
A Personalized Glomerulus Chip Engineered from Stem Cell-Derived Epithelium and Vascular Endothelium

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human iPS Cell Culture
2.2. Intermediate Mesoderm Cell Differentiation
2.3. Vascular Endothelial Cell (viEC) Differentiation
2.4. viEC Magnet-Activated Cell Sorting
2.5. Glomerulus Chip Device Functionalization and Cell Seeding
2.6. Personalized Glomerulus Chip Culture Maintenance
2.7. Functional Analysis of the Glomerular Filtration Barrier
2.8. Immunostaining and Microscopy
2.9. Modeling Drug-Induced Nephrotoxicity
3. Results
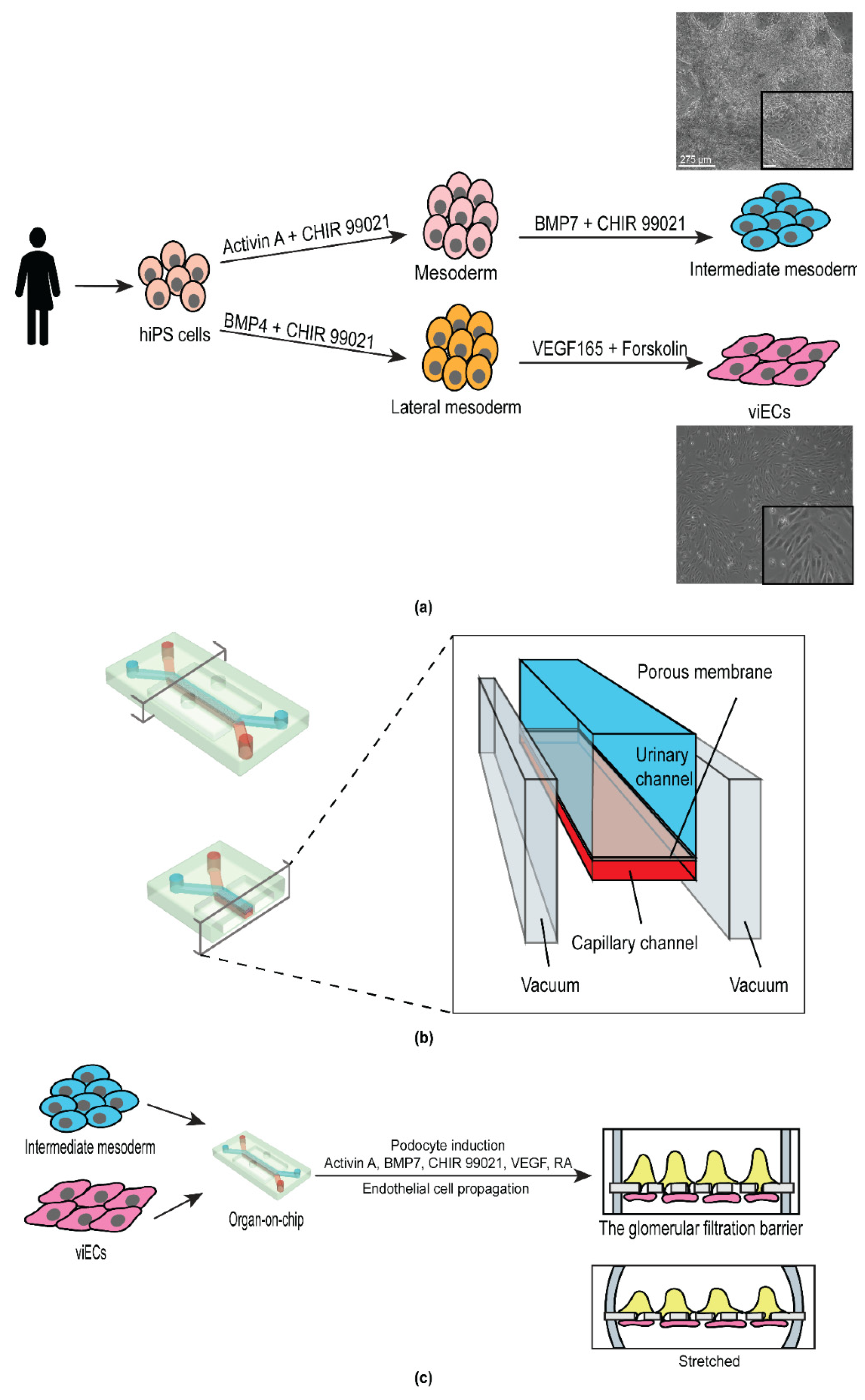
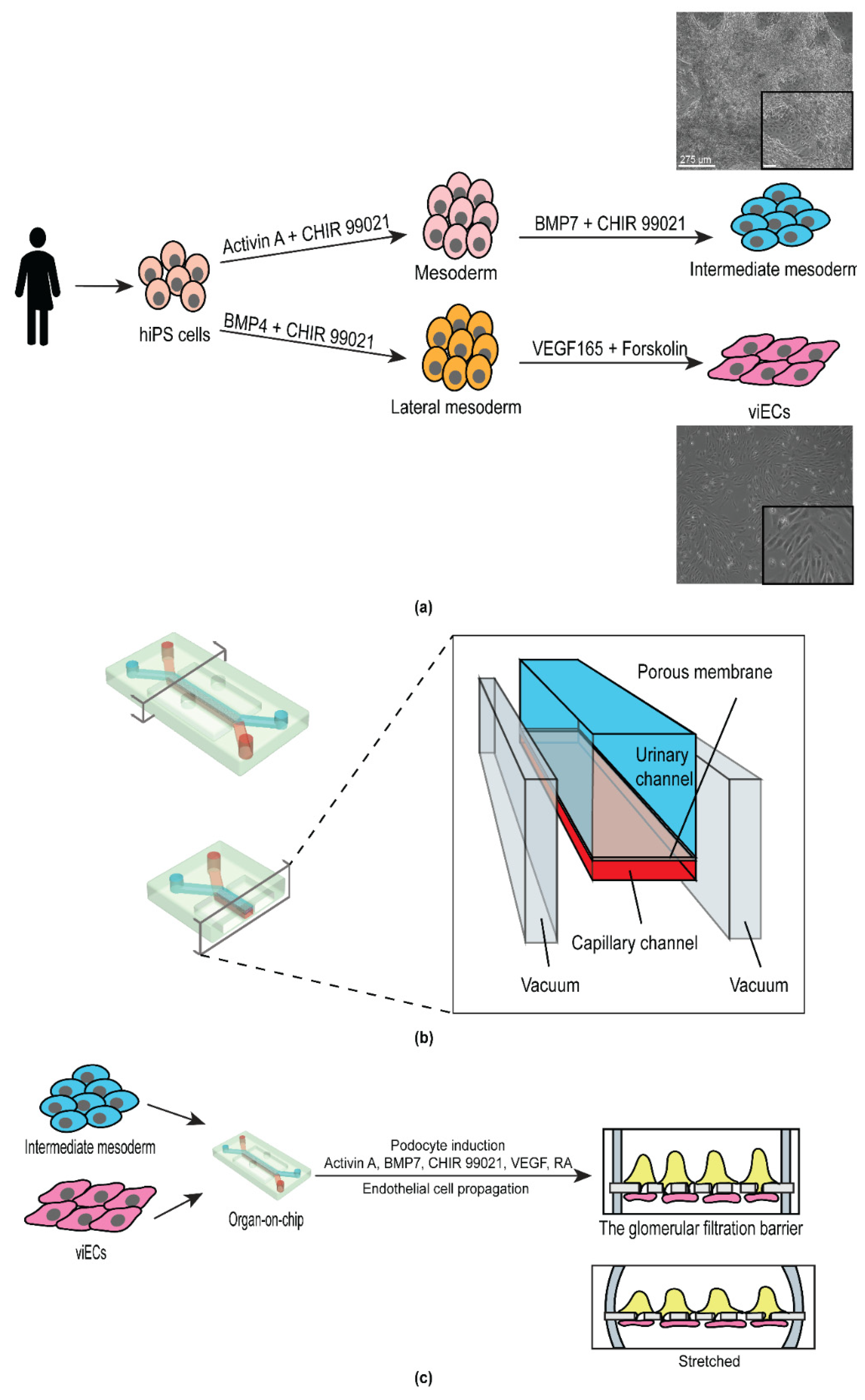
3.1. Derivation of Intermediate Mesoderm and Vascular Endothelial Cells from Human iPS Cells
3.2. Design and Development of the Personalized Glomerulus Chip
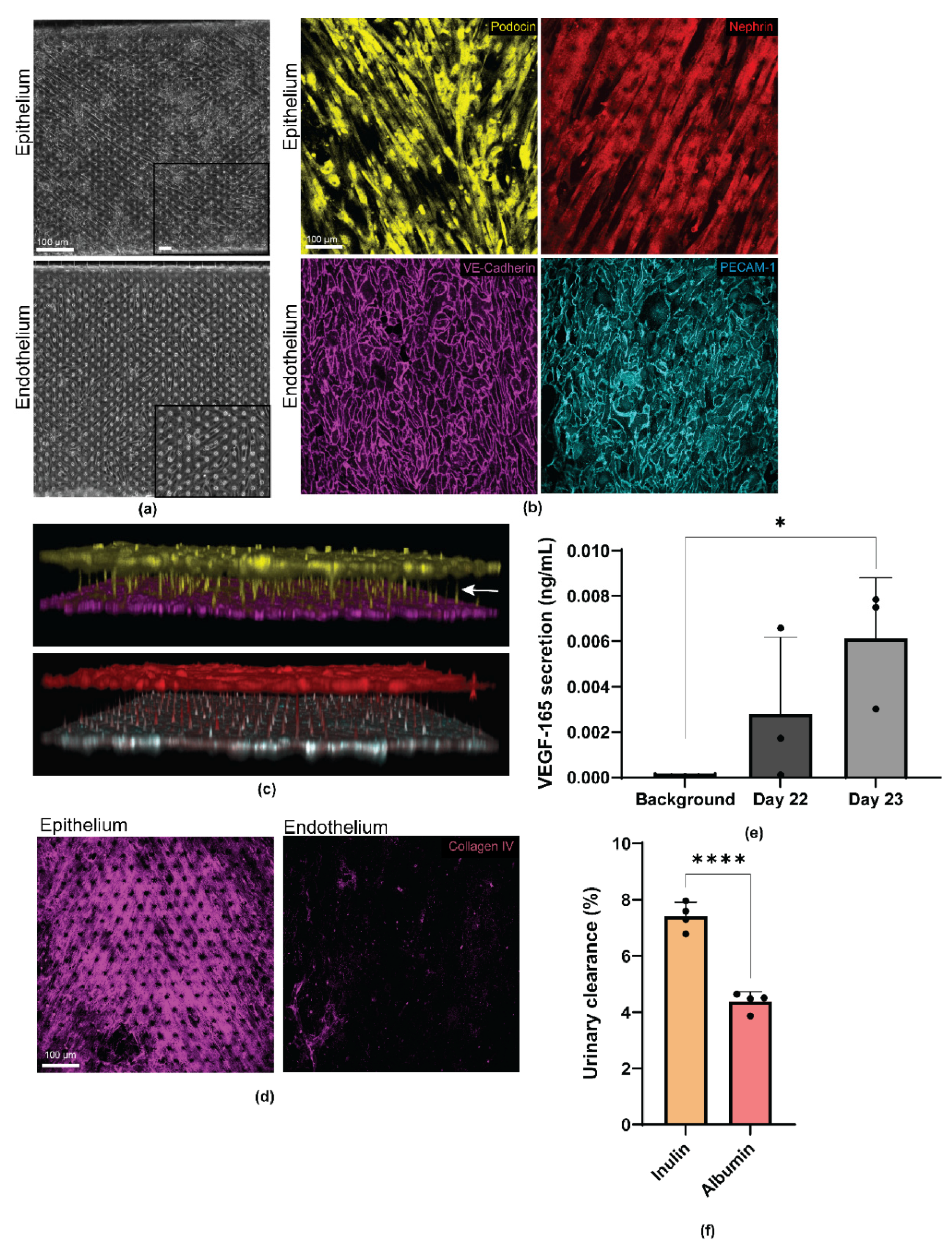
3.3. Differentiated Human iPS Cell-Derived Podocytes and Vascular Endothelium Emulate the Structure of the Kidney Glomerulus
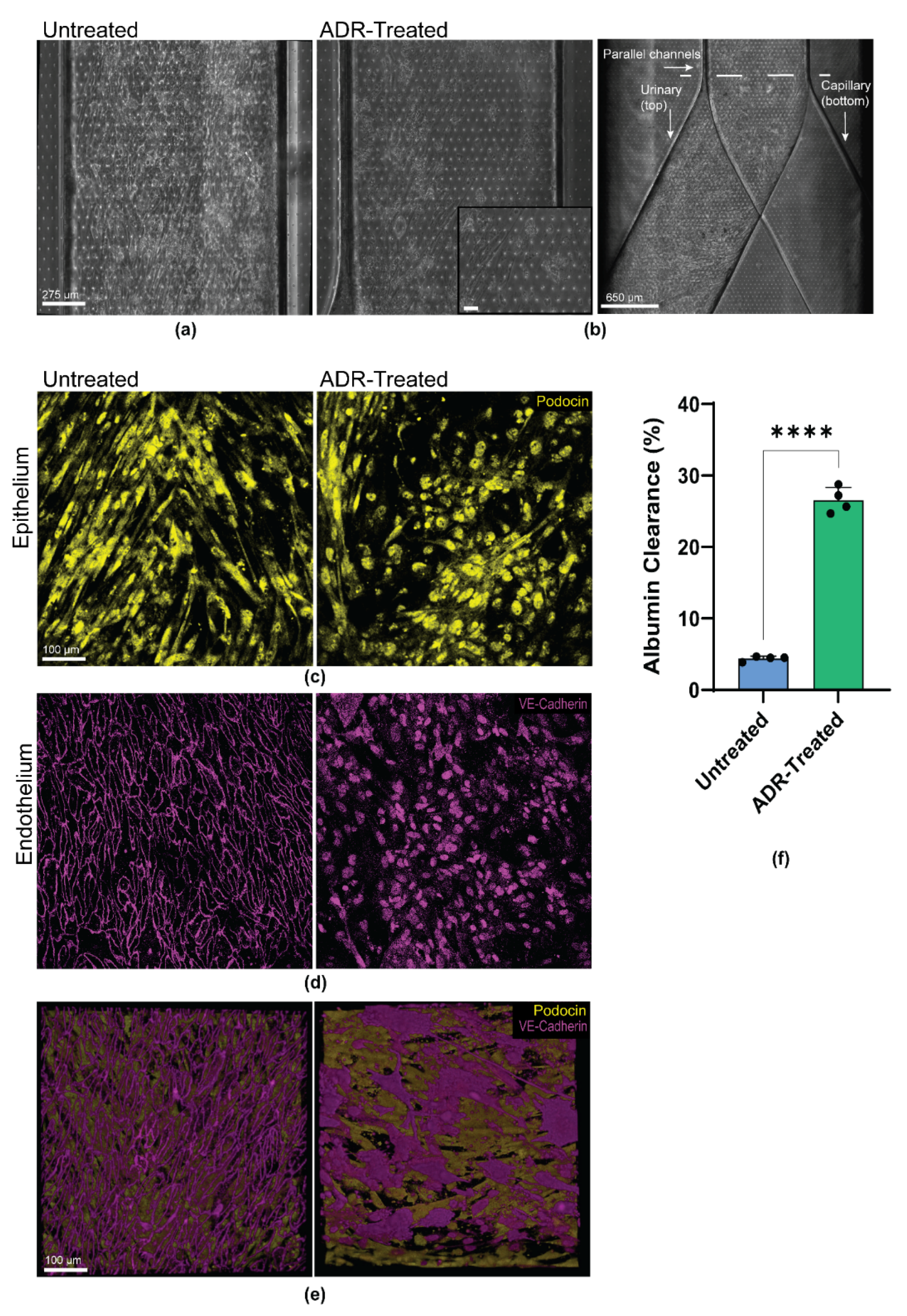
3.4. Personalized Glomerulus Chip Models Drug-Induced Nephropathy
4. Discussion
4.1. The Personalized Glomerulus Chip
4.2. Model Limitations and Outlook
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chronic Kidney Disease in the United States. 2021. Available online: https://www.cdc.gov/kidneydisease/publications-resources/ckd-national-facts.html (accessed on 2 June 2021).
- McCullough, K.P.; Morgenstern, H.; Saran, R.; Herman, W.H.; Robinson, B.M. Projecting ESRD Incidence and Prevalence in the United States through 2030. J. Am. Soc. Nephrol. 2019, 30, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Virani, S.S.; Alonso, A.; Aparicio, H.J.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics—2021 Update. Circulation 2021, 143, e254–e743. [Google Scholar] [CrossRef]
- Gheith, O.; Farouk, N.; Nampoory, N.; Halim, M.A.; Al-Otaibi, T. Diabetic Kidney Disease: World Wide Difference of Prevalence and Risk Factors. J. Nephropharmacol. 2015, 5, 49–56. [Google Scholar] [CrossRef]
- Dąbrowska-Bender, M.; Dykowska, G.; Żuk, W.; Milewska, M.; Staniszewska, A. The Impact on Quality of Life of Dialysis Patients with Renal Insufficiency. Patient Prefer. Adherence 2018, 12, 577–583. [Google Scholar] [CrossRef] [Green Version]
- de Boer, I.H.; Alpers, C.E.; Azeloglu, E.U.; Balis, U.G.J.; Barasch, J.M.; Barisoni, L.; Blank, K.N.; Bomback, A.S.; Brown, K.; Dagher, P.C.; et al. Rationale and Design of the Kidney Precision Medicine Project. Kidney Int. 2021, 99, 498–510. [Google Scholar] [CrossRef]
- Nagata, M. Podocyte Injury and Its Consequences. Kidney Int. 2016, 89, 1221–1230. [Google Scholar] [CrossRef]
- Gil, C.L.; Hooker, E.; Larrivée, B. Diabetic Kidney Disease, Endothelial Damage, and Podocyte-Endothelial Crosstalk. Kidney Med. 2021, 3, 105–115. [Google Scholar] [CrossRef]
- Park, J.; Shrestha, R.; Qiu, C.; Kondo, A.; Huang, S.; Werth, M.; Li, M.; Barasch, J.; Suszták, K. Single-Cell Transcriptomics of the Mouse Kidney Reveals Potential Cellular Targets of Kidney Disease. Science 2018, 360, 758–763. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Malone, A.F.; Donnelly, E.L.; Kirita, Y.; Uchimura, K.; Ramakrishnan, S.M.; Gaut, J.P.; Humphreys, B.D. Single-Cell Transcriptomics of a Human Kidney Allograft Biopsy Specimen Defines a Diverse Inflammatory Response. J. Am. Soc. Nephrol. JASN 2018, 29, 2069–2080. [Google Scholar] [CrossRef]
- Pammolli, F.; Magazzini, L.; Riccaboni, M. The Productivity Crisis in Pharmaceutical R&D. Nat. Rev. Drug Discov. 2011, 10, 428–438. [Google Scholar] [CrossRef]
- Wu, H.; Uchimura, K.; Donnelly, E.L.; Kirita, Y.; Morris, S.A.; Humphreys, B.D. Comparative Analysis and Refinement of Human PSC-Derived Kidney Organoid Differentiation with Single-Cell Transcriptomics. Cell Stem Cell 2018, 23, 869–881.e8. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, R.; Bonner, M.G.; Musah, S. Harnessing Developmental Plasticity to Pattern Kidney Organoids. Cell Stem Cell 2021, 28, 587–589. [Google Scholar] [CrossRef]
- Howden, S.E.; Wilson, S.B.; Groenewegen, E.; Starks, L.; Forbes, T.A.; Tan, K.S.; Vanslambrouck, J.M.; Holloway, E.M.; Chen, Y.-H.; Jain, S.; et al. Plasticity of Distal Nephron Epithelia from Human Kidney Organoids Enables the Induction of Ureteric Tip and Stalk. Cell Stem Cell 2021, 28, 671–684.e6. [Google Scholar] [CrossRef]
- Zhou, M.; Zhang, X.; Wen, X.; Wu, T.; Wang, W.; Yang, M.; Wang, J.; Fang, M.; Lin, B.; Lin, H. Development of a Functional Glomerulus at the Organ Level on a Chip to Mimic Hypertensive Nephropathy. Sci. Rep. 2016, 6, 31771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Du, G.; Zhang, B.; Zhang, H.; Yin, R.; Zhang, W.; Yang, S.-M. Efficient Drug Screening and Nephrotoxicity Assessment on Co-Culture Microfluidic Kidney Chip. Sci. Rep. 2020, 10, 6568. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced Pluripotent Stem Cell Lines Derived from Human Somatic Cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Okita, K.; Ichisaka, T.; Yamanaka, S. Generation of Germline-Competent Induced Pluripotent Stem Cells. Nature 2007, 448, 313–317. [Google Scholar] [CrossRef]
- Musah, S.; Mammoto, A.; Ferrante, T.C.; Jeanty, S.S.F.; Hirano-Kobayashi, M.; Mammoto, T.; Roberts, K.; Chung, S.; Novak, R.; Ingram, M.; et al. Mature Induced-Pluripotent-Stem-Cell-Derived Human Podocytes Reconstitute Kidney Glomerular-Capillary-Wall Function on a Chip. Nat. Biomed. Eng. 2017, 1, 1–12. [Google Scholar] [CrossRef]
- Musah, S.; Dimitrakakis, N.; Camacho, D.M.; Church, G.M.; Ingber, D.E. Directed Differentiation of Human Induced Pluripotent Stem Cells into Mature Kidney Podocytes and Establishment of a Glomerulus Chip. Nat. Protoc. 2018, 13, 1662–1685. [Google Scholar] [CrossRef]
- Kwist, K.; Bridges, W.C.; Burg, K.J.L. The Effect of Cell Passage Number on Osteogenic and Adipogenic Characteristics of D1 Cells. Cytotechnology 2016, 68, 1661–1667. [Google Scholar] [CrossRef]
- Jin, W.; Penington, C.J.; McCue, S.W.; Simpson, M.J. A Computational Modelling Framework to Quantify the Effects of Passaging Cell Lines. PLoS ONE 2017, 12, e0181941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurer, L.L.; Latham, J.D.; Landis, R.W.; Song, D.H.; Epstein, T.; Philbert, M.A. 1,3-Dinitrobenzene Neurotoxicity – Passage Effect in Immortalized Astrocytes. Neurotoxicology 2016, 53, 74–84. [Google Scholar] [CrossRef]
- Zhong, F.; Wang, W.; Lee, K.; He, J.C.; Chen, N. Role of C/EBP-α in Adriamycin-Induced Podocyte Injury. Sci. Rep. 2016, 6, 33520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Tao, T.; Su, W.; Yu, H.; Yu, Y.; Qin, J. A Disease Model of Diabetic Nephropathy in a Glomerulus-on-a-Chip Microdevice. Lab Chip 2017, 17, 1749–1760. [Google Scholar] [CrossRef]
- Petrosyan, A.; Cravedi, P.; Villani, V.; Angeletti, A.; Manrique, J.; Renieri, A.; De Filippo, R.E.; Perin, L.; Da Sacco, S. A Glomerulus-on-a-Chip to Recapitulate the Human Glomerular Filtration Barrier. Nat. Commun. 2019, 10, 3656. [Google Scholar] [CrossRef] [Green Version]
- Abutaleb, N.O.; Truskey, G.A. Human IPSCs Stretch to Improve Tissue-Engineered Vascular Grafts. Cell Stem Cell 2020, 26, 136–137. [Google Scholar] [CrossRef] [PubMed]
- Atchison, L.; Abutaleb, N.O.; Snyder-Mounts, E.; Gete, Y.; Ladha, A.; Ribar, T.; Cao, K.; Truskey, G.A. IPSC-Derived Endothelial Cells Affect Vascular Function in a Tissue-Engineered Blood Vessel Model of Hutchinson-Gilford Progeria Syndrome. Stem Cell Rep. 2020, 14, 325–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patsch, C.; Challet-Meylan, L.; Thoma, E.C.; Urich, E.; Heckel, T.; O’Sullivan, J.F.; Grainger, S.J.; Kapp, F.G.; Sun, L.; Christensen, K.; et al. Generation of Vascular Endothelial and Smooth Muscle Cells from Human Pluripotent Stem Cells. Nat. Cell Biol. 2015, 17, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Abrahamson, D.R. Role of the Podocyte (and Glomerular Endothelium) in Building the GBM. Semin. Nephrol. 2012, 32, 342–349. [Google Scholar] [CrossRef] [Green Version]
- Byron, A.; Randles, M.J.; Humphries, J.D.; Mironov, A.; Hamidi, H.; Harris, S.; Mathieson, P.W.; Saleem, M.A.; Satchell, S.C.; Zent, R.; et al. Glomerular Cell Cross-Talk Influences Composition and Assembly of Extracellular Matrix. J. Am. Soc. Nephrol. 2014, 25, 953–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobeika, L.; Barati, M.T.; Caster, D.J.; McLeish, K.R.; Merchant, M.L. Characterization of Glomerular Extracellular Matrix by Proteomic Analysis of Laser-Captured Microdissected Glomeruli. Kidney Int. 2017, 91, 501–511. [Google Scholar] [CrossRef] [Green Version]
- Suh, J.H.; Miner, J.H. The Glomerular Basement Membrane as a Barrier to Albumin. Nat. Rev. Nephrol. 2013, 9, 470–477. [Google Scholar] [CrossRef] [Green Version]
- Sand, J.M.; Larsen, L.; Hogaboam, C.; Martinez, F.; Han, M.; Røssel Larsen, M.; Nawrocki, A.; Zheng, Q.; Asser Karsdal, M.; Leeming, D.J. MMP Mediated Degradation of Type IV Collagen Alpha 1 and Alpha 3 Chains Reflects Basement Membrane Remodeling in Experimental and Clinical Fibrosis—Validation of Two Novel Biomarker Assays. PLoS ONE 2013, 8, e84934. [Google Scholar] [CrossRef]
- Liu, P.; Xie, X.; Jin, J. Isotopic Nitrogen-15 Labeling of Mice Identified Long-Lived Proteins of the Renal Basement Membranes. Sci. Rep. 2020, 10, 5317. [Google Scholar] [CrossRef] [Green Version]
- Tufró, A. VEGF Spatially Directs Angiogenesis during Metanephric Development In Vitro. Dev. Biol. 2000, 227, 558–566. [Google Scholar] [CrossRef] [Green Version]
- Glass, N.R.; Takasako, M.; Er, P.X.; Titmarsh, D.M.; Hidalgo, A.; Wolvetang, E.J.; Little, M.H.; Cooper-White, J.J. Multivariate Patterning of Human Pluripotent Cells under Perfusion Reveals Critical Roles of Induced Paracrine Factors in Kidney Organoid Development. Sci. Adv. 2020, 6, eaaw2746. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.C.; Muthukrishnan, S.D.; Oxburgh, L. A Synthetic Niche for Nephron Progenitor Cells. Dev. Cell 2015, 34, 229–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, M.; Zhang, L.; Liu, Y.; Livingston, M.J.; Chen, J.-K.; Nahman, N.S.; Liu, F.; Dong, Z. Autophagy Is Activated to Protect against Podocyte Injury in Adriamycin-Induced Nephropathy. Am. J. Physiol.-Ren. Physiol. 2017, 313, F74–F84. [Google Scholar] [CrossRef] [Green Version]
- Maschmeyer, I.; Lorenz, A.K.; Schimek, K.; Hasenberg, T.; Ramme, A.P.; Hübner, J.; Lindner, M.; Drewell, C.; Bauer, S.; Thomas, A.; et al. A Four-Organ-Chip for Interconnected Long-Term Co-Culture of Human Intestine, Liver, Skin and Kidney Equivalents. Lab Chip 2015, 15, 2688–2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schetz, M.; Dasta, J.; Goldstein, S.; Golper, T. Drug-Induced Acute Kidney Injury. Curr. Opin. Crit. Care 2005, 11, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Cardiorenal Syndrome: Classification, Pathophysiology, Diagnosis, and Treatment Strategies: A Scientific Statement from the American Heart Association | Circulation. Available online: https://www.ahajournals.org/doi/full/10.1161/CIR.0000000000000664 (accessed on 2 June 2021).
- Qu, Y.; An, F.; Luo, Y.; Lu, Y.; Liu, T.; Zhao, W.; Lin, B. A Nephron Model for Study of Drug-Induced Acute Kidney Injury and Assessment of Drug-Induced Nephrotoxicity. Biomaterials 2018, 155, 41–53. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roye, Y.; Bhattacharya, R.; Mou, X.; Zhou, Y.; Burt, M.A.; Musah, S. A Personalized Glomerulus Chip Engineered from Stem Cell-Derived Epithelium and Vascular Endothelium. Micromachines 2021, 12, 967. https://doi.org/10.3390/mi12080967
Roye Y, Bhattacharya R, Mou X, Zhou Y, Burt MA, Musah S. A Personalized Glomerulus Chip Engineered from Stem Cell-Derived Epithelium and Vascular Endothelium. Micromachines. 2021; 12(8):967. https://doi.org/10.3390/mi12080967
Chicago/Turabian StyleRoye, Yasmin, Rohan Bhattacharya, Xingrui Mou, Yuhao Zhou, Morgan A. Burt, and Samira Musah. 2021. "A Personalized Glomerulus Chip Engineered from Stem Cell-Derived Epithelium and Vascular Endothelium" Micromachines 12, no. 8: 967. https://doi.org/10.3390/mi12080967
APA StyleRoye, Y., Bhattacharya, R., Mou, X., Zhou, Y., Burt, M. A., & Musah, S. (2021). A Personalized Glomerulus Chip Engineered from Stem Cell-Derived Epithelium and Vascular Endothelium. Micromachines, 12(8), 967. https://doi.org/10.3390/mi12080967