
Regulation of Toxin Production in Clostridium perfringens

Abstract
:1. Introduction
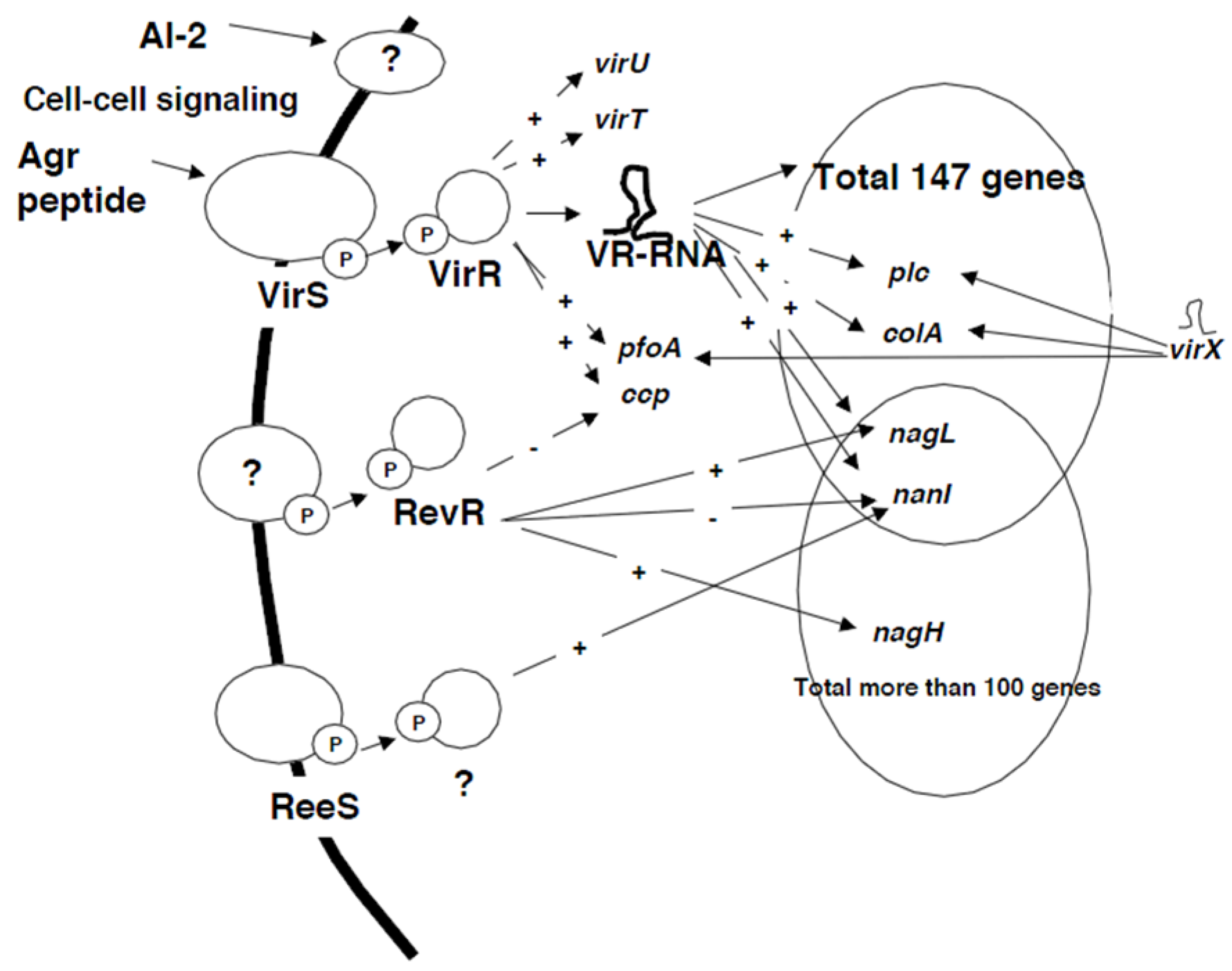
2. The Regulation of Toxin Genes by Two-Component Regulatory Systems
2.1. The Regulation of Toxin Genes by the VirS/VirR System
2.2. The RevR System
2.3. ReeS (Regulator of Extracellular Enzymes Sensor)
2.4. Regulation by RNA Molecules
2.5. VR-RNA
2.6. VirT and VirU
2.7. VirX
3. Regulation of Toxin Genes by Cell-Cell Communication
3.1. The Autoinducer 2 (AI-2) System
3.2. The agr System
4. Other Types of Regulation
CodY
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Hatheway, C.L. Toxigenic clostridia. Clin. Microbiol. Rev. 1990, 3, 66–98. [Google Scholar] [CrossRef] [PubMed]
- Petit, L.; Gibert, M.; Popoff, M.R. Clostridium perfringens: Toxinotype and genotype. Trends Microbiol. 1999, 7, 104–110. [Google Scholar] [CrossRef]
- Rood, J.I. Virulence genes of Costridium perfringens. Annu. Rev. Microbiol. 1998, 52, 333–360. [Google Scholar] [CrossRef] [PubMed]
- Hassan, K.A.; Elbourne, L.D.; Tetu, S.G.; Melville, S.B.; Rood, J.I.; Paulsen, I.T. Genomic analyses of Clostridium perfringens isolates from five toxinotypes. Res. Microbiol. 2015, 166, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Ohtani, K.; Hirakawa, H.; Ohshima, K.; Yamashita, A.; Shiba, T.; Ogasawara, N.; Hattori, M.; Kuhara, S.; Hayashi, H. Complete genome sequence of Clostridium perfringens, an anaerobic flesh-eater. Proc. Natl. Acad. Sci. USA 2002, 99, 996–1001. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Shimizu, T. Regulation of toxin gene expression in Clostridium perfringens. Res. Microbiol. 2015, 166, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Wanner, B.L. Is cross regulation by phosphorylation of two-component respomse regulator proteins important in bacteria? J. Bacteriol. 1992, 174, 2053–2058. [Google Scholar] [PubMed]
- Dziejman, M.; Mekalanos, J.J. Two-component signal transduction and its role in the expression of bacterial virulance factors. In Two-Component Signal Transduction; Hoch, J.A., Silhavy, T.J., Eds.; American Society for Microbiology: Washington, WA, USA, 1995; pp. 305–317. [Google Scholar]
- Shimizu, T.; Ba-Thein, W.; Tamaki, M.; Hayashi, H. The virR gene, a member of a class of two-component response regulators, regulates the production of the perfringolysin O, collagenase, and hemagglutinin in Clostridium perfringens. J. Bacteriol. 1994, 176, 1616–1623. [Google Scholar] [PubMed]
- Ba-Thein, W.; Lyristis, M.; Ohtani, K.; Nisbet, I.T.; Hayashi, H.; Rood, J.I.; Shimizu, T. The virr/virs locus regulates the transcription of genes encoding extracellular toxin production in Clostridium perfringens. J. Bactreriol. 1996, 178, 2514–2520. [Google Scholar]
- Lyristis, M.; Bryant, A.E.; Sloan, J.; Awad, M.M.; Nisbet, I.T.; Stevens, D.L.; Rood, J.I. Identification and molecular analysis of a locus that regulates extracellular toxin production in Clostridium perfringens. Mol. Microbiol. 1994, 12, 761–777. [Google Scholar] [CrossRef] [PubMed]
- Banu, S.; Ohtani, K.; Yaguchi, H.; Swe, T.; Cole, S.T.; Hayashi, H.; Shimizu, T. Identification of novel virr/virs-regulated genes in Clostridium perfringens. Mol. Microbol. 2000, 35, 854–864. [Google Scholar] [CrossRef]
- Shimizu, T.; Yaguchi, H.; Ohtani, K.; Banu, S.; Hayashi, H. Clostridial VirR/VirS regulon involves a regulatory RNA molecule for expression of toxins. Mol. Microbiol. 2002, 43, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Cheung, J.K.; Keyburn, A.L.; Carter, G.P.; Lanckriet, A.L.; Van Immerseel, F.; Moore, R.J.; Rood, J.I. The VirSR two-component signal transduction system regulates NetB toxin production in Clostridium perfringens. Infect. Immun. 2010, 78, 3064–3072. [Google Scholar] [CrossRef] [PubMed]
- Cheung, J.K.; Dupuy, B.; Deveson, D.S.; Rood, J.I. The spatial organization of the virr boxes is critical for virr-mediated expression of the perfringolysin O gene, pfoA, from Clostridium perfringens. J. Bacteriol. 2004, 186, 3321–3330. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Kawsar, H.I.; Okumura, K.; Hayashi, H.; Shimizu, T. The virr/virs regulatory cascade affects transcription of plasmid-encoded putative virulence genes in Clostridium perfringens. FEMS Microbiol. Lett. 2003, 222, 137–141. [Google Scholar] [CrossRef]
- Ma, M.; Vidal, J.; Saputo, J.; McClane, B.A.; Uzal, F. The virs/virr two-component system regulates the anaerobic cytotoxicity, intestinal pathogenicity, and enterotoxemic lethality of Clostridium perfringens type C isolate CN3685. mBio 2011, 2, e00338-10. [Google Scholar] [CrossRef] [PubMed]
- Sayeed, S.; Uzal, F.A.; Fisher, D.J.; Saputo, J.; Vidal, J.E.; Chen, Y.; Gupta, P.; Rood, J.I.; McClane, B.A. Beta toxin is essential for the intestinal virulence of Clostridium perfringens type C disease isolate CN3685 in a rabbit ileal loop model. Mol. Microbiol. 2008, 67, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Vidal, J.E.; Ohtani, K.; Shimizu, T.; McClane, B.A. Contact with enterocyte-like caco-2 cells induces rapid upregulation of toxin production by clostridium perfringens type C isolates. Cell Microbiol. 2009, 11, 1306–1328. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ma, M.; Uzal, F.A.; McClane, B.A. Host cell-induced signaling causes Clostridium perfringens to upregulate production of toxins important for intestinal infections. Gut Microbes 2014, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gibert, M.; Petit, L.; Raffestin, S.; Okabe, A.; Popoff, M.R. Clostridium perfringens iota-toxin requires activation of both binding and enzymatic components for cytopathic activity. Infect. Immun. 2000, 68, 3848–3853. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Hirakawa, H.; Tashiro, K.; Yoshizawa, S.; Kuhara, S.; Shimizu, T. Identification of a two-component VirR/VirS regulon in Clostridium perfringens. Anaerobe 2010, 16, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Hiscox, T.J.; Harrison, P.F.; Chakravorty, A.; Choo, J.M.; Ohtani, K.; Shimizu, T.; Cheung, J.K.; Rood, J.I. Regulation of sialidase production in Clostridium perfringens by the orphan sensor histidine kinase rees. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Lamarche, M.G.; Wanner, B.L.; Crepin, S.; Harel, J. The phosphate regulon and bacterial virulence: A regulatory network connecting phosphate homeostasis and pathogenesis. FEMS Microbiol. Rev. 2008, 32, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.E.; Hoch, J.A. Essentiality, bypass, and targeting of the YycFG (VicRK) two-component regulatory system in gram-positive bacteria. J. Bacteriol. 2008, 190, 2645–2648. [Google Scholar] [CrossRef] [PubMed]
- Hiscox, T.J.; Chakravorty, A.; Choo, J.M.; Ohtani, K.; Shimizu, T.; Cheung, J.K.; Rood, J.I. Regulation of virulence by the RevrR response regulator in Clostridium perfringens. Infect. Immun. 2011, 79, 2145–2153. [Google Scholar] [CrossRef] [PubMed]
- Johansson, J.; Cossart, P. RNA-mediated control of virulence gene expression in bacterial pathogens. Trends Microbiol. 2003, 11, 280–285. [Google Scholar] [CrossRef]
- Romby, P.; Charpentier, E. An overview of RNAs with regulatory functions in gram-positive bacteria. Cell Mol. Life Sci. 2009, 67, 217–237. [Google Scholar] [CrossRef] [PubMed]
- Geissmann, T.; Possedko, M.; Huntzinger, E.; Fechter, P.; Ehresmann, C.; Romby, P. Regulatory RNAs as mediators of virulence gene expression in bacteria. Handb. Exp. Pharmacol. 2006, 137, 9–43. [Google Scholar]
- Romby, P.; Vandenesch, F.; Wagner, E.G. The role of RNAs in the regulation of virulence-gene expression. Curr. Opin. Microbiol. 2006, 9, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Okumura, K.; Ohtani, K.; Hayashi, H.; Shimizu, T. Characterization of genes regulated directly by the virr/virs system in Clostridium perfringens. J. Bacteriol. 2008, 190, 7719–7727. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Bhowmik, S.K.; Hayashi, H.; Shimizu, T. Identification of a novel locus that regulates expression of toxin genes in Clostridium perfringens. FEMS Microbiol. Lett. 2002, 209, 109–114. [Google Scholar] [CrossRef]
- Ohtani, K.; Hirakawa, H.; Paredes-Sabja, D.; Tashiro, K.; Kuhara, S.; Sarker, M.R.; Shimizu, T. Unique regulatory mechanism of sporulation and enterotoxin production in Clostridium perfringens. J. Bacteriol. 2013, 195, 2931–2936. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed]
- Bassler, B.L. How bacteria talk to each other: Regulation of gene expression by quorum sensing. Curr. Opin. Microbiol. 1999, 2, 582–587. [Google Scholar] [CrossRef]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold. Spring Harb. Perspect. Med. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev Bio.l 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed]
- Bassler, B.L. Small talk. Cell-to-cell communication in bacteria. Cell 2002, 109, 421–424. [Google Scholar] [CrossRef]
- Lyon, G.J.; Novick, R.P. Peptide signaling in Staphylococcus aureus and other gram-positive bacteria. Peptides 2004, 25, 1389–1403. [Google Scholar] [CrossRef] [PubMed]
- Schauder, S.; Shokat, K.; Surette, M.G.; Bassler, B.L. The luxs family of bacterial autoinducers: Biosynthesis of a novel quorum-sensing signal molecule. Mol. Microbiol. 2001, 41, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Hayashi, H.; Shimizu, T. The luxs gene is involved in cell-cell signaling for toxin production in Clostridium perfringens. Mol. Microbiol. 2002, 44, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Xavier, K.B.; Bassler, B.L. Luxs quorum sensing: More than just a numbers game. Curr. Opin. Microbiol. 2003, 6, 191–197. [Google Scholar] [CrossRef]
- Federle, M.J. Autoinducer-2-based chemical communication in bacteria: Complexities of interspecies signaling. Contrib. Microbiol. 2009, 16, 18–32. [Google Scholar] [PubMed]
- Belizario, J.E.; Napolitano, M. Human microbiomes and their roles in dysbiosis, common diseases, and novel therapeutic approaches. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.; Novick, R.P. Autocrine regulation of toxin synthesis by Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 1995, 92, 1619–1623. [Google Scholar] [CrossRef] [PubMed]
- Novick, R.P.; Muir, T.W. Virulence gene regulation by peptides in Staphylococci and other gram-positive bacteria. Curr. Opin. Microbiol. 1999, 2, 40–45. [Google Scholar] [CrossRef]
- Peng, H.-L.; Novic, R.P.; Kreiswirth, B.; Kornblum, J.; Schlievert, P. Cloning, characterization, and sequencing of an accessory gene regulator (agr) in Staphylococcus aureus. J. Bacteriol. 1988, 170, 4365–4372. [Google Scholar] [PubMed]
- George, E.A.; Muir, T.W. Molecular mechanisms of agr quorum sensing in virulent staphylococci. Chembiochem 2007, 8, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Autret, N.; Raynaud, C.; Dubail, I.; Berche, P.; Charbit, A. Identification of the agr locus of listeria monocytogenes: Role in bacterial virulence. Infect. Immun. 2003, 71, 4463–4471. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Yuan, Y.; Hassan, S.; Wang, R.; Wang, Y.; Shimizu, T. Virulence gene regulation by the agr system in Clostridium perfringens. J. Bacteriol. 2009, 191, 3919–3927. [Google Scholar] [CrossRef] [PubMed]
- Higashi, Y.; Chazono, M.; Inoue, K.; Yanagase, Y.; Amano, T.; Shimada, K. Complementation of theta toxinogenecity between mutants of two groups of Clostridium perfringens. Biken. J. 1973, 16, 1–9. [Google Scholar] [PubMed]
- Imagawa, T.; Tatsuki, T.; Higashi, Y.; Amano, T. Complementation characteristics of newly isolated mutants from two groups of strains of Clostridium perfringens. Biken J. 1981, 24, 13–21. [Google Scholar] [PubMed]
- Imagawa, T.; Higashi, Y. An activity which restores theta toxin activity in some theta toxin-deficient mutants of Clostridium perfringens. Microbiol. Immunol. 1992, 36, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Rood, J.I.; Wilkinson, R.G. Isolation and characterization of Clostridium perfringens mutants altered in broth hemagglutinin and sialidase production. J. Bacteriol. 1975, 123, 419–427. [Google Scholar] [PubMed]
- Li, J.; Chen, J.; Vidal, J.E.; McClane, B.A. The agr-like quorum-sensing system regulates sporulation and production of enterotoxin and beta2 toxin by Clostridium perfringens type A non-food-borne human gastrointestinal disease strain F5603. Infect. Immun. 2011, 79, 2451–2459. [Google Scholar] [CrossRef] [PubMed]
- Novick, R.P. Autoinduction and signal transduction in the regulation of Staphylococcal virulence. Mol. Microbiol. 2003, 48, 1429–1449. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; McClane, B.A. Role of the agr-like quorum-sensing system in regulating toxin production by Clostridium perfringens type B strains cn1793 and cn1795. Infect. Immun. 2012, 80, 3008–3017. [Google Scholar] [CrossRef] [PubMed]
- Vidal, J.E.; Ma, M.; Saputo, J.; Garcia, J.; Uzal, F.A.; McClane, B.A. Evidence that the agr-like quorum sensing system regulates the toxin production, cytotoxicity and pathogenicity of Clostridium perfringens type C isolate CN3685. Mol. Microbiol. 2012, 83, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Rood, J.I.; McClane, B.A. Epsilon-toxin production by Clostridium perfringens type D strain CN3718 is dependent upon the agr operon but not the VirS/VirR two-component regulatory system. mBio 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ma, M.; Sarker, M.R.; McClane, B.A. CodY is a global regulator of virulence-associated properties for Clostridium perfringens type D strain CN3718. mBio 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Stenz, L.; Francois, P.; Whiteson, K.; Wolz, C.; Linder, P.; Schrenzel, J. The CodY pleiotropic repressor controls virulence in gram-positive pathogens. FEMS Immunol. Med. Microbiol. 2011, 62, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Chateau, A.; van Schaik, W.; Joseph, P.; Handke, L.D.; McBride, S.M.; Smeets, F.M.; Sonenshein, A.L.; Fouet, A. Identification of CodY targets in Bacillus anthracis by genome-wide in vitro binding analysis. J. Bacteriol. 2013, 195, 1204–1213. [Google Scholar] [CrossRef] [PubMed]
- Van Schaik, W.; Chateau, A.; Dillies, M.A.; Coppee, J.Y.; Sonenshein, A.L.; Fouet, A. The global regulator CodY regulates toxin gene expression in Bacillus anthracis and is required for full virulence. Infect. Immun. 2009, 77, 4437–4445. [Google Scholar] [CrossRef] [PubMed]
- Dineen, S.S.; Villapakkam, A.C.; Nordman, J.T.; Sonenshein, A.L. Repression of Clostridium difficile toxin gene expression by CodY. Mol. Microbiol. 2007, 66, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Belitsky, B.R.; Sonenshein, A.L. Genome-wide identification of Bacillus subtilis CodY-binding sites at single-nucleotide resolution. Proc. Natl. Acad. Sci. USA 2013, 110, 7026–7031. [Google Scholar] [CrossRef] [PubMed]
- Bouillaut, L.; Dubois, T.; Sonenshein, A.L.; Dupuy, B. Integration of metabolism and virulence in Clostridium difficile. Res. Microbiol. 2015, 166, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Connan, C.; Popoff, M.R. Two-component systems and toxinogenesis regulation in Clostridium botulinum. Res. Microbiol. 2015, 166, 332–343. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Type | Toxins Produced | |||
|---|---|---|---|---|
| Alpha | Beta | Epsilon | Iota | |
| A | + | − | − | − |
| B | + | + | + | − |
| C | + | + | − | − |
| D | + | − | + | − |
| E | + | − | − | + |
| Type | Toxin (gene) | Regulation by VirR/VirS Ref: [9,10,14,16,17,22,57] | Regulation by agr Ref: [50,55,57,58,59] | VR-RNA Ref: [12,13,22] | VirT Ref: [31] | VirU Ref: [31] | VirX Ref: [32] | CodY Ref: [60] | ReeS Ref: [23] | RevR Ref: [26] |
|---|---|---|---|---|---|---|---|---|---|---|
| A | CPA (plc) | Yes (Yes) | Yes (Yes) | (Yes) | No | No | (Yes) | NR | No | No |
| PFO (pfoA) | Yes (Yes) | Yes (Yes) | No | (Yes) | (Yes) | (Yes) | NR | No | No | |
| collagenase (colA) | (Yes) | (Yes) | (Yes) | No | No | (Yes) | NR | No | No | |
| sialidase (nanI) | Yes | NR | (Yes) | NR | NR | NR | NR | (Yes) | (Yes) | |
| Hyaluronidase (nagH) | (Yes) | NR | (Yes) | NR | NR | NR | NR | No | (Yes) | |
| Hyaluronidase (nagL) | (Yes) | NR | (Yes) | NR | NR | NR | NR | No | (Yes) | |
| α-clostripain (ccp) | (Yes) | NR | No | (Yes) | (Yes) | NR | NR | No | (Yes) | |
| CPE (cpe) | NR | Yes | NR | NR | NR | NR | NR | No | No | |
| NetB (netB) | Yes | NR | NR | NR | NR | NR | NR | No | No | |
| CPB2 (cpb2) | Yes | Yes | (Yes) | NR | NR | NR | NR | No | NR | |
| B | CPA (plc) | NR | Yes | NR | NR | NR | NR | NR | NR | NR |
| PFO (pfoA) | NR | Yes | NR | NR | NR | NR | NR | NR | NR | |
| CPB (cpb) | NR | Yes | NR | NR | NR | NR | NR | NR | NR | |
| ETX (etx) | NR | No | NR | NR | NR | NR | NR | NR | NR | |
| CPB2 (cpb2) | NR | No | NR | NR | NR | NR | NR | NR | NR | |
| C | CPA (plc) | Yes | Yes | NR | NR | NR | NR | NR | NR | NR |
| PFO (pfoA) | Yes | Yes | NR | NR | NR | NR | NR | NR | NR | |
| CPB (cpb) | Yes | Yes | NR | NR | NR | NR | NR | NR | NR | |
| D | CPA (plc) | Yes | Yes | NR | NR | NR | NR | No | NR | NR |
| PFO (pfoA) | Yes | Yes | NR | NR | NR | NR | No | NR | NR | |
| ETX (etx) | No | Yes(Yes) | NR | NR | NR | NR | Yes | NR | NR | |
| E | ITX (iap and ibp) | NR (for activity Yes) | NR | NR | NR | NR | NR | NR | NR | NR |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ohtani, K.; Shimizu, T. Regulation of Toxin Production in Clostridium perfringens. Toxins 2016, 8, 207. https://doi.org/10.3390/toxins8070207
Ohtani K, Shimizu T. Regulation of Toxin Production in Clostridium perfringens. Toxins. 2016; 8(7):207. https://doi.org/10.3390/toxins8070207
Chicago/Turabian StyleOhtani, Kaori, and Tohru Shimizu. 2016. "Regulation of Toxin Production in Clostridium perfringens" Toxins 8, no. 7: 207. https://doi.org/10.3390/toxins8070207
APA StyleOhtani, K., & Shimizu, T. (2016). Regulation of Toxin Production in Clostridium perfringens. Toxins, 8(7), 207. https://doi.org/10.3390/toxins8070207