Characterization of Single-Nucleus Electrical Properties by Microfluidic Constriction Channel




Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
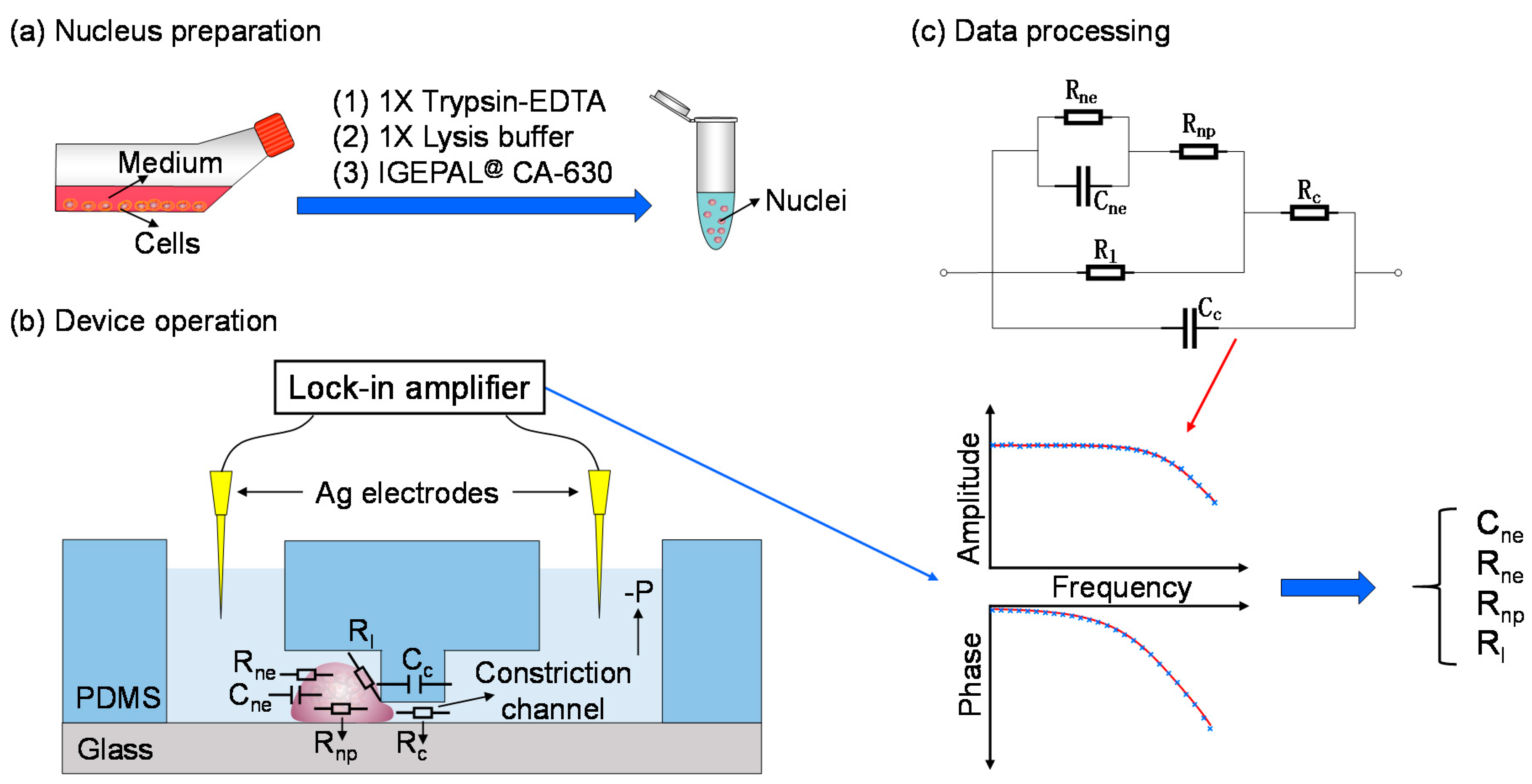
2.2. Nucleus Preparation
2.3. Device Fabrication
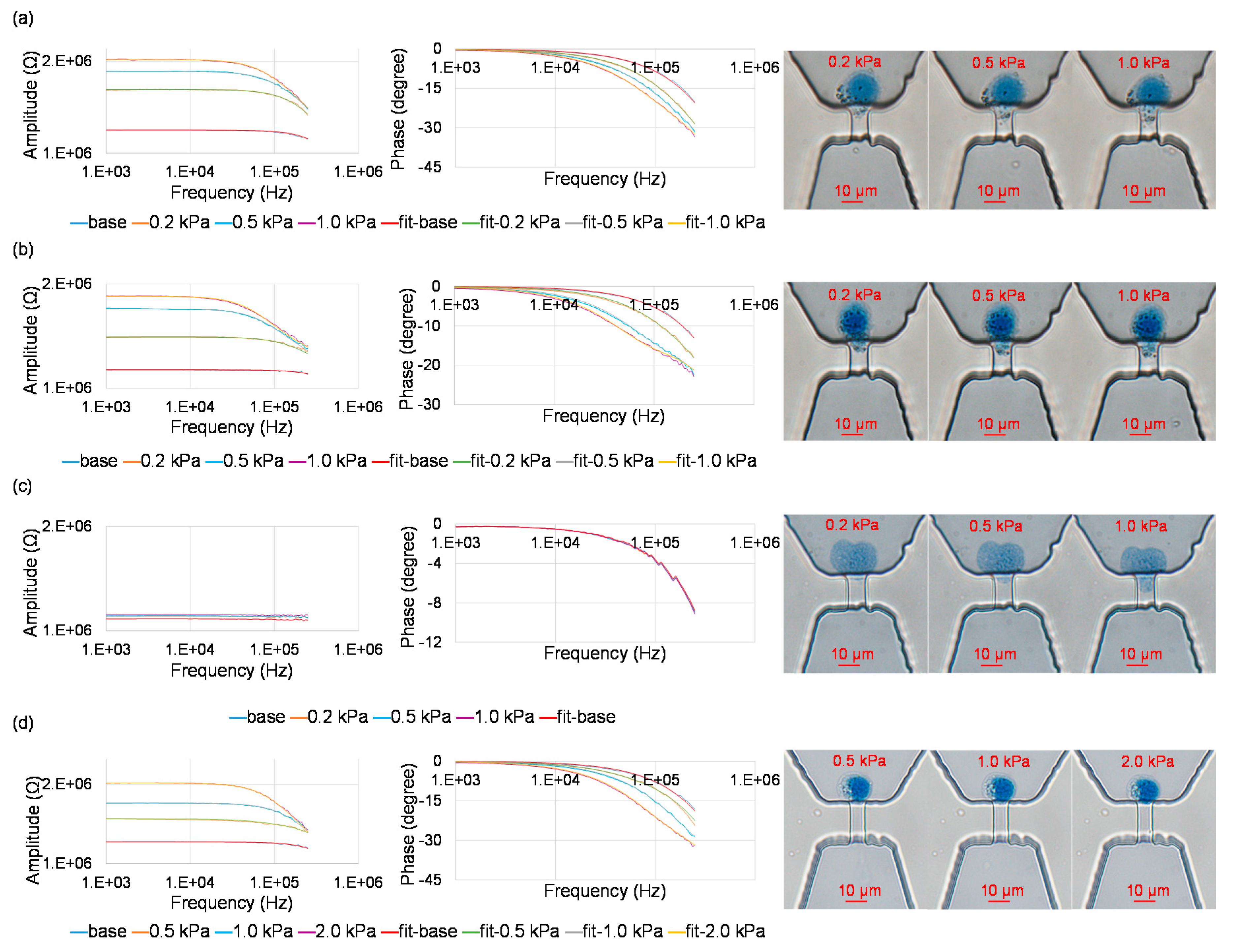
2.4. Device Operation
2.5. Data Processing
2.6. Statistics
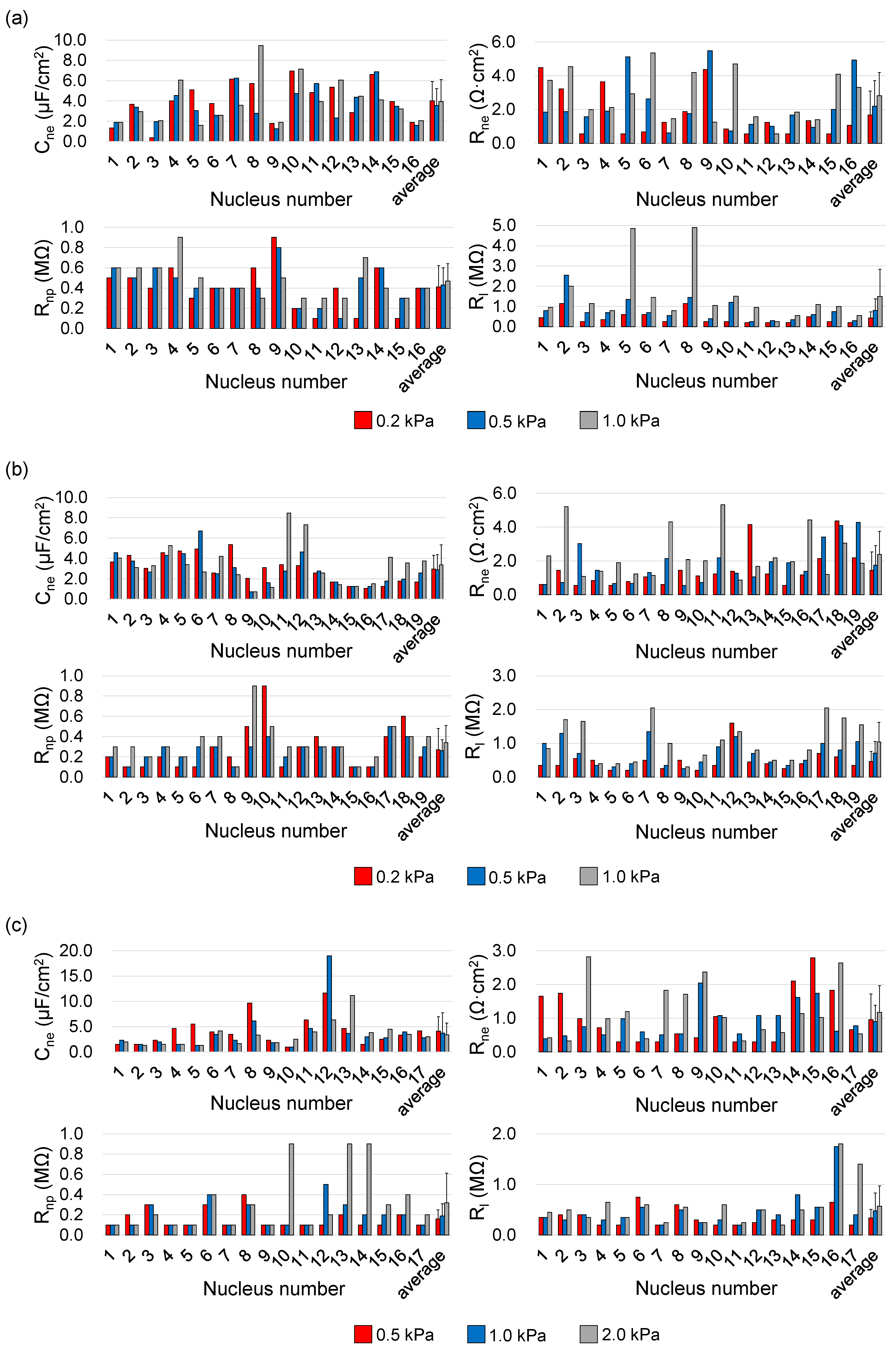
3. Results and Discussion
4. Conclusions and Future Developments
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Loewenstein, W.R.; Kanno, Y. Some electrical properties of the membrane of a cell nucleus. Nature 1962, 195, 462–464. [Google Scholar] [CrossRef]
- Loewenstein, W.R.; Kanno, Y. Some electrical properties of a nuclear membrane examined with a microelectrode. J. Gen Physiol. 1963, 46, 1123–1140. [Google Scholar] [CrossRef] [PubMed]
- Loewenstein, W.R.; Kanno, Y. The electrical conductance and potential across the membrane of some cell nuclei. J. Cell Biol. 1963, 16, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Loewenstein, W.R. Permeability of a nuclear membrane: changes during normal development and changes induced by growth hormone. Science 1965, 150, 909–910. [Google Scholar] [PubMed]
- Loewenstein, W.R.; Kanno, Y.; Ito, S. Permeability of nuclear membranes. Ann. N. Y. Acad. Sci. 1966, 137, 708–716. [Google Scholar] [CrossRef]
- Palmer, L.G.; Civan, M.M. Distribution of Na +, K + and C1- between nucleus and cytoplasm in chironomus salivary gland cells. J. Membr. Biol. 1977, 33, 41–61. [Google Scholar] [CrossRef]
- Kanno, Y.; Loewenstein, W.R. A study of the nucleus and cell membranes of oocytes with an intra-cellular electrode. Exp. Cell Res. 1963, 31, 149–166. [Google Scholar] [CrossRef]
- Kanno, Y.; Ashman, R.F.; Loewenstein, W.R. Nucleus and cell membrane conductance in marine oocytes. Exp. Cell Res. 1965, 39, 184–189. [Google Scholar] [CrossRef]
- Dale, B. Voltage clamp of the nuclear envelope. Proc. R. Soc. B 1994, 255, 119–124. [Google Scholar]
- Danker, T. Using atomic force microscopy to investigate patch-clamped nuclear membrane. Cell Biol. Int. 1997, 21, 747–757. [Google Scholar] [CrossRef]
- Mazzanti, M. Ion permeability of the nuclear envelope. News Physiol. Sci. 1998, 13, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Danker, T. Nuclear hourglass technique: An approach that detects electrically open nuclear pores in xenopus laevis oocyte. Proc. Natl. Acad. Sci. USA 1999, 96, 13530–13535. [Google Scholar] [CrossRef] [PubMed]
- Danker, T. Electrophoretic plugging of nuclear pores by using the nuclear hourglass technique. J. Membr. Biol. 2001, 184, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Ziervogel, H. Electrorotation of lymphocytes--the influence of membrane events and nucleus. Biosci. Rep. 1986, 6, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Asami, K.; Takahashi, Y.; Takashima, S. Dielectric properties of mouse lymphocytes and erythrocytes. Biochim. Biophys. Acta 1989, 1010, 49–55. [Google Scholar] [CrossRef]
- Griffith, A.W.; Cooper, J.M. Single-cell measurements of human neutrophil activation using electrorotation. Anal. Chem. 1998, 70, 2607–2612. [Google Scholar] [CrossRef]
- Polevaya, Y. Time domain dielectric spectroscopy study of human cells. II. normal and malignant white blood cells. Biochim. Biophys. Acta 1999, 1419, 257–271. [Google Scholar] [CrossRef]
- Ermolina, I.; Polevaya, Y.; Feldman, Y. Study of normal and malignant white blood cells by time domain dielectric spectroscopy. IEEE Trans. Dielectr. Electr. Insul. 2001, 8, 253–261. [Google Scholar] [CrossRef]
- Egger, M.; Donath, E. Electrorotation measurements of diamide-induced platelet activation changes. Biophys. J. 1995, 68, 364. [Google Scholar] [CrossRef]
- Feldman, Y.; Ermolina, I.; Hayashi, Y. Time domain dielectric spectroscopy study of biological systems. IEEE Trans. Dielectr. Electr. Insul. 2003, 10, 728–753. [Google Scholar] [CrossRef]
- Raicu, V.; Raicu, G.; Turcu, G. Dielectric properties of yeast cells as simulated by the two-shell model. Biochim. Biophys. Acta 1996, 1274, 143–148. [Google Scholar] [CrossRef]
- Matzke, A.J.; Weiger, T.M.; Matzke, M. Ion channels at the nucleus: electrophysiology meets the genome. Mol. Plant 2010, 3, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Giulian, D.; Diacumakos, E.G. Electrophysiological mapping of compartments within a mammalian-cell. J. Cell Biol. 1977, 72, 86–103. [Google Scholar] [CrossRef] [PubMed]
- Irimajiri, A. Passive electrical properties of cultured murine lymphoblast (L 5178 Y) with reference to its cytoplasmic membrane, nuclear envelope, and intracellular phases. J. Membr. Biol. 1978, 38, 209–232. [Google Scholar] [CrossRef] [PubMed]
- Gimsa, J. Dielectrophoresis and electrorotation of neurospora slime and murine myeloma cells. Biophys. J. 1991, 60, 749–760. [Google Scholar] [CrossRef][Green Version]
- Oberleithner, H. Imaging nuclear pores of aldosterone-sensitive kidney cells by atomic force microscopy. Proc. Natl. Acad. Sci. USA 1994, 91, 9784–9788. [Google Scholar] [CrossRef]
- Garner, A.L. Time domain dielectric spectroscopy measurements of HL-60 cell suspensions after microsecond and nanosecond electrical pulses. IEEE Trans. Plasma Sci. 2004, 32, 2073–2084. [Google Scholar] [CrossRef]
- Garner, A.L. Ultrashort electric pulse induced changes in cellular dielectric properties. Biochem. Biophys. Res. Commun. 2007, 362, 139–144. [Google Scholar] [CrossRef][Green Version]
- Massimi, M. Dielectric Characterization of hepatocytes in suspension and embedded into two different polymeric scaffolds. Colloids Surf. B Biointerfaces 2013, 102, 700–707. [Google Scholar] [CrossRef]
- Moisescu, M.G. Changes of cell electrical parameters induced by electroporation. A dielectrophoresis study. Biochim. Biophys. Acta 2013, 1828, 365–372. [Google Scholar] [CrossRef]
- Zhuang, J. Pulsed electric field induced dielectric evolution of mammalian cells. In Proceedings of the IEEE International Power Modulator and High Voltage Conference, San Diego, CA, USA, 3–7 June 2013. [Google Scholar]
- Anderson, S.E.; Bau, H.H. Electrical detection of cellular penetration during microinjection with carbon nanopipettes. Nanotechnology 2014, 25, 245102. [Google Scholar] [CrossRef]
- Sano, M.B. In-vitro bipolar nano- and microsecond electro-pulse bursts for irreversible electroporation therapies. Bioelectrochemistry 2014, 100, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Stacey, M.W.; Sabuncu, A.C.; Beskok, A. Dielectric characterization of costal cartilage chondrocytes. Biochim. Biophys. Acta 2014, 1840, 146–152. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Murovec, T. Modeling of transmembrane potential in realistic multicellular structures before electroporation. Bioelectrochem. Bioenerg. 2016, 111, 2286–2295. [Google Scholar] [CrossRef] [PubMed]
- Sabuncu, A.C. Microfluidic impedance spectroscopy as a tool for quantitative biology and biotechnology. Biomicrofluidics 2012, 6, 137–152. [Google Scholar] [CrossRef]
- Zhou, Y. Single cell studies of mouse embryonic stem cell (mESC) differentiation by electrical impedance measurements in a microfluidic device. Biosens. Bioelectron. 2016, 81, 249–258. [Google Scholar] [CrossRef]
- Reynolds, C.R.; Tedeschi, H. Permeability properties of mammalian cell nuclei in living cells and in vitro. J. Cell Sci. 1984, 70, 197–207. [Google Scholar]
- Mazzanti, M.; DeFeliceI, L.J. Ion channel in the nuclear envelope. Nature 1990, 343, 764–767. [Google Scholar] [CrossRef]
- Matzke, A.J.M. A large conductance ion channel in the nuclear envelope of higher plant cell. FEBS Lett. 1992, 302, 81–85. [Google Scholar] [CrossRef]
- Raicu, V. Dielectric properties of rat liver in vivo: Analysis by modeling hepatocytes in the tissue architecture. Bioelectrochem. Bioenerg. 1998, 47, 333–342. [Google Scholar] [CrossRef]
- Vanderstraeten, J.; Vander Vorst, A. Theoretical evaluation of dielectric absorption of microwave energy at the scale of nucleic acids. Bioelectromagnetics 2004, 25, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Bai, W.; Zhao, K.S.; Asami, K. Dielectric properties of E. coli cell as simulated by the three-shell spheroidal model. Bioelectrochem. Bioenerg. 2006, 122, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Gerber, H.L.; Bassi, A.; Tseng, C.C. Determining a pulse coupling to subcellular components with a frequency-domain transfer function. IEEE Trans. Plasma Sci. 2006, 34, 1425–1430. [Google Scholar] [CrossRef]
- Kotnik, T.; Miklavc, D. Theoretical evaluation of voltage inducement on internal membranes of biological cells exposed to electric fields. Biophys. J. 2006, 90, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Shibatani, S. Electrorotation of vacuoles isolated from barley mesophyll cells. Bioelectrochem. Bioenerg. 1993, 29, 327–335. [Google Scholar] [CrossRef]
- Chun-Chieh, C.; Ke, W.; Yi, Z.; Deyong, C.; Beiyuan, F.; Chia-Hsun, H. Mechanical property characterization of hundreds of single nuclei based on microfluidic constriction channel. Cytom. A 2018, 93, 1–7. [Google Scholar]
- Gravesen, P.; Branebjerg, J.; Jensen, O.S. Microfluidics-a review. J. Micromech. Microeng. 1993, 3, 168. [Google Scholar] [CrossRef]
- Gross, A.; Schoendube, J.; Zimmermann, S.; Steeb, M.; Zengerle, R.; Koltay, P. Technologies for single-cell isolation. Int. J. Mol. Sci. 2015, 16, 16897–16919. [Google Scholar] [CrossRef]
- Shields, C.W., IV; Reyes, C.D.; López, G.P. Microfluidic cell sorting: a review of the advances in the separation of cells from debulking to rare cell isolation. Lab Chip 2015, 15, 1230–1249. [Google Scholar]
- Warkiani, M.E.; Wu, L.; Tay, A.K.P.; Han, J. Large-volume microfluidic cell sorting for biomedical applications. Annu. Rev. Biomed. Eng. 2015, 17, 1. [Google Scholar] [CrossRef]
- Mu, X.; Zheng, W.; Sun, J.; Zhang, W.; Jiang, X. Microfluidics for manipulating cells. Small 2013, 9, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Ayliffe, H.E.; Frazier, A.B.; Rabbitt, R.D. Electric impedance spectroscopy using microchannels with integrated metal electrodes. IEEE J. Microelectromech. Syst. 1999, 8, 50–57. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nucleus Type | Constriction Channel Dimensions | IGEPAL@ CA-630 | Aspiration Pressure (kPa) | Cne (μF/cm2) | Rne (Ω·cm2) | Rnp (MΩ) | Nucleus Number |
|---|---|---|---|---|---|---|---|
| A549 | 7 μm × 8 μm | 0.01% | 0.2 | 4.02 ± 1.91 | 1.67 ± 1.37 | 0.41 ± 0.21 | N = 16 |
| 0.5 | 3.55 ± 1.66 | 2.20 ± 1.52 | 0.43 ± 0.17 | ||||
| 1.0 | 3.94 ± 2.16 | 2.82 ± 1.43 | 0.47 ± 0.17 | ||||
| 0.02% | 0.2 | 2.97 ± 1.32 | 1.45 ± 1.07 | 0.27 ± 0.21 | N = 19 | ||
| 0.5 | 2.90 ± 1.47 | 1.76 ± 1.14 | 0.26 ± 0.11 | ||||
| 1.0 | 3.38 ± 1.96 | 2.38 ± 1.37 | 0.34 ± 0.17 | ||||
| SW620 | 5 μm × 5 μm | 0.01% | 0.5 | 4.13 ± 2.82 | 0.96 ± 0.76 | 0.16 ± 0.09 | N = 17 |
| 1.0 | 3.74 ± 4.03 | 0.90 ± 0.48 | 0.19 ± 0.12 | ||||
| 2.0 | 3.38 ± 2.37 | 1.18 ± 0.79 | 0.32 ± 0.29 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, H.; Zhang, Y.; Chen, D.; Tan, H.; Zheng, Y.; Wang, J.; Chen, J. Characterization of Single-Nucleus Electrical Properties by Microfluidic Constriction Channel. Micromachines 2019, 10, 740. https://doi.org/10.3390/mi10110740
Liang H, Zhang Y, Chen D, Tan H, Zheng Y, Wang J, Chen J. Characterization of Single-Nucleus Electrical Properties by Microfluidic Constriction Channel. Micromachines. 2019; 10(11):740. https://doi.org/10.3390/mi10110740
Chicago/Turabian StyleLiang, Hongyan, Yi Zhang, Deyong Chen, Huiwen Tan, Yu Zheng, Junbo Wang, and Jian Chen. 2019. "Characterization of Single-Nucleus Electrical Properties by Microfluidic Constriction Channel" Micromachines 10, no. 11: 740. https://doi.org/10.3390/mi10110740
APA StyleLiang, H., Zhang, Y., Chen, D., Tan, H., Zheng, Y., Wang, J., & Chen, J. (2019). Characterization of Single-Nucleus Electrical Properties by Microfluidic Constriction Channel. Micromachines, 10(11), 740. https://doi.org/10.3390/mi10110740