
Sex Is a Determinant for Deoxynivalenol Metabolism and Elimination in the Mouse


Abstract
:1. Introduction
2. Results
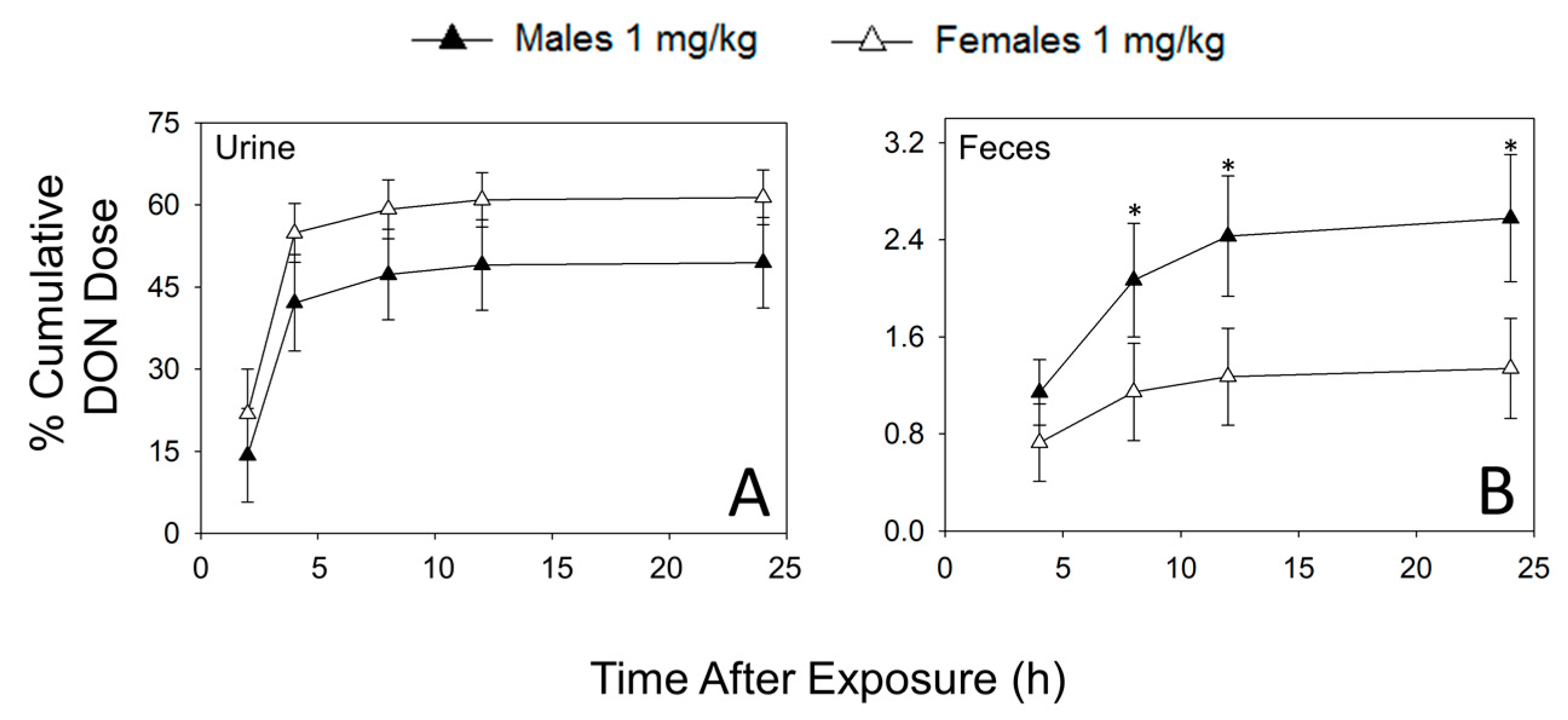
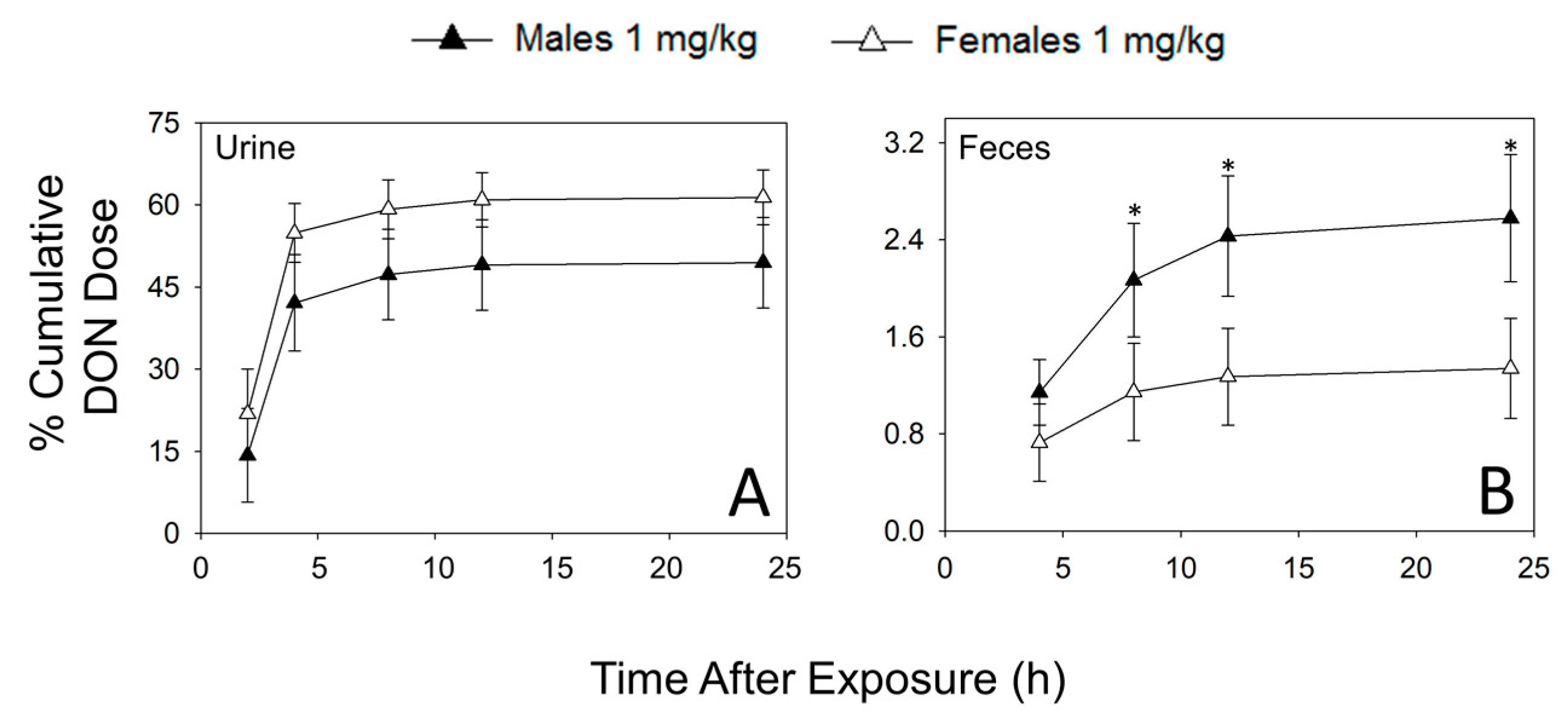
2.1. Urinary and Fecal Excretion of DON and DON Metabolites Differ by Sex
2.2. Hepatic and Renal Levels of DON Metabolites Differ by Sex
2.3. Ex Vivo DON Glucuronidation Rates by Microsomes Differ by Sex
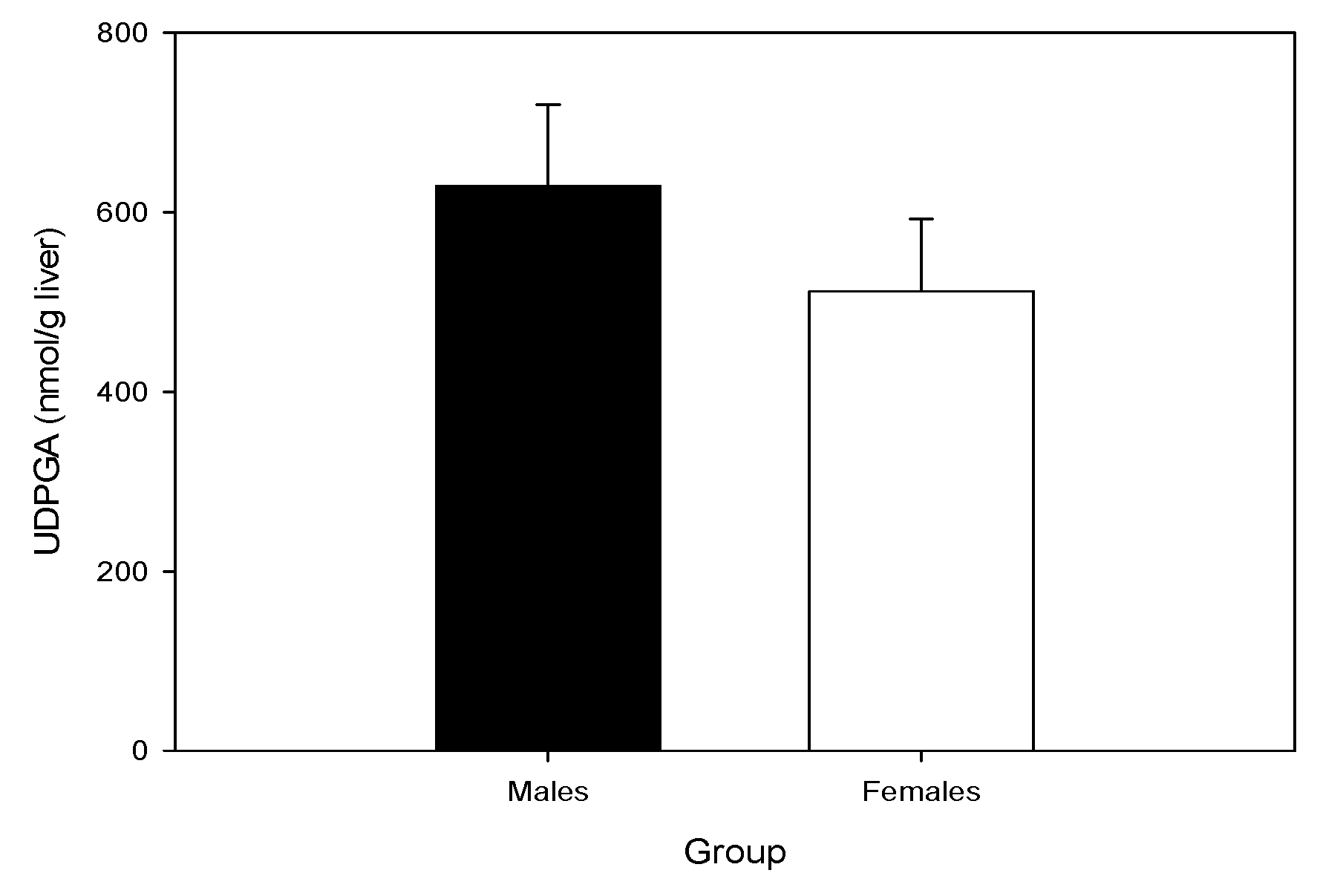
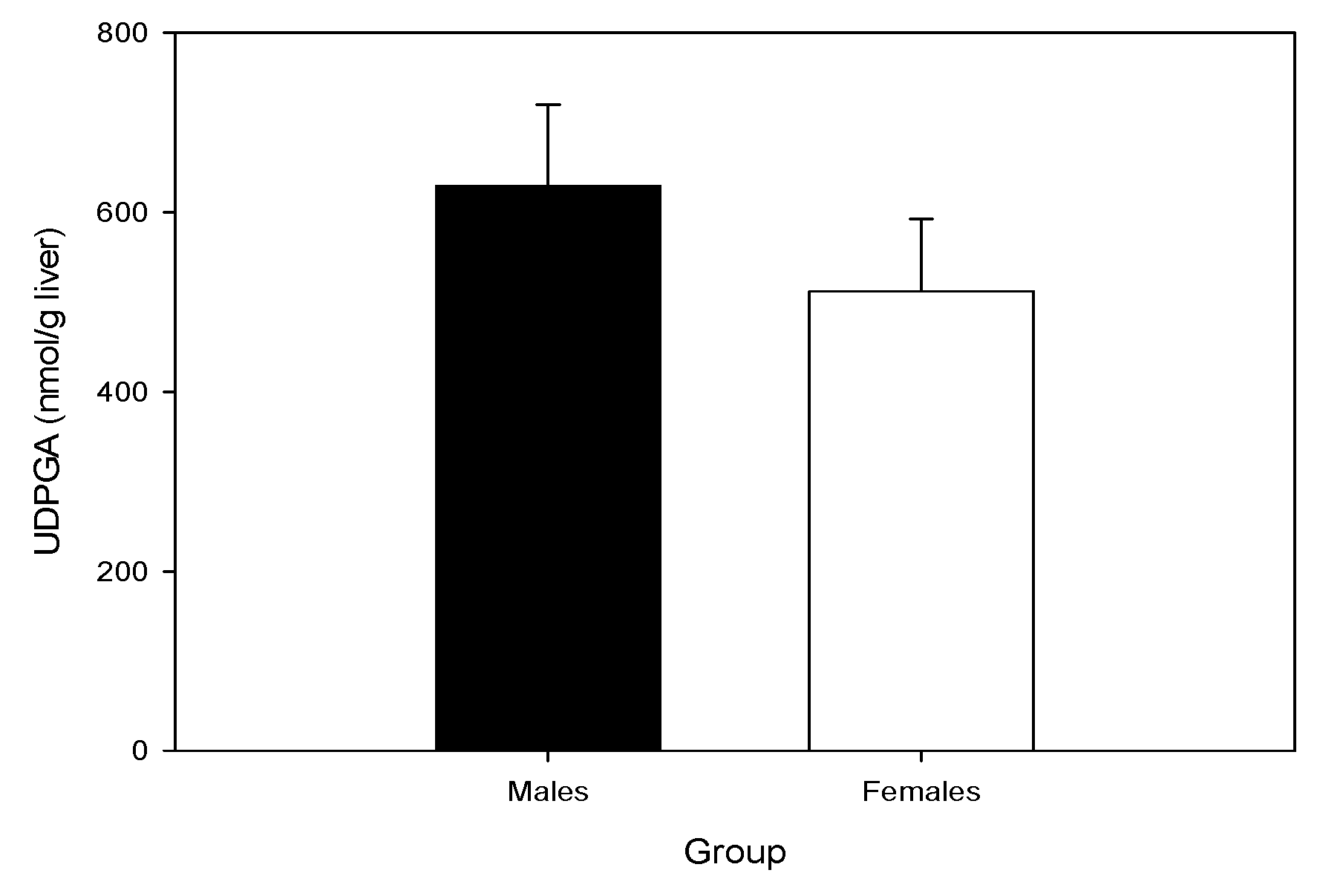
2.4. Hepatic UDP-GA Concentrations Do Not Differ by Sex
3. Discussion
4. Conclusions
5. Experimental
5.1. Animals
5.2. Chemicals and Reagents
5.3. Experimental Designs
5.4. ELISA of Urine and Feces
5.5. LC–MS/MS Analysis of Excreta and Tissues
5.6. Hepatic and Renal Microsomal Conversion of DON to DON Glucuronides
5.7. Determination of Hepatic UDP-GA Concentrations
5.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M. From the gut to the brain: Journey and pathophysiological effects of the food-associated trichothecene mycotoxin deoxynivalenol. Toxins (Basel) 2013, 5, 784–820. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.H.; Dohnal, V.; Huang, L.L.; Kuca, K.; Yuan, Z.H. Metabolic pathways of trichothecenes. Drug Metab. Rev. 2010, 42, 250–267. [Google Scholar] [CrossRef] [PubMed]
- Sarkanj, B.; Warth, B.; Uhlig, S.; Abia, W.A.; Sulyok, M.; Klapec, T.; Krska, R.; Banjari, I. Urinary analysis reveals high deoxynivalenol exposure in pregnant women from croatia. Food Chem. Toxicol. 2013, 62, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Warth, B.; Sulyok, M.; Fruhmann, P.; Berthiller, F.; Schuhmacher, R.; Hametner, C.; Adam, G.; Frohlich, J.; Krska, R. Assessment of human deoxynivalenol exposure using an LC-MS/MS based biomarker method. Toxicol. Lett. 2012, 211, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Schwartz-Zimmermann, H.E.; Hametner, C.; Nagl, V.; Fiby, I.; Macheiner, L.; Winkler, J.; Danicke, S.; Clark, E.; Pestka, J.J.; Berthiller, F. Glucuronidation of deoxynivalenol (DON) by different animal species: Identification of iso-DON glucuronides and iso-deepoxy-don glucuronides as novel don metabolites in pigs, rats, mice, and cows. Arch. Toxicol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, S.; Ivanova, L.; Faeste, C.K. Enzyme-assisted synthesis and structural characterization of the 3-, 8-, and 15-glucuronides of deoxynivalenol. J. Agric. Food Chem. 2013, 61, 2006–2012. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, S.; Ivanova, L.; Faeste, C.K. Correction to enzyme-assisted synthesis and structural characterization of the 3-, 8-, and 15-glucuronides of deoxynivalenol. J. Agric. Food Chem. 2016, 64, 3732. [Google Scholar] [CrossRef] [PubMed]
- Maul, R.; Warth, B.; Schebb, N.H.; Krska, R.; Koch, M.; Sulyok, M. In vitro glucuronidation kinetics of deoxynivalenol by human and animal microsomes and recombinant human UGT enzymes. Arch. Toxicol. 2015, 89, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Maul, R.; Warth, B.; Kant, J.S.; Schebb, N.H.; Krska, R.; Koch, M.; Sulyok, M. Investigation of the hepatic glucuronidation pattern of the fusarium mycotoxin deoxynivalenol in various species. Chem. Res. Toxicol. 2012, 25, 2715–2717. [Google Scholar] [CrossRef] [PubMed]
- Wan, D.; Huang, L.L.; Pan, Y.H.; Wu, Q.H.; Chen, D.M.; Tao, Y.F.; Wang, X.; Liu, Z.L.; Li, J.; Wang, L.Y.; et al. Metabolism, distribution, and excretion of deoxynivalenol with combined techniques of radiotracing, high-performance liquid chromatography ion trap time-of-flight mass spectrometry, and online radiometric detection. J. Agric. Food Chem. 2014, 62, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Schwartz-Zimmermann, H.E.; Fruhmann, P.; Danicke, S.; Wiesenberger, G.; Caha, S.; Weber, J.; Berthiller, F. Metabolism of deoxynivalenol and deepoxy-deoxynivalenol in broiler chickens, pullets, roosters and turkeys. Toxins (Basel) 2015, 7, 4706–4729. [Google Scholar] [CrossRef] [PubMed]
- Warth, B.; Del Favero, G.; Wiesenberger, G.; Puntscher, H.; Woelflingseder, L.; Fruhmann, P.; Sarkanj, B.; Krska, R.; Schuhmacher, R.; Adam, G.; et al. Identification of a novel human deoxynivalenol metabolite enhancing proliferation of intestinal and urinary bladder cells. Sci. Rep. 2016, 6, 33854. [Google Scholar] [CrossRef] [PubMed]
- Schwartz-Zimmermann, H.E.; Hametner, C.; Nagl, V.; Slavik, V.; Moll, W.D.; Berthiller, F. Deoxynivalenol (DON) sulfonates as major don metabolites in rats: From identification to biomarker method development, validation and application. Anal. Bioanal. Chem. 2014, 406, 7911–7924. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, T.; Takeda, H.; Ohi, T. Structure of a novel metabolite from deoxynivalenol, a trichothecene mycotoxin, in animals. Agric. Biol. Chem. 1983, 47, 2133–2135. [Google Scholar] [CrossRef]
- Worrell, N.R.; Mallett, A.K.; Cook, W.M.; Baldwin, N.C.P.; Shepherd, M.J. The role of gut microorganisms in the metabolism of deoxynivalenol administered to rats. Xenobiotica 1989, 19, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.W. Observations on flora of alimentary tract of animals and factors affecting its composition. J. Pathol. Microbiol. 1965, 89, 95–122. [Google Scholar] [CrossRef]
- Avantaggiato, G.; Havenaar, R.; Visconti, A. Evaluation of the intestinal absorption of deoxynivalenol and nivalenol by an in vitro gastrointestinal model, and the binding efficacy of activated carbon and other adsorbent materials. Food Chem. Toxicol. 2004, 42, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V.M.T.; Solfrizzo, M.; De Girolamo, A.; Chulze, S.N.; Torres, A.M.; Visconti, A. LC-MS/MS characterization of the urinary excretion profile of the mycotoxin deoxynivalenol in human and rat. J. Chrom. B Anal. Technol. Biomed. Life Sci. 2011, 879, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Gratz, S.W.; Duncan, G.; Richardson, A.J. The human fecal microbiota metabolizes deoxynivalenol and deoxynivalenol-3-glucoside and may be responsible for urinary deepoxy-deoxynivalenol. Appl. Environ. Microbiol. 2013, 79, 1821–1825. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.D.; Bates, M.A.; Bursian, S.J.; Link, J.E.; Flannery, B.M.; Sugita-Konishi, Y.; Watanabe, M.; Zhang, H.B.; Pestka, J.J. Comparison of emetic potencies of the 8-ketotrichothecenes deoxynivalenol, 15-acetyldeoxynivalenol, 3-acetyldeoxynivalenol, fusarenon X, and nivalenol. Toxicol. Sci. 2013, 131, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Flannery, B.M.; Wu, W.D.; Pestka, J.J. Characterization of deoxynivalenol-induced anorexia using mouse bioassay. Food Chem. Toxicol. 2011, 49, 1863–1869. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Pan, X.; Zhou, H.R.; Pestka, J.J. Modulation of inflammatory gene expression by the ribotoxin deoxynivalenol involves coordinate regulation of the transcriptome and translatome. Toxicol. Sci. 2013, 131, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol-induced proinflammatory gene expression: Mechanisms and pathological sequelae. Toxins (Basel) 2010, 2, 1300–1317. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.S.; Flannery, B.M.; Pestka, J.J. Murine anorectic response to deoxynivalenol (vomitoxin) is sex-dependent. Toxins (Basel) 2015, 7, 2845–2859. [Google Scholar] [CrossRef] [PubMed]
- Howell, S.R.; Klaassen, C. Circadian variation of hepatic UDP-glucuronic acid and the glucuronidation of xenobiotics in mice. Toxicol. Lett. 1991, 57, 73–79. [Google Scholar] [CrossRef]
- Eriksen, G.S.; Pettersson, H. Lack of de-epoxidation of Type B trichothecenes in incubates with human faeces. Food Addit. Contam. 2003, 20, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.C.; Hopton, R.P.; White, K.L.M.; Fisher, J.; Cade, J.E.; Wild, C.P. Assessment of deoxynivalenol metabolite profiles in uk adults. J. Agric. Food Chem. 2011, 49, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Schwartz-Zimmermann, H.E.; Wiesenberger, G.; Unbekannt, C.; Hessenberger, S.; Schatzmayr, D.; Berthiller, F. Reaction of (conjugated) deoxynivalenol with sulphur reagents-novel metabolites, toxicity and application. World Mycotoxin J. 2014, 7, 187–197. [Google Scholar] [CrossRef]
- Stechman, M.J.; Ahmad, B.N.; Loh, N.Y.; Reed, A.A.C.; Stewart, M.; Wells, S.; Hough, T.; Bentley, L.; Cox, R.D.; Brown, S.D.M.; et al. Establishing normal plasma and 24-hour urinary biochemistry ranges in C3H, Balb/c and C57Bl/6j mice following acclimatization in metabolic cages. Lab. Anim. 2010, 44, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Zhao, Y.L.; Nadai, M.; Naruhashi, K.; Shimizu, A.; Takagi, K.; Hasegawa, T. Gender-related differences in expression and function of hepatic p-glycoprotein and multidrug resistance-associated protein (MRP2) in rats. Life Sci. 2006, 79, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.D.; Csanaky, I.L.; Klaassen, C.D. Effects of aging on mRNA profiles for drug-metabolizing enzymes and transporters in livers of male and female mice. Drug Metab. Dispos. 2012, 40, 1216–1225. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, H.E.; Hametner, C.; Slavik, V.; Greitbauer, O.; Bichl, G.; Kunz-Vekiru, E.; Schatzmayr, D.; Berthiller, F. Characterization of three deoxynivalenol sulfonates formed by reaction of deoxynivalenol with sulfur reagents. J. Agric. Food Chem. 2013, 61, 8941–8948. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J.; Amuzie, C.J. Tissue distribution and proinflammatory cytokine gene expression following acute oral exposure to deoxynivalenol: Comparison of weanling and adult mice. Food Chem. Toxicol. 2008, 46, 2826–2831. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.S.; Flannery, B.M.; Gardner, E.M.; Pestka, J.J. High sensitivity of aged mice to deoxynivalenol (vomitoxin)-induced anorexia corresponds to elevated proinflammatory cytokine and satiety hormone responses. Toxins (Basel) 2015, 7, 4199–4215. [Google Scholar] [CrossRef] [PubMed]
- Lake, B.G. Preparation and characterization of microsomal fractions for studies of xenobiotic metabolism. In Biochemical Toxicology: A Partical Approach; Snell, K., Mullock, B., Eds.; IRL Press: Oxford, UK, 1987; pp. 183–251. [Google Scholar]
- Singh, J.; Schwarz, L.R.; Wiebel, F.J. A rapid enzymic procedure for the determination of picomole amounts of UDP-glucuronic acid. Biochem. J. 1980, 189, 369–372. [Google Scholar] [CrossRef] [PubMed]
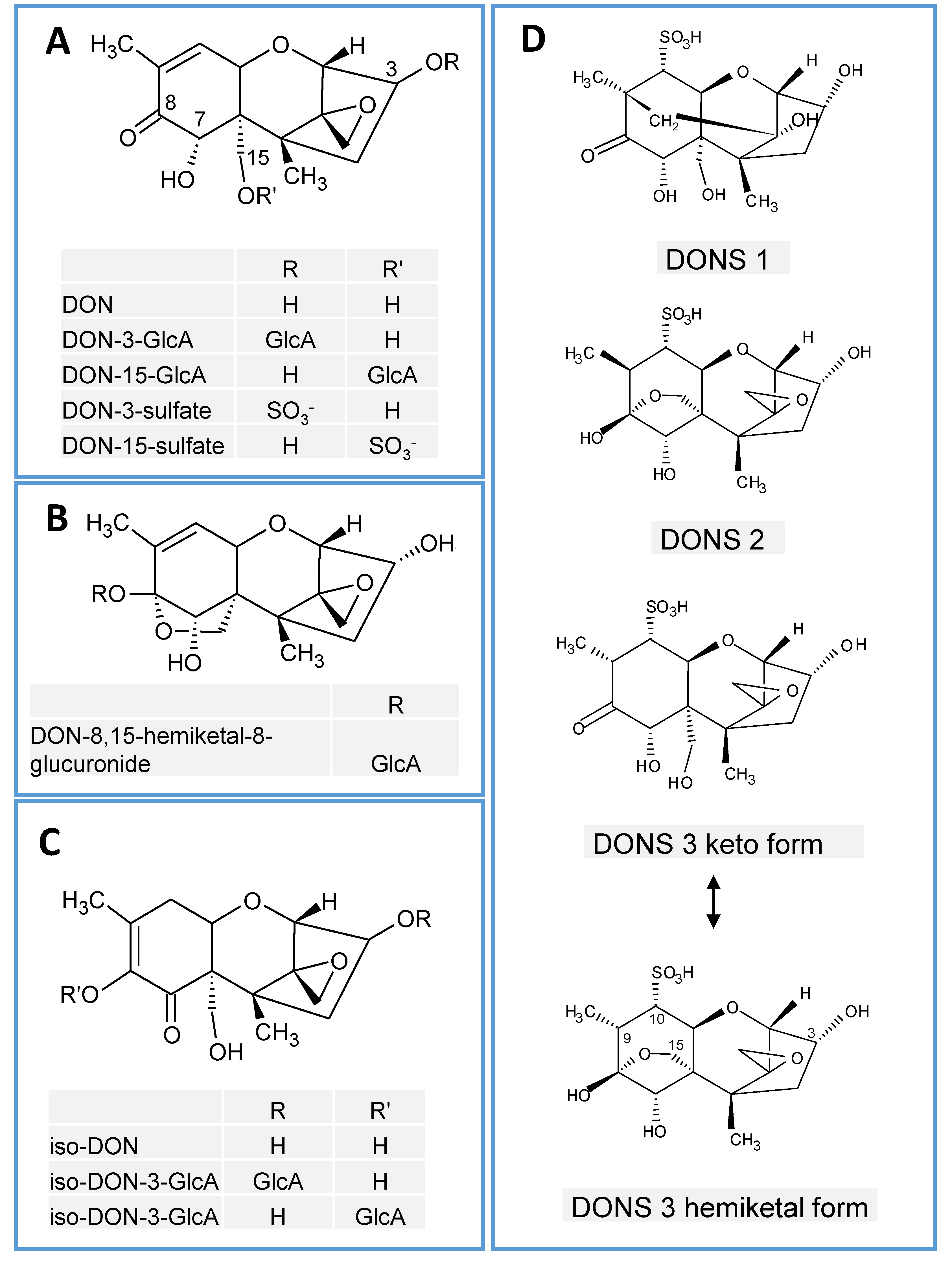

{kind=link}
{kind=link}
{kind=link}
| % Cumulative DON Dose | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Time | 2 h | 4 h | 8 h | 12 h | 24 h | |||||
| Sex | ♂ | ♀ | ♂ | ♀ | ♂ | ♀ | ♂ | ♀ | ♂ | ♀ |
| DON | 11.1 ± 0.4 | 20.4 ± 7.5 | 32.9 ± 7.0 | 50.4 ± 4.6 | 35.9 ± 6.9 | 53.8 ± 4.5 * | 37.1 ± 6.9 | 54.9 ± 4.3 * | 37.3 ± 6.9 | 55.2 ± 4.3 * |
| DON-3-GlcA | 7.6 ± 4.8 | 7.1 ± 2.7 | 25.0 ± 4.9 | 20.0 ± 2.2 | 27.9 ± 4.4 | 21.5 ± 2.2 | 28.9 ± 4.4 | 22.0 ± 2.1 | 29.1 ± 4.4 | 22.1 ± 2.1 |
| DON-8,15-hk-8-GlcA | 2.2 ± 1.4 | 2.4 ± 0.9 | 7.4 ± 1.4 | 6.7 ± 0.7 | 8.3 ± 1.3 | 7.3 ± 0.7 | 8.6 ± 1.3 | 7.4 ± 0.7 | 8.7 ± 1.3 | 7.4 ± 0.7 |
| iso-DON-8-GlcA | 0.9 ± 0.6 | 1.1 ± 0.4 | 3.1 ± 0.6 | 3.1 ± 0.3 | 3.6 ± 0.6 | 3.5 ± 0.3 | 3.9 ± 0.6 | 3.6 ± 0.3 | 4.0 ± 0.6 | 3.7 ± 0.3 |
| DON-3-sulfate | ND | 0.2 ± 0.1 * | ND | 0.5 ± 0.1 * | ND | 0.5 ± 0.1 * | ND | 0.5 ± 0.1 * | ND | 0.6 ± 0.1 * |
| DON-15-sulfate | ND | 0.2 ± 0.1 * | ND | 0.6 ± 0.1 | ND | 0.6 ± 0.0 | ND | 0.6 ± 0.1 * | ND | 0.6 ± 0.1 * |
| Total | 21.7 ± 13.1 | 31.1 ± 11.5 | 68.4 ± 13.9 | 81.2 ± 7.8 | 75.7 ± 13.1 | 87.2 ± 7.6 | 78.5 ± 13.2 | 89.1 ± 7.3 | 79.1 ± 13.1 | 89.6 ± 7.2 |
| % Cumulative DON Dose | ||||||||
|---|---|---|---|---|---|---|---|---|
| Time | 2 h | 4 h | 8 h | 12 h | ||||
| Sex | ♂ | ♀ | ♂ | ♀ | ♂ | ♀ | ♂ | ♀ |
| DON | 1.10 ± 0.24 | 0.68 ± 0.29 | 1.90 ± 0.41 | 1.00 ± 0.35 * | 2.32 ± 0.45 | 1.15 ± 0.36 * | 2.48 ± 0.49 | 1.21 ± 0.37 * |
| DON S1 | 0.04 ± 0.01 | 0.02 ± 0.01 | 0.29 ± 0.07 | 0.19 ± 0.03 | 0.62 ± 0.05 | 0.38 ± 0.03 * | 0.85 ± 0.08 | 0.47 ± 0.02 * |
| DON S2 | 0.16 ± 0.05 | 0.05 ± 0.02 * | 1.56 ± 0.43 | 0.73 ± 0.15 | 3.46 ± 0.40 | 1.51 ± 0.15 * | 4.93 ± 0.57 | 1.93 ± 0.16 * |
| DON S3 | 0.03 ± 0.01 | 0.01 ± 0.01 | 0.22 ± 0.09 | 0.13 ± 0.03 | 0.29 ± 0.10 | 0.15 ± 0.03 | 0.35 ± 0.04 | 0.17 ± 0.03 |
| Total | 1.34 ± 0.21 | 0.75 ± 0.31 | 3.93 ± 0.84 | 2.12 ± 0.43 | 6.70 ± 0.86 | 3.32 ± 0.44 | 8.30 ± 1.05 | 4.05 ± 0.43 |
| Concentration (nmol/g) | ||||||
|---|---|---|---|---|---|---|
| Time | 1 h | 2 h | 4 h | |||
| Sex | ♂ | ♀ | ♂ | ♀ | ♂ | ♀ |
| DON | 1.31 ± 0.15 | 1.76 ± 0.16 | 0.55 ± 0.09 | 0.59 ± 0.05 | 0.10 ± 0.01 | 0.09 ± 0.01 |
| DON-3-GlcA | 3.00 ± 0.35 | 1.45 ± 0.18 * | 1.10 ± 0.22 | 0.50 ± 0.10 * | 0.63 ± 0.30 | 0.12 ± 0.04 * |
| DON-8,15-hk-8-GlcA | 0.94 ± 0.13 | 0.46 ± 0.06 * | 0.33 ± 0.05 | 0.15 ± 0.02 * | 0.17 ± 0.08 | 0.03 ± 0.01 |
| iso-DON-8-GlcA | 0.22 ± 0.03 | 0.09 ± 0.02 * | 0.09 ± 0.02 | 0.03 ± 0.01 * | 0.08 ± 0.03 | 0.02 ± 0.01 |
| DON-3-sulfate | ND | 0.02 ± 0.003 * | ND | 0.01 ± 0.004 * | ND | ND |
| DON-15-sulfate | ND | 0.02 ± 0.004 * | ND | 0.02 ± 0.003 * | ND | ND |
| Concentration (nmol/g) | ||||||
|---|---|---|---|---|---|---|
| Time | 1 h | 2 h | 4 h | |||
| Sex | ♂ | ♀ | ♂ | ♀ | ♂ | ♀ |
| DON | 2.09 ± 0.14 | 1.76 ± 0.16 | 0.63 ± 0.08 | 0.59 ± 0.05 | 0.16± 0.01 | 0.09 ± 0.01 * |
| DON-3-GlcA | 2.86 ± 0.31 | 0.98 ± 0.09 * | 0.99 ± 0.12 | 0.33 ± 0.04 * | 0.17 ± 0.04 | 0.06 ± 0.01 * |
| DON-8,15-hk-8-GlcA | 2.33 ± 0.27 | 0.74 ± 0.03 * | 0.93 ± 0.11 | 0.21 ± 0.02 * | 0.15 ± 0.05 | 0.05 ± 0.01 |
| iso-DON-8-GlcA | 0.43 ± 0.04 | 0.08 ± 0.02 * | 0.19 ± 0.02 | 0.02 ± 0.01 * | 0.06 ± 0.02 | ND |
| DON Glucuronide Formation (pmol/min/mg protein) | |||||||
|---|---|---|---|---|---|---|---|
| Sex | Species | DON (µM) | DON-3-GlcA | DON-15-GlcA | iso-DON-8-GlcA | iso-DON-3-GlcA | DON-8,15 -hk -8-GlcA |
| ♂ | Mouse | 5 | 529 ± 239 | 72 ± 3 | 37 ± 1 | 33 ± 8 | 33 ± 8 |
| ♀ | Mouse | 5 | 279 ± 269 * | 35 ± 0 * | 18 ± 0 * | 18 ± 0 | 18 ± 0 |
| ♂ | Mouse | 20 | 1760 ± 309 | 210 ± 29 | 145 ± 22 | 108 ± 19 | 114 ± 17 |
| ♀ | Mouse | 20 | 1068 ± 31 | 212 ± 4 | 125 ± 5 | 90 ± 5 | 84 ± 4 |
| ♂ | Human | 5 | 15 ± 15 | 35 ± 0 | ND | ND | ND |
| ♀ | Human | 5 | ND | 35 ± 0 | ND | ND | ND |
| ♂ | Human | 20 | 44 ± 0 | 224 ± 2 | ND | ND | ND |
| ♀ | Human | 20 | 44 ± 0 | 231 ± 14 | ND | ND | ND |
| Analyte | Ret. Time (min) | Precursor Ion (m/z) | Ion Species | DP (V) | Product Ions (m/z) | CE (eV) | Ion Ratio (qual/quant) | LOD of Quant ** (ng/mL) |
|---|---|---|---|---|---|---|---|---|
| DON | 5.85 | 355.1 | [M+CH3CO2]− | −50 | 59.1/265.1 | −38/−18 | 0.24 | 0.7 |
| DOM-1 | 7.01 | 339.1 | [M+CH3CO2]− | −50 | 59.1/249.1 | −40/−17 | 0.36 | 0.6 |
| DON-3-sulfate | 4.98 | 375.1 | [M−H]− | −125 | 345.1/247.1 | −36/−38 | 0.80 | 0.2 |
| DON-15-sulfate | 4.87 | 375.1 | [M−H]− | −110 | 96.9/247.1 | −38/−38 | 0.06 | 0.03 |
| DOM-3-sulfate | 5.70 | 359.1 | [M−H]− | −125 | 96.9/329.1 | −38/−34 | 0.15 | 0.1 |
| DOM-15-sulfate | 5.83 | 359.1 | [M−H]− | −125 | 96.9/79.9 | −28/−118 | 0.48 | 0.06 |
| DON-3-GlcA | 5.36 | 471.1 | [M−H]− | −100 | 113.1/265.1 * | −35/−38 | 0.38 | 0.6 |
| DON-8,15-hk-8-GlcA | 4.50 | 471.1 | [M−H]− | −100 | 113.1/265.1 | −35/−38 | 0.12 | 0.5 |
| i-DON-8-GlcA | 2.71 | 471.1 | [M−H]− | −100 | 441.1/113.1 | −25/−35 | 0.47 | 0.4 |
| i-DON-3-GlcA | 5.72 | 471.1 | [M−H]− | −100 | 441.1/113.1 | −25/−35 | 0.13 | 0.4 |
| DONS 1 | 2.26 | 377.1 | [M−H]− | −130 | 79.9/331.0 | −98/−52 | 0.04 | 0.08 |
| DONS 2 | 2.82 | 377.1 | [M−H]− | −105 | 80.9/79.9 | −68/−98 | 0.15 | 0.08 |
| DONS 3 | 3.64 | 377.1 | [M−H]− | −130 | 79.9/347.0 | −98/−36 | 0.40 | 0.8 |
| DOMS 2 | 3.93 | 361.1 | [M−H]− | −25 | 80.9/249.1 | −65/−30 | 0.09 | 0.06 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pestka, J.J.; Clark, E.S.; Schwartz-Zimmermann, H.E.; Berthiller, F. Sex Is a Determinant for Deoxynivalenol Metabolism and Elimination in the Mouse. Toxins 2017, 9, 240. https://doi.org/10.3390/toxins9080240
Pestka JJ, Clark ES, Schwartz-Zimmermann HE, Berthiller F. Sex Is a Determinant for Deoxynivalenol Metabolism and Elimination in the Mouse. Toxins. 2017; 9(8):240. https://doi.org/10.3390/toxins9080240
Chicago/Turabian StylePestka, James J., Erica S. Clark, Heidi E. Schwartz-Zimmermann, and Franz Berthiller. 2017. "Sex Is a Determinant for Deoxynivalenol Metabolism and Elimination in the Mouse" Toxins 9, no. 8: 240. https://doi.org/10.3390/toxins9080240
APA StylePestka, J. J., Clark, E. S., Schwartz-Zimmermann, H. E., & Berthiller, F. (2017). Sex Is a Determinant for Deoxynivalenol Metabolism and Elimination in the Mouse. Toxins, 9(8), 240. https://doi.org/10.3390/toxins9080240