
Polyphasic Approach Including MALDI-TOF MS/MS Analysis for Identification and Characterisation of Fusarium verticillioides in Brazilian Corn Kernels


 ,
,  and
and
Abstract
:1. Introduction
2. Results
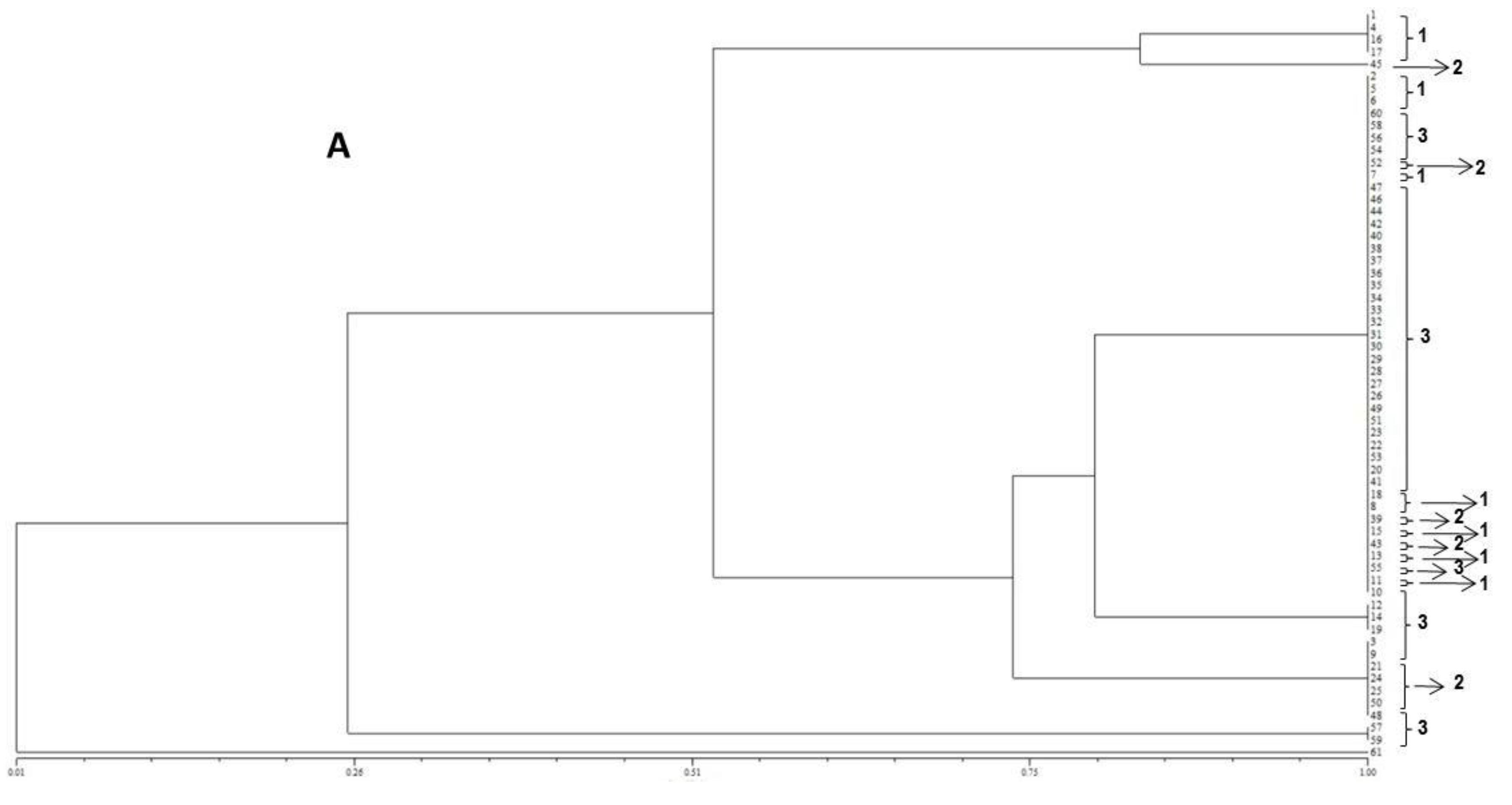
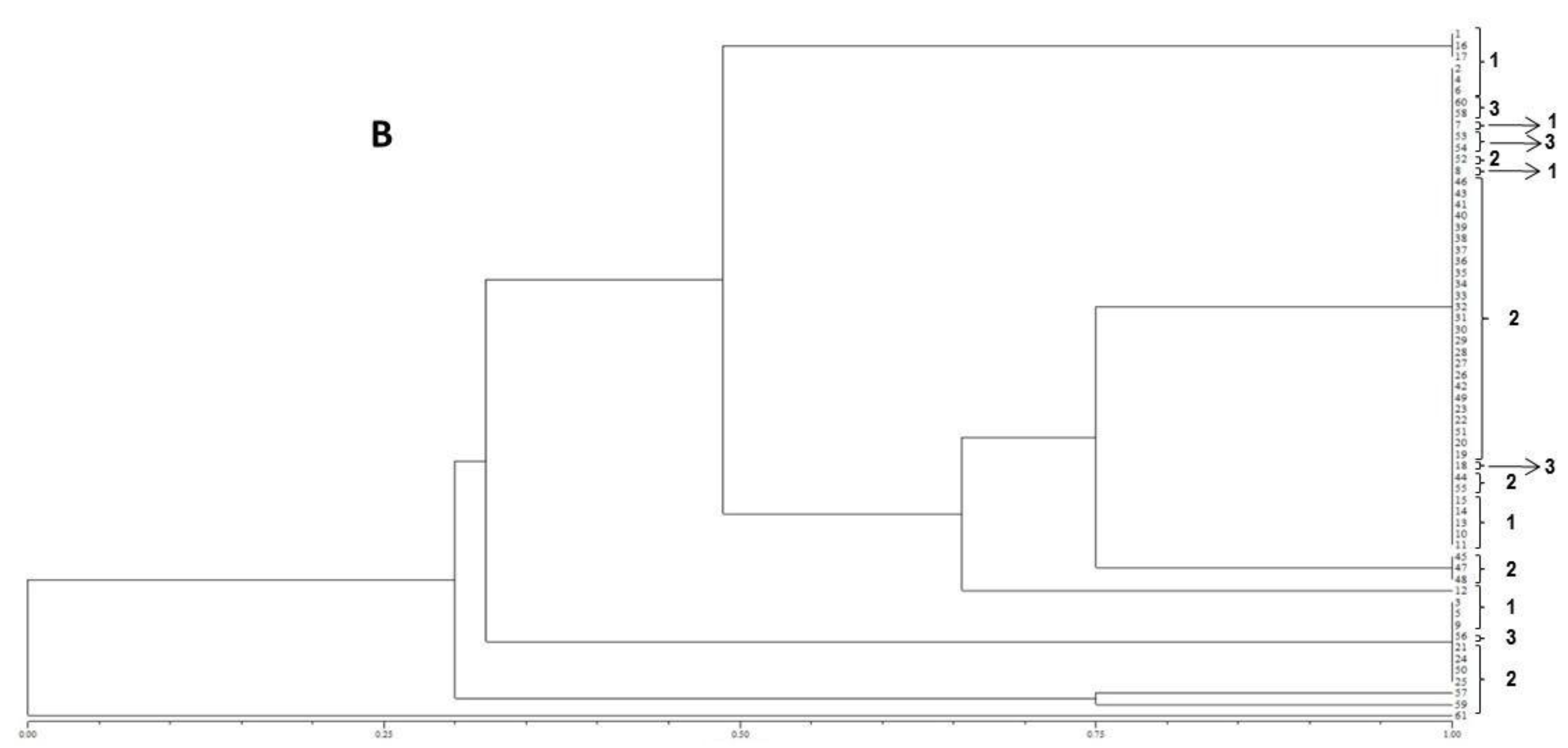
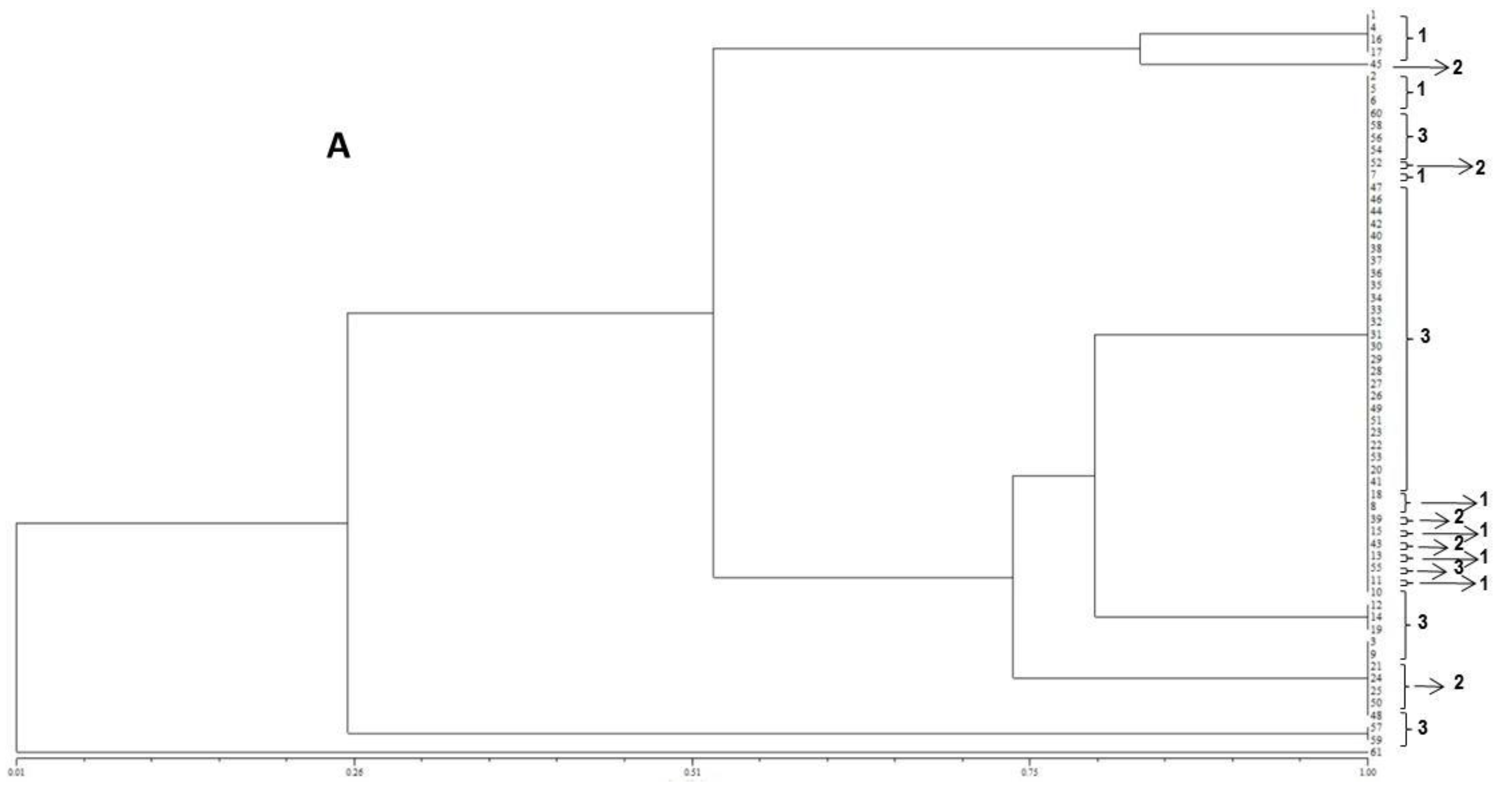
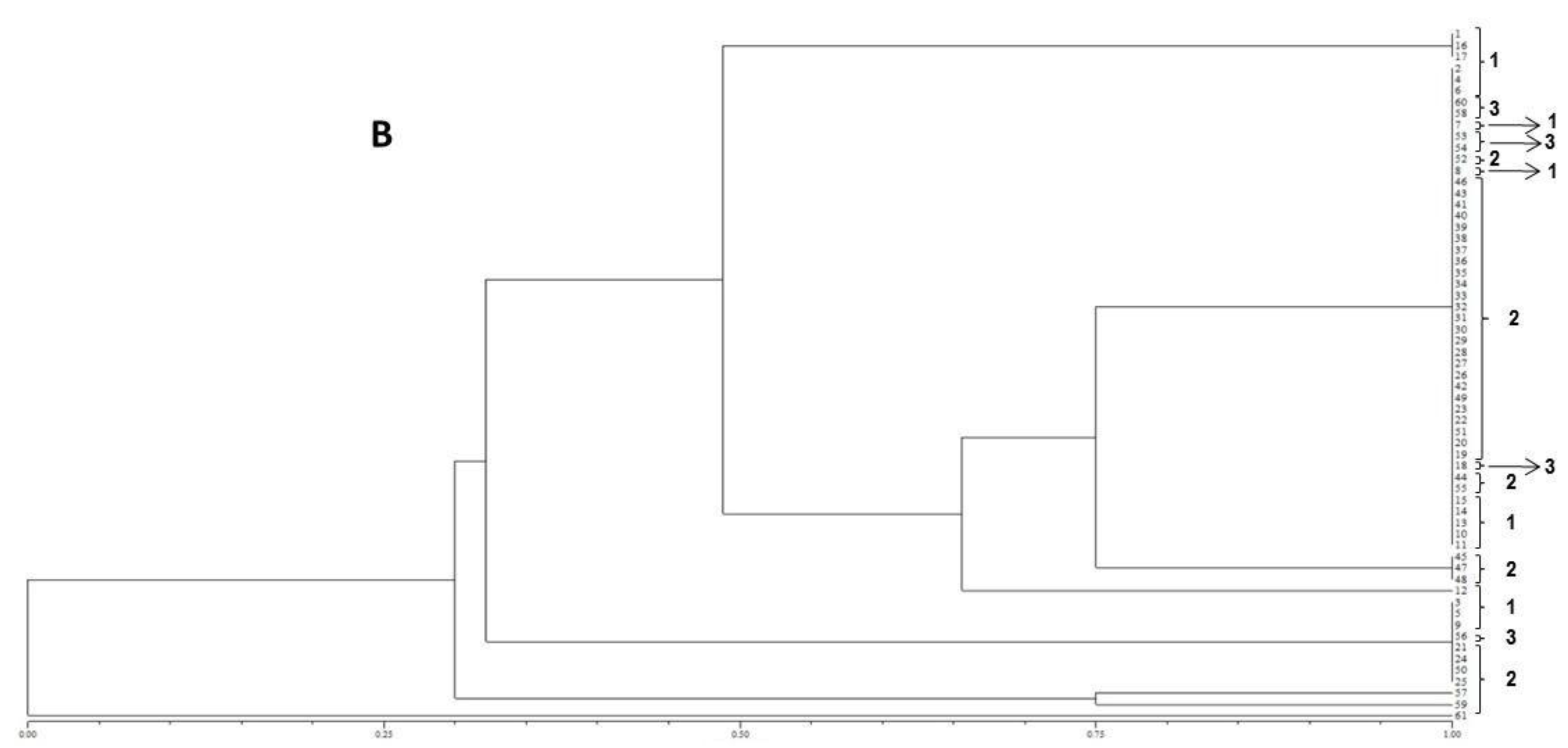
2.1. Isolation and Identification of Fungal Strains
2.2. Detection of FUM1 Gene
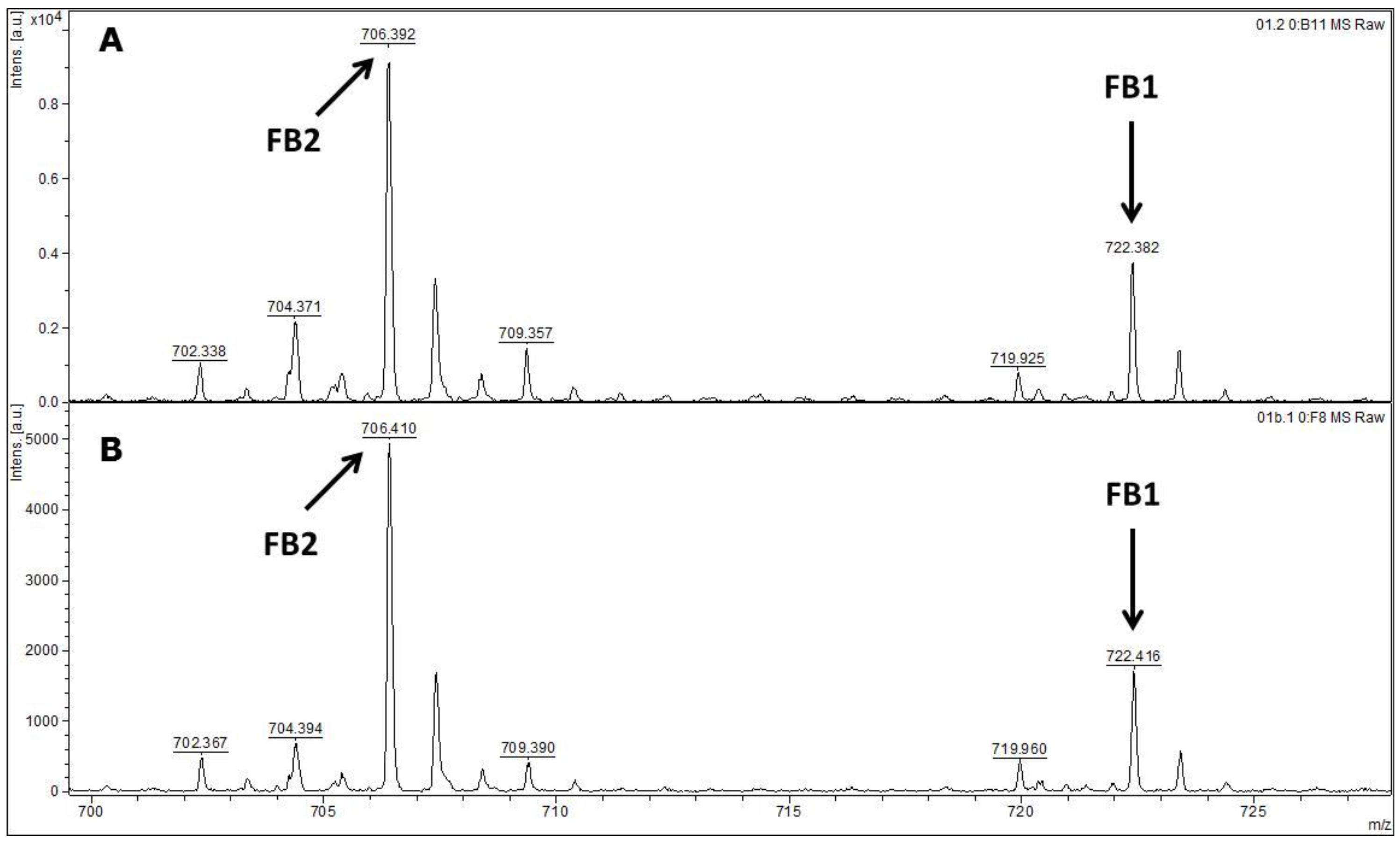
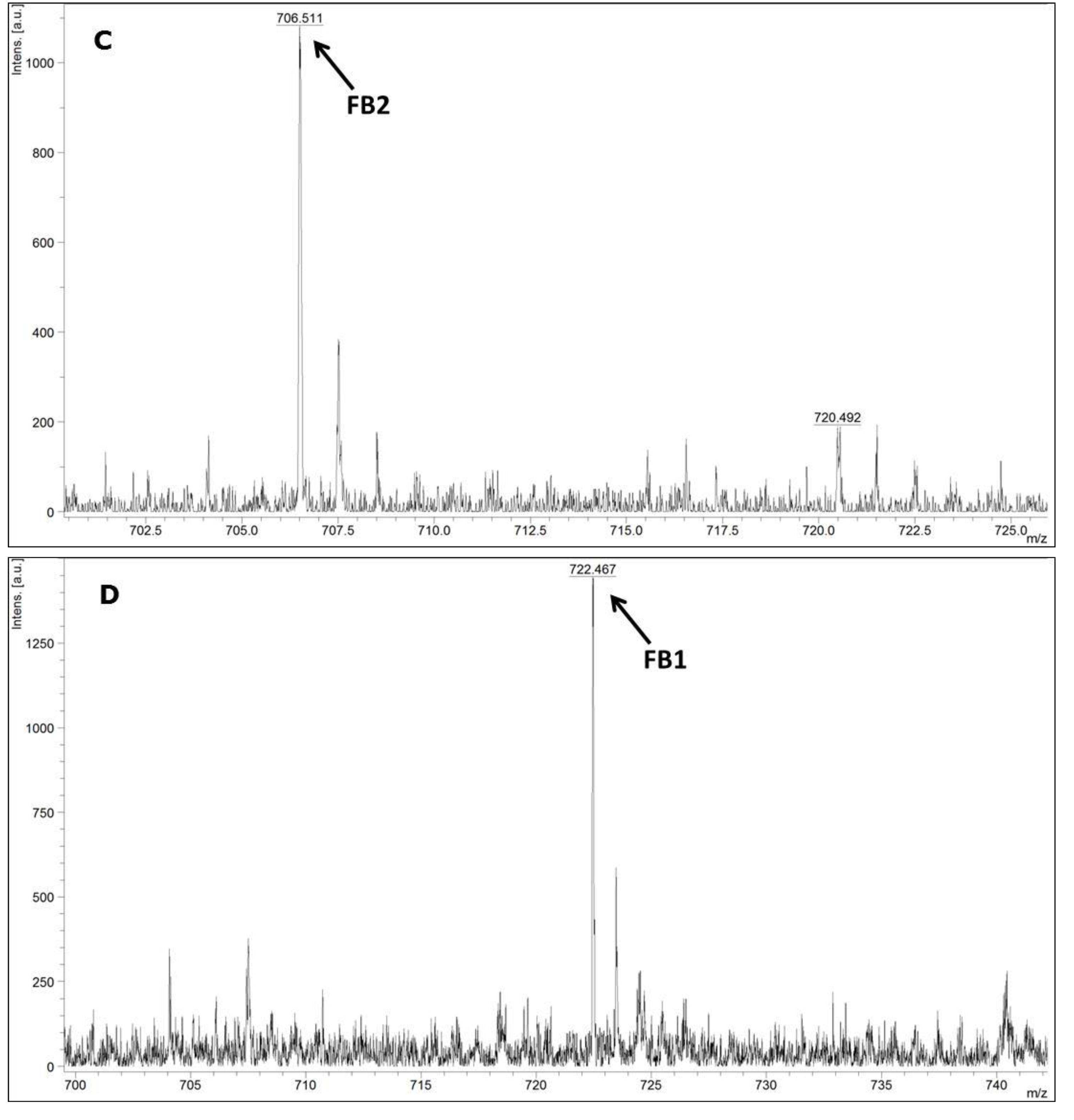
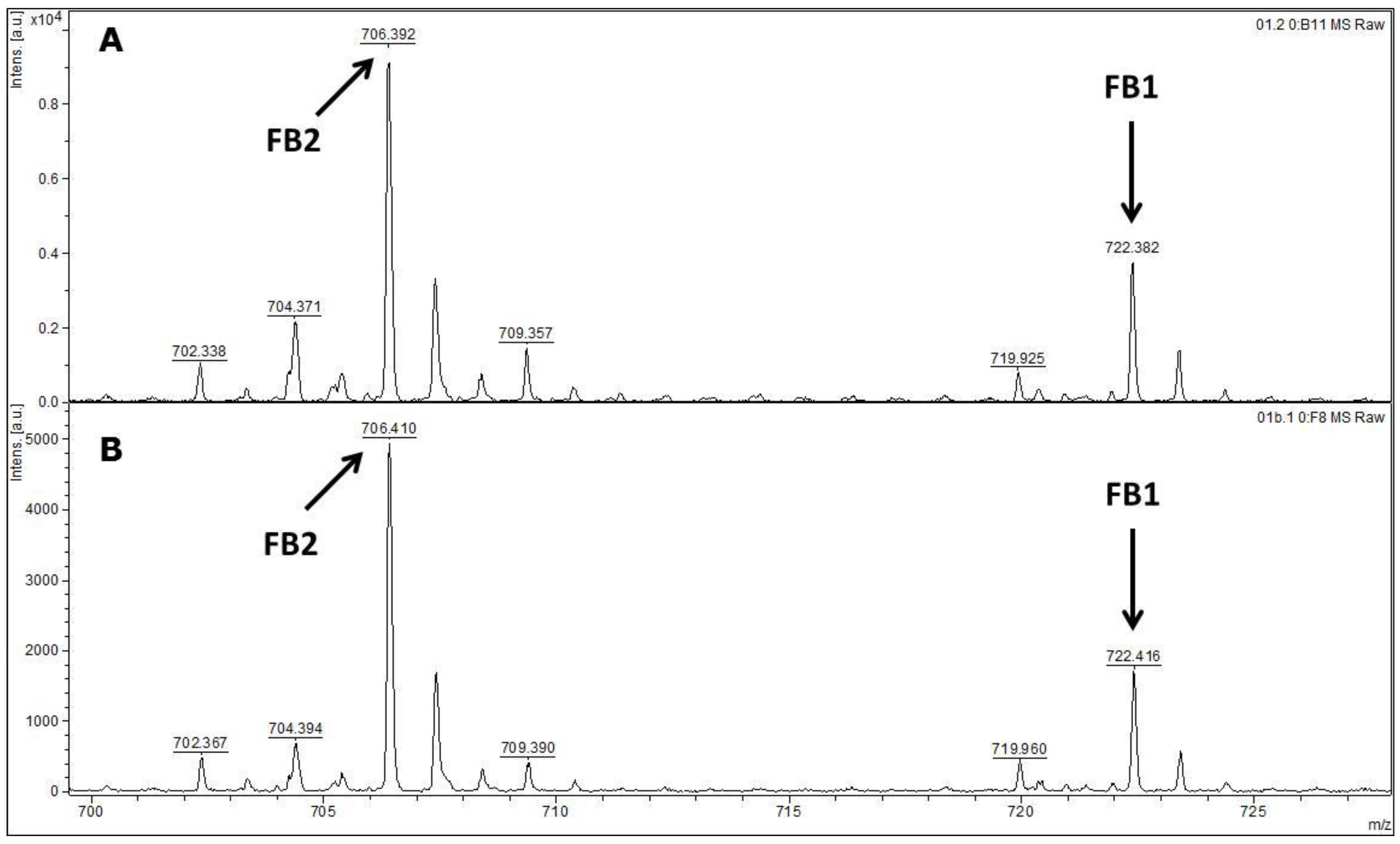
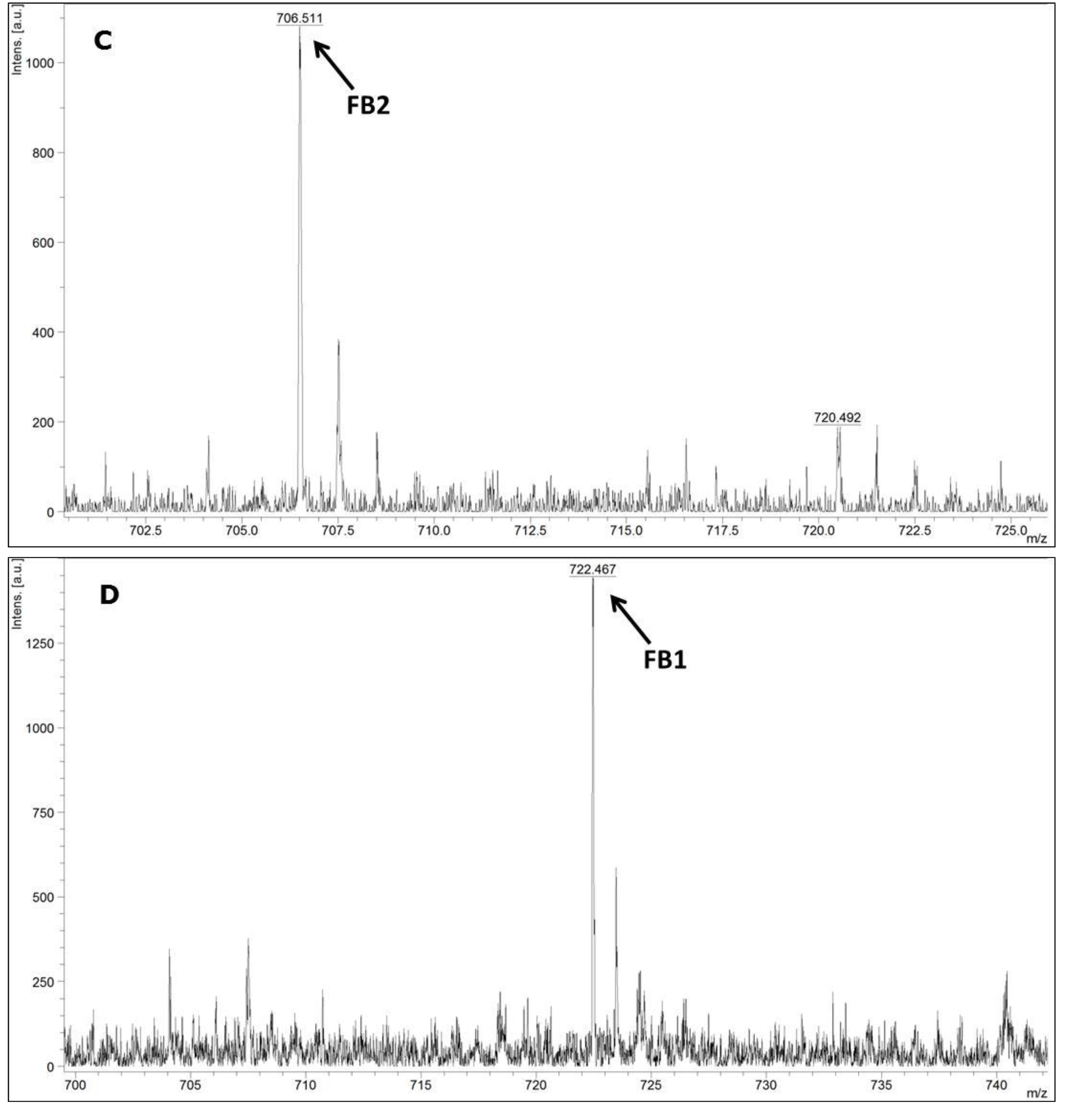
2.3. Detection of Fumonisins by MALDI-TOF MS/MS
3. Discussion
4. Experimental Section
4.1. Corn Kernel Samples
4.2. Fungal Isolation and Identification
4.3. Mycelium Growth and DNA Extraction
4.4. Molecular Biology
4.5. Analyses of Genetic Variability
4.6. Statistical Analyses NTSYS-PC
4.7. Fungal Identification by MALDI-TOF MS
4.8. Detection of Fumonisins by MALDI-TOF MS/MS
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Duarte, J.O.; Cruz, J.C.; Garcia, J.C.; Mattoso, M.J. Economia da Produção. In Cultivo do Milho, 6th ed.; Cruz, J.C., Ed.; Embrapa Milho e Sorgo: Sete Lagoas, Brazil, 2010. (In Portuguese) [Google Scholar]
- Maciel, M.; Ottoni, C.; Santos, C.; Lima, N.; Moreira, K.; Souza-Motta, C. Production of polygalacturonases by Aspergillus section Nigri strains in a fixed bed reactor. Molecules 2013, 18, 1660–1671. [Google Scholar] [CrossRef] [PubMed]
- Maciel, M.D.H.C.; Ottoni, C.A.; Herculano, P.N.; Porto, T.S.; Porto, A.L.F.; Santos, C.; Lima, N.; Moreira, K.A.; Souza-Motta, C. Purification of polygalacturonases produced by Aspergillus niger using an aqueous two-phase system. Fluid Phase Equilib. 2014, 371, 125–130. [Google Scholar] [CrossRef]
- Ottoni, C.A.; Santos, C.; Kozakiewicz, Z.; Lima, N. White-rot fungi capable of decolourising textile dyes under alkaline conditions. Folia Microbiol. 2013, 58, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Passarini, M.R.Z.; Santos, C.; Lima, N.; Berlinck, R.G.S.; Sette, L.D. Filamentous fungi from the Atlantic marine sponge Dragmacidon reticulatum. Arch. Microbiol. 2013, 195, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Paterson, R.R.M.; Sariah, M.; Lima, N.; Zainal-Abidin, M.A.; Santos, C. Mutagenic and inhibitory compounds produced by fungi affect detrimentally diagnosis and phylogenetic analyses. Curr. Bioact. Comp. 2008, 4, 245–257. [Google Scholar]
- Siqueira, V.; Oliveira, H.M.B.; Santos, C.; Paterson, R.R.M.; Gusmão, N.B.; Lima, N. Filamentous fungi in drinking water, particularly in relation to biofilm formation. Int. J. Environ. Res. Public Health 2011, 8, 456–469. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, V.M.; Oliveira, H.M.B.; Santos, C.; Paterson, R.R.M.; Gusmão, N.; Lima, N. Biofilms from a Brazilian water distribution system include filamentous fungi. Can. J. Microbiol. 2013, 59, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Souza, P.N.C.; Grigoletto, T.L.B.; Moraes, L.A.B.; Abreu, L.M.; Guimarães, L.H.S.; Santos, C.; Galvão, L.R.; Cardoso, P.G. Production and chemical characterization of pigments in filamentous fungi. Microbiology 2016, 162. [Google Scholar] [CrossRef]
- Marín, P.; Magan, N.; Vázquez, C.; González-Jaén, M.T. Differential effect of environmental conditions on the growth and regulation of the fumonisin biosynthetic gene FUM1 in the maize pathogens and fumonisin producers Fusarium verticillioides and Fusarium proliferatum. FEMS Microbiol. Ecol. 2010, 73, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, M.L.M.C.; Tanaka, S.; Ito, S. Fumonisin B1 production by Fusarium proliferatum strains isolated from Allium fistulosum plants and seeds in Japan. Lett. Appl. Microbiol. 2009, 48, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Maheshwar, P.K.; Moharram, S.A.; Janardhana, G.R. Detection of fumonisin producing Fusarium verticillioides in paddy (Oryza sativa L.) using polymerase chain reaction (PCR). Braz. J. Microbiol. 2009, 40, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rangel, D.; Sanjuan-Badillo, A.; Plasencia, J. Fumonisin production by Fusarium verticillioides isolated from maize in Mexico and development of a polymerase chain reaction to detect potential toxigenic strains in grains. J. Agric. Food Chem. 2005, 53, 8565–8571. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.N.; Araújo, J.; Durigon, E.L.; Corrêa, B. Sequence variability in the FUM1 gene of Fusarium verticillioides strains. Can. J. Microbiol. 2007, 53, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasa, M.Y.; González-Jaen, M.T.; Dass, R.S.; Raj, A.P.C.; Janardhana, G.R. A PCR-based assay for the detection and differentiation of potential fumonisin producing Fusarium verticillioides isolated from Indian Maize Kernels. Food Biotechnol. 2010, 22, 160–170. [Google Scholar] [CrossRef]
- Bayraktar, H.; Dolar, F.S.; Maden, S. Use of RAPD and ISSR markers in detection of genetic variation and population structure among Fusarium oxysporum f. sp. ciceris Isolates on Chickpea in Turkey. J. Phytopathol. 2008, 156, 146–154. [Google Scholar] [CrossRef]
- González-Jaén, M.T.; Mirete, S.; Patiño, B.; López-Errasquín, E.; Vázquez, C. Genetic markers for the analysis of variability and for production of specific diagnostic sequences in fumonisin-producing strains of Fusarium verticillioides. Eur. J. Plant Pathol. 2004, 110, 525–532. [Google Scholar] [CrossRef]
- Rodrigues, P.; Santos, C.; Venâncio, A.; Lima, N. Species identification of Aspergillus section Flavi isolates from Portuguese almonds using phenotypic, including MALDI-TOF ICMS, and molecular approaches. J. Appl. Microbiol. 2011, 111, 877–892. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.; Paterson, R.M.R.; Venâncio, A.; Lima, N. Filamentous fungal characterisations by matrix-assisted laser desorption/ionisation time of flight mass spectrometry. J. Appl. Microbiol. 2010, 108, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.C.; Chalfoun, S.M.; Batista, L.R.; Santos, C.; Lima, N. Use of a polyphasic approach including MALDI-TOF MS for identification of Aspergillus section Flavi strains isolated from food commodities in Brazil. Ann. Microbiol. 2015, 65, 2119–2129. [Google Scholar] [CrossRef]
- Pereira, L.; Dias, N.; Santos, C.; Lima, N. The use of MALDI-TOF ICMS as an alternative tool for Trichophyton rubrum identification and typing. Enferm. Infecc. Microbiol. Clín. 2014, 32, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.M.E.; Santos, C.; Sampaio, P.; Romeo, O.; Almeida-Paes, R.; Pais, C.; Lima, N.; Zancope-Oliveira, R.M. Development and optimization of a new MALDI-TOF protocol for identification of the Sporothrix species complex. Res. Microbiol. 2015, 166, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Marvin, L.F.; Roberts, M.A.; Fay, L.B. Matrix assisted laser desorption ⁄ionization time-of-flight mass spectrometry in clinical chemistry. Clin. Chim. Acta 2003, 337, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Kemptner, J.; Marchetti-Deschmann, M.; Mach, R.; Druzhinina, I.S.; Kubicek, C.P.; Allmaier, G. Evaluation of matrix-assisted laser desorption/ionization (MALDI) preparation techniques for surface characterization of intact Fusarium spores by MALDI linear time-of flight mass spectrometry. Rapid Commun. Mass Spectrom. 2009, 23, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Kemptner, J.; Marchetti-Deschmann, M.; Kubicek, C.P.; Allmaier, G. Development of a MALDI two-layer volume sample preparation technique for analysis of colored conidia spores of Fusarium by MALDI linear TOF mass spectrometry. Anal. Bioanal. Chem. 2009, 395, 1373–1383. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Marchetti-Deschmann, M.; Winkler, W.; Lohninger, H.; Allmaier, G. Intact cell/spore mass spectrometry of Fusarium macro conidia for fast isolate and species differentiation. In Detection of Biological Agents for the Prevention of Bioterrorism; NATO Science for Peace and Security Series—A: Chemistry and Biology; Banoub, J., Ed.; Springer: Dordrecht, The Netherlands, 2009; pp. 47–63. [Google Scholar]
- Santos, C.; Ventura, J.A.; Costa, H.; Fernandes, P.M.B.; Lima, N. MALDI-TOF MS to identify the pineapple pathogen Fusarium guttiforme and its antagonist Trichoderma asperellum on decayed pineapple. Trop. Plant Pathol. 2015, 40, 227–232. [Google Scholar] [CrossRef]
- Santos, C.; Capela, R.; Pereira, C.S.G.P.; Valente, E.; Gouveia, L.; Pannecouque, C.; De Clercq, E.; Moreira, R.; Gomes, P. Structure-activity relationships for dipeptide prodrugs of acyclovir: Implications for prodrug design. Eur. J. Med. Chem. 2009, 44, 2339–2346. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing Professional: Sydney, Australia, 2006. [Google Scholar]
- Marín, P.; Moretti, A.; Ritieni, A.; Jurado, M.; Vázquez, C.; González-Jaén, M. Phylogenetic analyses and toxigenic profiles of Fusarium equiseti and Fusarium acuminatum isolated from cereals from Southern Europe. Food Microbiol. 2012, 31, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, V.; Rajaraja, R.; Patharjan, S.; Karthikeyan, P.; Saravanakumar, P.; Siva, M.; Aruna Bhavani, P.S.; Palani, P. PCR based detection of fumonisin producing strains of Fusarium verticillioides and gene related to toxin production. Curr. Bot. 2011, 2, 34–37. [Google Scholar]
- Lima, M.L.F. Caracterização Molecular de Espécies de Metarhizium e Patogenicidade Sobre Diatraea saccharalis. Ph.D. Thesis, Federal University of Pernambuco, Recife, Brazil, 2005. [Google Scholar]
- Dias, N.; Santos, C.; Portela, M.; Lima, N. Toenail onychomycosis in a Portuguese geriatric population. Mycopathologia 2011, 172, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Makun, H.A.; Dutton, M.F.; Njobeh, P.B.; Phoku, J.Z.; Yah, C.S. Incidence, phylogeny and mycotoxigenic potentials of fungi isolated from rice in Niger State, Nigeria. J. Food Saf. 2011, 31, 334–349. [Google Scholar] [CrossRef]
- Kong, W.; Xie, T.; Li, J.; Wei, J.; Qiu, F.; Qi, A.; Zheng, Y.; Yang, M. Analysis of fumonisins B1 and B2 in spices and aromatic and medicinal herbs by HPLC-FLD with on-line post-column derivatization and positive confirmation by LC-MS/MS. Analyst 2012, 137, 3166–3174. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F.; Brandão-Filho, S.P. Geographical expansion of visceral leishmaniasis in the State of Pernambuco. Rev. Soc. Bras. Med. Trop. 2006, 39, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Patino, B.; Mirete, S.; Gonzalez-Jaen, T.; Mule, G.; Vasquez, C. PCR detection assay for fumonisin producing Fusarium verticillioides strains. J. Food Prot. 2004, 67, 1278–1283. [Google Scholar] [PubMed]
- Lieckfeldt, E.; Meyer, W.; Börner, T. Rapid identification and differentiation of yeasts by DNA and PCR fingerprinting. J. Basic Microbiol. 1993, 33, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Meyer, W.; Mitchell, T.G. Polymerase chain reaction fingerprint in fungi three using single primers specific to minisatelites and simple repetitive DNA sequence: Strain variation in Cryptococcus neoformans. Eletrophoresis 1995, 6, 1649–1656. [Google Scholar]
- Rohlf, F.J. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System; Applied Biostatistics: Setauket, NY, USA, 1988. [Google Scholar]
- Jaccard, P. Nouvelles recherches sur la distribuition horale. Bull. Soc. Sci. Nat. 1908, 44, 223–270. [Google Scholar]
- Ono, E.Y.S.; Fungaro, M.H.P.; Sofia, S.H.; Miguel, T.A.; Sugiura, Y.; Hirooka, E.Y. Fusarium verticillioides strains isolated from corn feed: Characterization by fumonisin production and RAPD fingerprinting. Braz. Arch. Biol. Technol. 2010, 53, 953–960. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
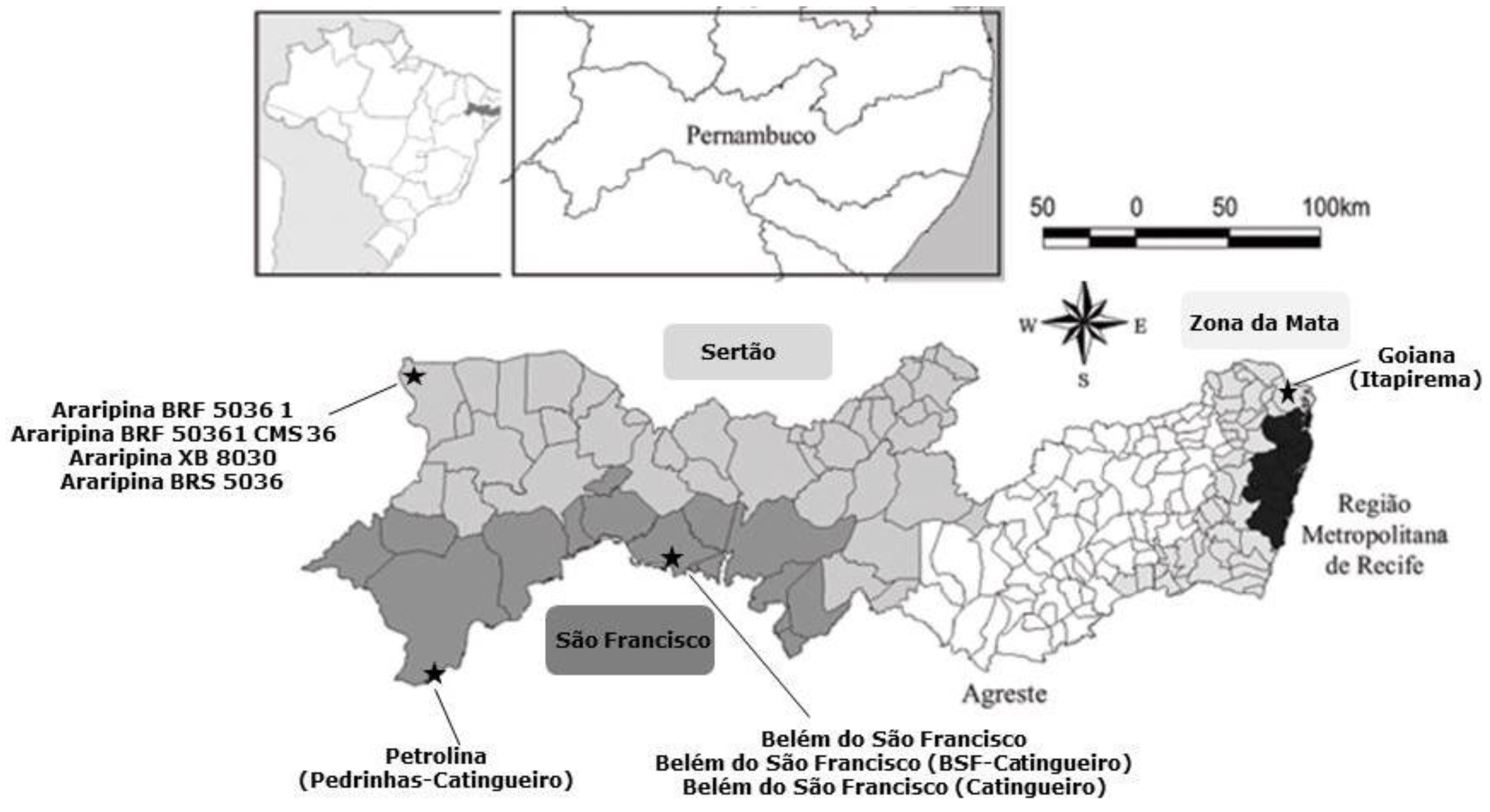
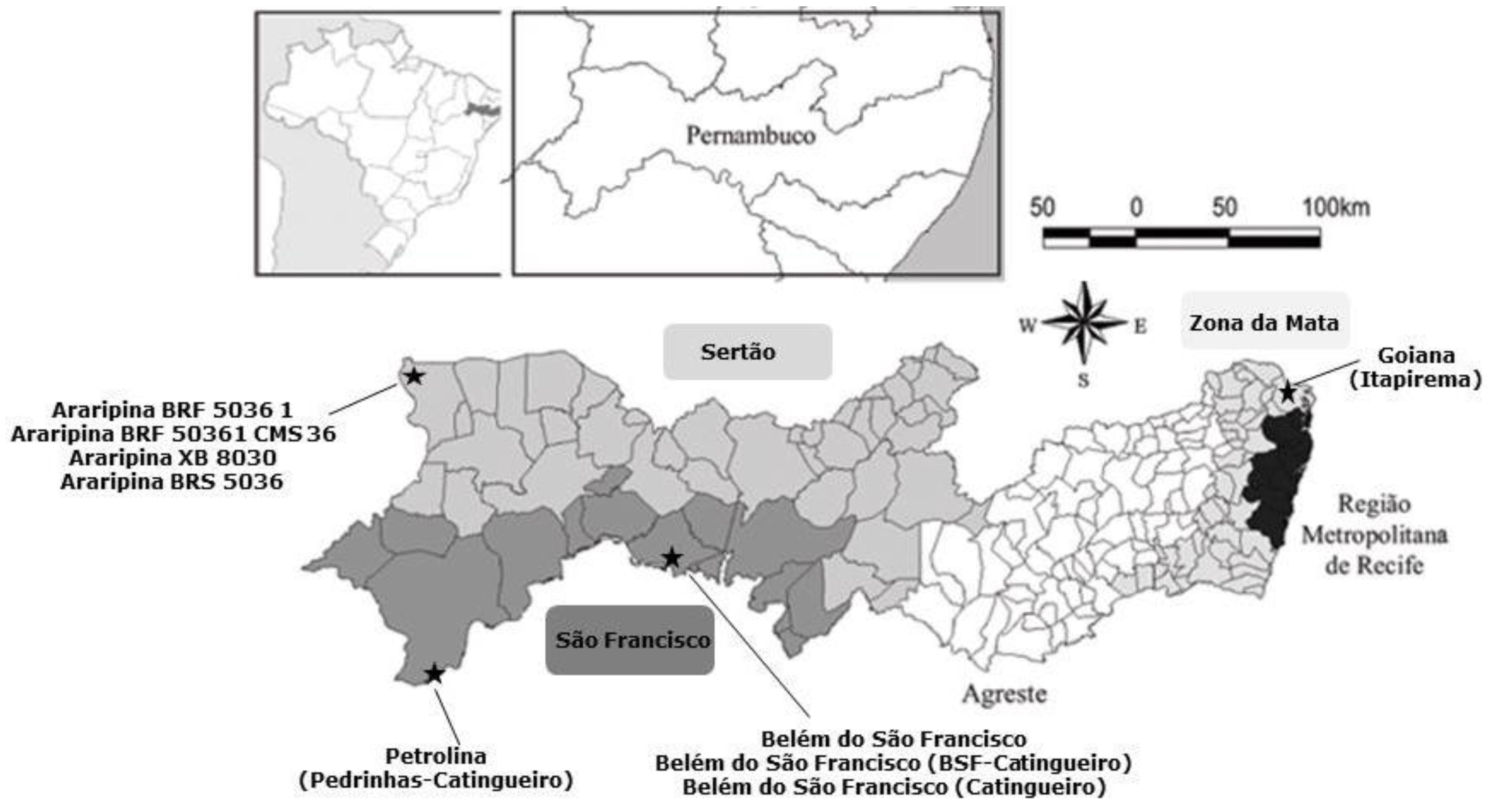
| Strains (Code N0: From 1 to 60) | Place | Region |
|---|---|---|
| 1 to 17 | Goiana (Itapirema) | Zona da Mata |
| 18 | Petrolina (Pedrinhas-Catingueiro) | São Francisco |
| 19 to 30 | Araripina XB 8030 | Sertão |
| 31 to 39 | Araripina BRF 5036-1 | Sertão |
| 40 to 45 | Araripina BRF 50361 CMS 36 | Sertão |
| 46 to 52 | Araripina BRS 5036 | Sertão |
| 53 to 60 | Belém do São Francisco | São Francisco |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, S.; Porto Carneiro-Leão, M.; Ferreira de Oliveira, B.; Souza-Motta, C.; Lima, N.; Santos, C.; Tinti de Oliveira, N. Polyphasic Approach Including MALDI-TOF MS/MS Analysis for Identification and Characterisation of Fusarium verticillioides in Brazilian Corn Kernels. Toxins 2016, 8, 54. https://doi.org/10.3390/toxins8030054
Chang S, Porto Carneiro-Leão M, Ferreira de Oliveira B, Souza-Motta C, Lima N, Santos C, Tinti de Oliveira N. Polyphasic Approach Including MALDI-TOF MS/MS Analysis for Identification and Characterisation of Fusarium verticillioides in Brazilian Corn Kernels. Toxins. 2016; 8(3):54. https://doi.org/10.3390/toxins8030054
Chicago/Turabian StyleChang, Susane, Mariele Porto Carneiro-Leão, Benny Ferreira de Oliveira, Cristina Souza-Motta, Nelson Lima, Cledir Santos, and Neiva Tinti de Oliveira. 2016. "Polyphasic Approach Including MALDI-TOF MS/MS Analysis for Identification and Characterisation of Fusarium verticillioides in Brazilian Corn Kernels" Toxins 8, no. 3: 54. https://doi.org/10.3390/toxins8030054
APA StyleChang, S., Porto Carneiro-Leão, M., Ferreira de Oliveira, B., Souza-Motta, C., Lima, N., Santos, C., & Tinti de Oliveira, N. (2016). Polyphasic Approach Including MALDI-TOF MS/MS Analysis for Identification and Characterisation of Fusarium verticillioides in Brazilian Corn Kernels. Toxins, 8(3), 54. https://doi.org/10.3390/toxins8030054