wang, X.; Jadav, J.; Vartak, A.; Parker, A.C. Analog Neurons that Signal with Spiking Frequencies. In Proceedings of the ICONS’19: International Conference on Neuromorphic Systems, Knoxville, TN, USA, 23–25 July 2019; p. 8. [Google Scholar] [CrossRef]
- Segev, I.; Parnas, I. Synaptic integration mechanisms. Theoretical and experimental investigation of temporal postsynaptic interactions between excitatory and inhibitory inputs. Biophys. J. 1983, 41, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Squire, L.R.; Berg, D.; Bloom, F.; Lac, S.; Ghosch, A.; Spitzer, N.C. Fundamental Neuroscience, 4th ed.; Elsevier Inc.: New York, NY, USA, 2014. [Google Scholar]
- Warrant, E.J. Seeing better at night: Life style, eye design and the optimum strategy of spatial and temporal summation. Vis. Res. 1999, 39, 1611–1630. [Google Scholar] [CrossRef]
- Lautenbacher, S.; Kunz, M.; Strate, P.; Nielsen, J.; Arendt-Nielsen, L. Age effects on pain thresholds, temporal summation and spatial summation of heat and pressure pain. Pain 2005, 115, 410–418. [Google Scholar] [CrossRef]
- Paulus, W.; Rothwell, J.C. Membrane resistance and shunting inhibition: Where biophysics meets state-dependent human neurophysiology. J. Physiol. 2016, 594, 2719–2728. [Google Scholar] [CrossRef]
- Huang, W.; Xia, X.; Zhu, C.; Steichen, P.; Quan, W.; Mao, W.; Yang, J.; Chu, L.; Li, X. Memristive Artificial Synapses for Neuromorphic Computing. Nano-Micro Lett. 2021, 13, 1–28. [Google Scholar] [CrossRef]
- Wang, Y.; Yin, L.; Huang, W.; Li, Y.; Huang, S.; Zhu, Y.; Yang, D.; Pi, X. Optoelectronic Synaptic Devices for Neuromorphic Computing. Adv. Intell. Syst. 2020, 3, 2000099. [Google Scholar] [CrossRef]
- Zhuge, X.; Wang, J.; Zhuge, F. Photonic Synapses for Ultrahigh-Speed Neuromorphic Computing. Phys. Status Solidi (RRL) Rapid Res. Lett. 2019, 13, 1900082. [Google Scholar] [CrossRef]
- Hao, D.; Zou, J.; Huang, J. Recent developments in flexible photodetectors based on metal halide perovskite. InfoMat 2019, 2, 139–169. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, Y.; Zhuge, F.; Tian, B.; Yan, M.; Li, Y.; He, Y.; Miao, X.S. Graphene–ferroelectric transistors as complementary synapses for supervised learning in spiking neural network. npj 2D Mater. Appl. 2019, 3, 31. [Google Scholar] [CrossRef]
- Dang, Z.; Guo, F.; Wu, Z.; Jin, K.; Hao, J. Interface Engineering and Device Applications of 2D Ultrathin Film/Ferroelectric Copolymer P(VDF-TrFE). Adv. Phys. Res. 2022, 2, 2200038. [Google Scholar] [CrossRef]
- Luo, Z.-D.; Zhang, S.; Liu, Y.; Zhang, D.; Gan, X.; Seidel, J.; Han, G.; Alexe, M.; Hao, Y. Dual-Ferroelectric-Coupling-Engineered Two-Dimensional Transistors for Multifunctional In-Memory Computing. ACS Nano 2022, 16, 3362–3372. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ref. | Natural Organic Material | Substrate Material | Solution | Film Coating Method | Film Baking | Electrodes | ||
|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | Time (min) | Ambient | ||||||
| [3] | Chitosan | Polyimide | rGO and Chitosan in DI water | Drop-casting | 150 | 90 | Ar | Ti/Au; Ti/Au |
| [4] | Chitosan | Glass | Chitosan and C3N4 in DI water | Drop-casting | RT | _ | Air | IZO; ITO |
| [5] | Chitosan | Glass | rGO and Chitosan in DI water | Spin-coating | RT | _ | Air | IZO; ITO |
| [14] | Zein | Glass | Zein in various solvents | Spin-coating | 50 | 30 | Air | Al; ITO |
| [15] | Gelatin | Si | Gelatin powder in DI water | Spin-coating | RT | 300 | Vacuum | Au; Au |
| [17] | Honey | Glass | Honey in DI water | Spin-coating | 90 | 540 | Air | Ag; ITO |
| [24] | Lignin | PET | Lignin powder in NH4OH and distilled water | Spin-coating | RT | 48h | _ | Au; ITO |
| [25] | Collagen | PET | Collagen powder in DI water | Spin-coating | 60 | 90 | Vacuum | Mg; ITO |
| [26] | Trypsin | Glass | Trypsin powder in Tris-Cl buffer | Drop-casting | RT | 48h | Air | Au; FTO |
| [27] | ι-car | SiO2/Si | ι-car powder in acetic acid and distilled water | Spin-coating | RT | 360 | Air | Ag; Pt |
| [30] | Dextran | Si | Dextran in DI water | Spin-coating | 70 | 60 | Air | Au; Au |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanim, M.M.H.; Templin, Z.; Zhao, F. Natural Organic Materials Based Memristors and Transistors for Artificial Synaptic Devices in Sustainable Neuromorphic Computing Systems. Micromachines 2023, 14, 235. https://doi.org/10.3390/mi14020235
Tanim MMH, Templin Z, Zhao F. Natural Organic Materials Based Memristors and Transistors for Artificial Synaptic Devices in Sustainable Neuromorphic Computing Systems. Micromachines. 2023; 14(2):235. https://doi.org/10.3390/mi14020235
Chicago/Turabian StyleTanim, Md Mehedi Hasan, Zoe Templin, and Feng Zhao. 2023. "Natural Organic Materials Based Memristors and Transistors for Artificial Synaptic Devices in Sustainable Neuromorphic Computing Systems" Micromachines 14, no. 2: 235. https://doi.org/10.3390/mi14020235
APA StyleTanim, M. M. H., Templin, Z., & Zhao, F. (2023). Natural Organic Materials Based Memristors and Transistors for Artificial Synaptic Devices in Sustainable Neuromorphic Computing Systems. Micromachines, 14(2), 235. https://doi.org/10.3390/mi14020235