
Fast Screening of Antibacterial Compounds from Fusaria


Abstract
:1. Introduction
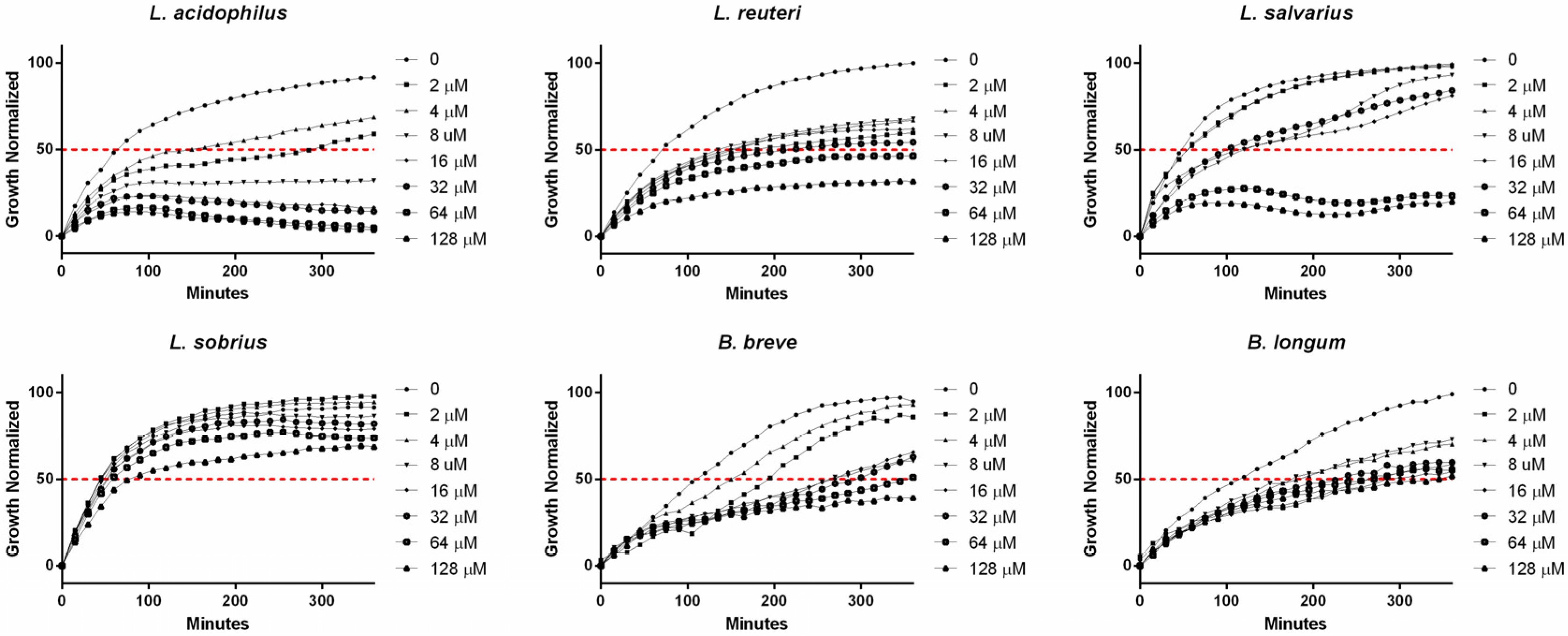
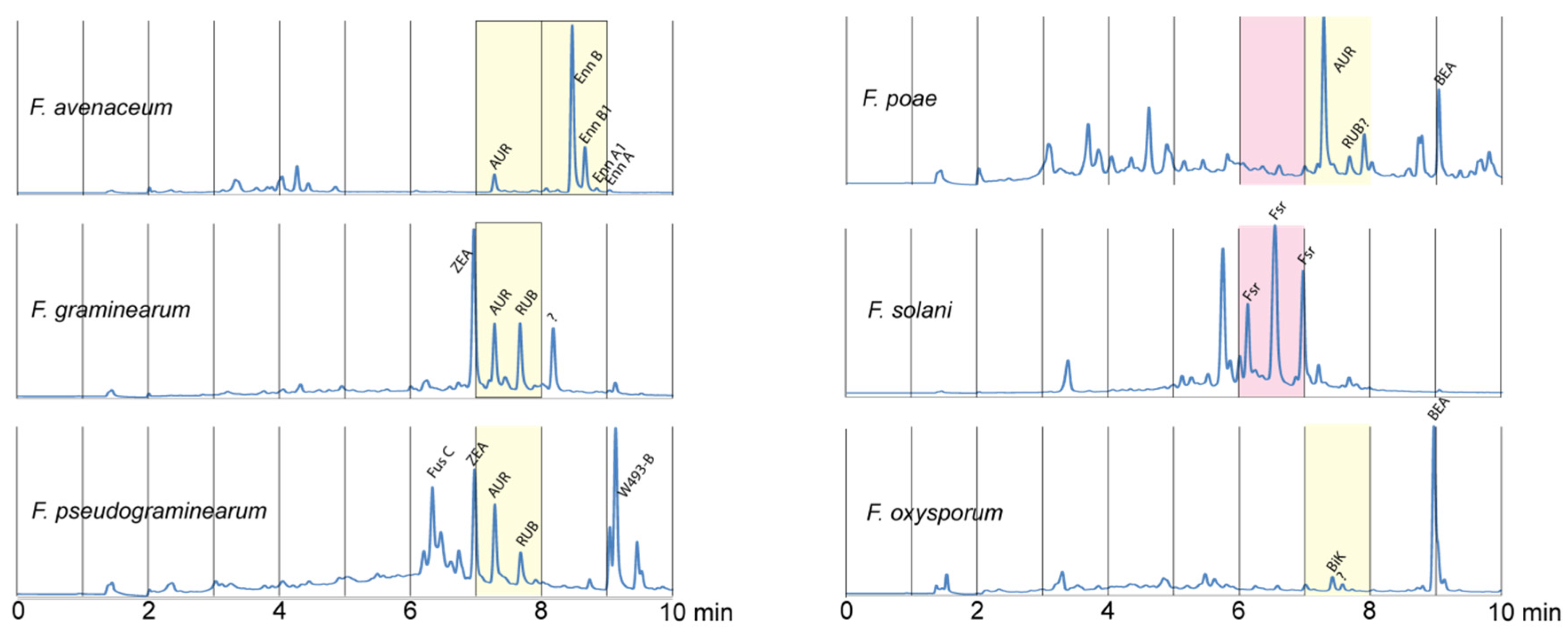
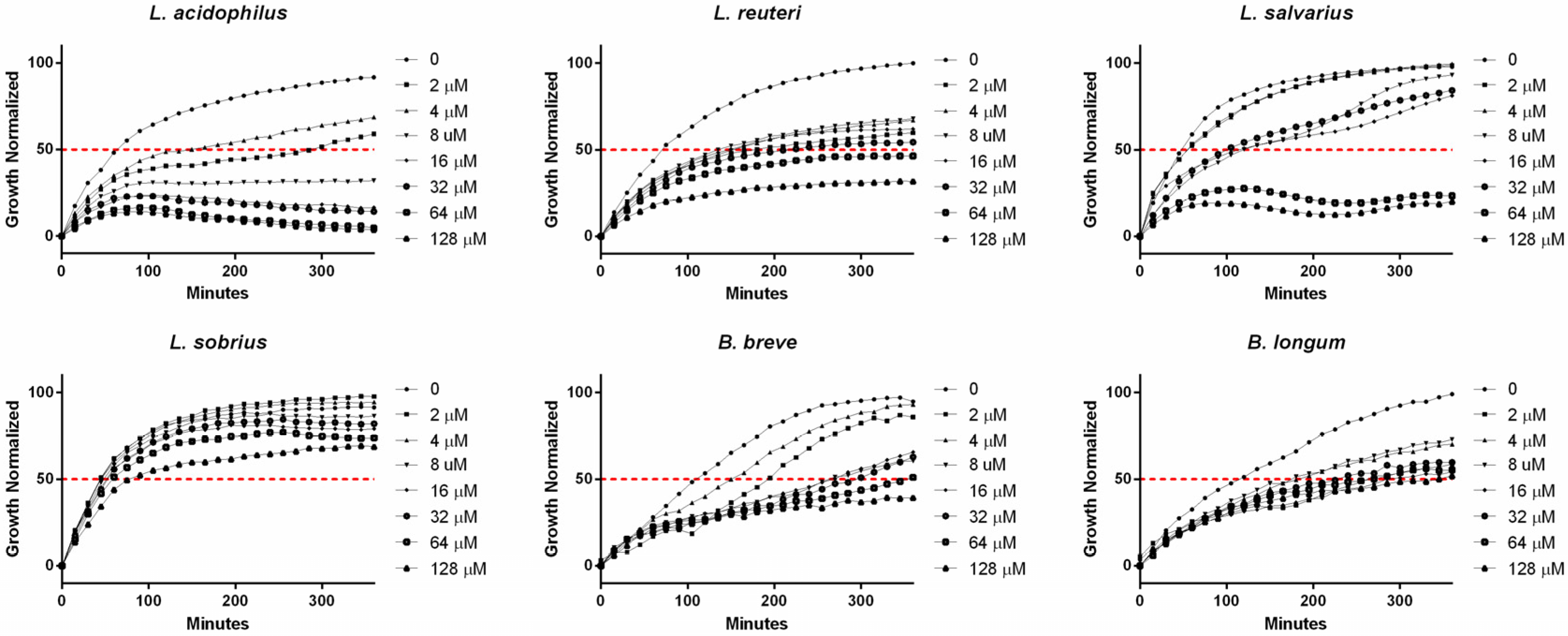
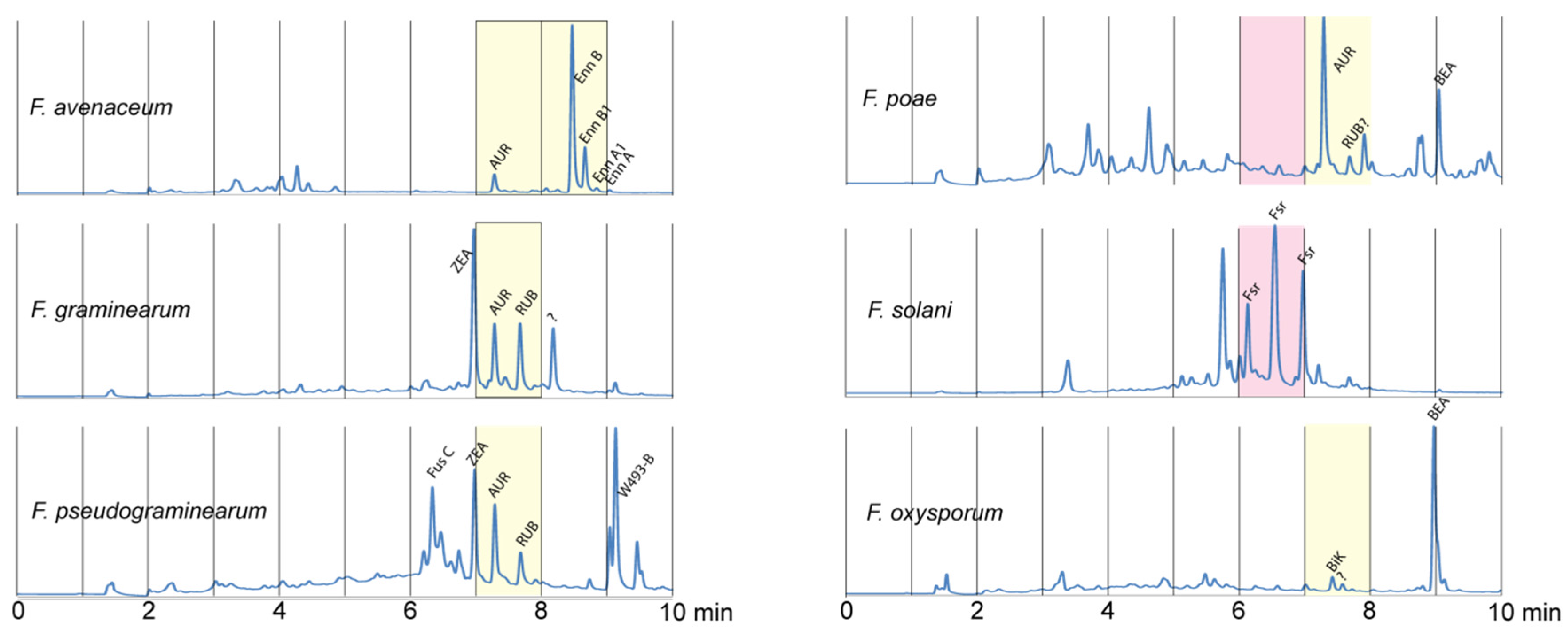
2. Results and Discussion
3. Conclusions
4. Materials and Methods
4.1. Secondary Metabolites
4.2. Bacterial Strains
4.3. Fungal Strains
4.4. Culture Media
4.5. Secondary Metabolite Fractionation
4.6. Antibiotic Susceptibility Tests
4.7. Digital Analysis
4.8. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fleming, A. On the antibacterial action of cultures of a Penicillium, with special reference to their use in the isolation of B. influenzae. Brit. J. Exp. Pathol. 1929, 10, 226–236. [Google Scholar] [CrossRef]
- Bugni, T.S.; Harper, M.K.; McCulloch, M.W.B.; Reppart, J.; Ireland, C.M. Fractionated marine invertebrate extract libraries for drug discovery. Molecules 2008, 13, 1372–1383. [Google Scholar] [CrossRef] [PubMed]
- Lang, G.; Mayhudin, N.A.; Mitova, M.I.; Sun, L.; van der Sar, S.; Blunt, J.W.; Cole, A.L.J.; Ellis, G.; Laatsch, H.; Munro, M.H.G. Evolving trends in the dereplication of natural product extracts: New methodology for rapid, small-scale investigation of natural product extracts. J. Nat. Prod. 2008, 71, 1595–1599. [Google Scholar] [CrossRef] [PubMed]
- Månsson, M.; Phipps, R.K.; Gram, L.; Munro, M.H.G.; Larsen, T.O.; Nielsen, K.F. Explorative solid-phase extraction (E-SPE) for accelerated microbial natural product discovery, dereplication, and purification. J. Nat. Prod. 2010, 73, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Bladt, T.T.; Durr, C.; Knudsen, P.B.; Kildgaard, S.; Frisvad, J.C.; Gotfredsen, C.H.; Seiffert, M.; Larsen, T.O. Bio-activity and dereplication-based discovery of ophiobolins and other fungal secondary metabolites targeting leukemia cells. Molecules 2013, 18, 14629–14650. [Google Scholar] [CrossRef] [PubMed]
- Fredborg, M.; Andersen, K.R.; Jørgensen, E.; Droce, A.; Olesen, T.; Jensen, B.B.; Rosenvinge, F.S.; Sondergaard, T.E. Real-time optical antimicrobial susceptibility testing. J. Clin. Microbiol. 2013, 51, 2047–2053. [Google Scholar] [CrossRef] [PubMed]
- Hansen, F.T.; Gardiner, D.M.; Lysøe, E.; Fuertes, P.R.; Tudzynski, B.; Wiemann, P.; Sondergaard, T.E.; Giese, H.; Brodersen, D.E.; Sørensen, J.L. An update to polyketide synthase and non-ribosomal synthetase genes and nomenclature in Fusarium. Fungal Genet. Biol. 2015, 75, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, G.W.; Li, Y.L.; Poulton, G.A. Pyrones. X. Lateropyrone, a new antibiotic from the fungus Fusarium lateritium Nees. Can. J. Chem. 1984, 62, 2101–2106. [Google Scholar] [CrossRef]
- Castlebury, L.A.; Sutherland, J.B.; Tanner, L.A.; Henderson, A.L.; Cerniglia, C.E. Use of a bioassay to evaluate the toxicity of beauvericin to bacteria. World J. Microbiol. Biotech. 1999, 15, 131–133. [Google Scholar] [CrossRef]
- Golinski, P.; Wnuk, S.; Chelkowski, J.; Visconti, A.; Schollenberger, M. Antibiotic Y: Biosynthesis by Fusarium avenaceum (Corda ex Fries) Sacc., isolation, and some physicochemical and biological properties. Appl. Environ. Microbiol. 1986, 51, 743–745. [Google Scholar] [PubMed]
- Hamill, R.L.; Higgens, C.E.; Boaz, H.E.; Gorman, M. The structure of beauvericin, a new depsipeptide antibiotic toxic to Artemia salina. Tetrahedron Lett. 1969, 4255–4258. [Google Scholar] [CrossRef]
- Meca, G.; Sospedra, I.; Adela Valero, M.; Manes, J.; Font, G.; Jose Ruiz, M. Antibacterial activity of the enniatin B, produced by Fusarium tricinctum in liquid culture, and cytotoxic effects on Caco-2 cells. Toxicol. Mech. Meth. 2011, 21, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Meca, G.; Sospedra, I.; Soriano, J.M.; Ritieni, A.; Moretti, A.; Manes, J. Antibacterial effect of the bioactive compound beauvericin produced by Fusarium proliferatum on solid medium of wheat. Toxicon 2010, 56, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Nilanonta, C.; Isaka, M.; Chanphen, R.; Thong-Orn, N.; Tanticharoen, M.; Thebtaranonth, Y. Unusual enniatins produced by the insect pathogenic fungus Verticillium hemipterigenum: Isolation and studies on precursor-directed biosynthesis. Tetrahedron 2003, 59, 1015–1020. [Google Scholar] [CrossRef]
- Nilanonta, C.; Isaka, M.; Kittakoop, P.; Palittapongarnpim, P.; Kamchonwongpaisan, S.; Pittayakhajonwut, D.; Tanticharoen, M.; Thebtaranonth, Y. Antimycobacterial and antiplasmodial cyclodepsipeptides from the insect pathogenic fungus Paecilomyces tenuipes BCC 1614. Planta Medica 2000, 66, 756–758. [Google Scholar] [CrossRef] [PubMed]
- Sebastia, N.; Meca, G.; Miguel Soriano, J.; Manes, J. Antibacterial effects of enniatins J1 and J3 on pathogenic and lactic acid bacteria. Food Chem. Toxicol. 2011, 49, 2710–2717. [Google Scholar] [CrossRef] [PubMed]
- Shenyakin, M.M.; Ovchinnikov, Y.A.; Ivanov, V.T.; Evstratov, A.V. Topochemical approach in studies of the structure-activity relation: Enantio-enniatin B. Nature 1967, 213, 412–413. [Google Scholar] [CrossRef] [PubMed]
- Tomoda, H.; Nishida, H.; Huang, X.H.; Masuma, R.; Kim, Y.K.; Omura, S. New cyclodepsipeptides, enniatins D, E and F produced by Fusarium sp. FO-1305. J. Antibiot. 1992, 45, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Son, S.W.; Kim, H.Y.; Choi, G.J.; Lim, H.K.; Jang, K.S.; Lee, S.O.; Lee, S.; Sung, N.D.; Kim, J.C. Bikaverin and fusaric acid from Fusarium oxysporum show antioomycete activity against Phytophthora infestans. J. Appl. Microbiol. 2008, 104, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Supothina, S.; Isaka, M.; Kirtikara, K.; Tanticharoen, M.; Thebtaranonth, Y. Enniatin production by the entomopathogenic fungus Verticillium hemipterigenum BCC 1449. J. Antibiot. 2004, 57, 732–738. [Google Scholar] [CrossRef] [PubMed]
- May, H.D.; Wu, Q.Z.; Blake, C.K. Effects of the Fusarium spp. mycotoxins fusaric acid and deoxynivalenol on the growth of Ruminococcus albus and Methanobrevibacter ruminantium. Can. J. Microbiol. 2000, 46, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Vesonder, R.F.; Ellis, J.J.; Rohwedder, W.K. Elaboration of vomitoxin and zearalenone by Fusarium isolates and the biological activity of Fusarium-produced toxins. Appl. Environ. Microbiol. 1981, 42, 1132–1134. [Google Scholar] [PubMed]
- Szkaradkiewicz, A.; Wal, M. Effect of cyclosporin on uropathogenic Escherichia coli adherence to human endothelial cells. Int. J. Antimicrob. Agents 2001, 18, 89–91. [Google Scholar] [CrossRef]
- O’Donnell, K.; Rooney, A.P.; Proctor, R.H.; Brown, D.W.; McCormick, S.P.; Ward, T.J.; Frandsen, R.J.N.; Lysøe, E.; Rehner, S.A.; Aoki, T.; et al. Phylogenetic analyses of RPB1 and RPB2 support a middle Cretaceous origin for a clade comprising all agriculturally and medically important fusaria. Fungal Genet. Biol. 2013, 52, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Arnstein, H.R.V.; Cook, A.H.; Lacey, M.S. Production of antibiotics by fungi. Part II: Production by Fusarium javanicum and other fusaria. Br. J. Exp. Pathol. 1946, 27, 349–355. [Google Scholar] [PubMed]
- Dvorska, J. Effects of aurofusarin, a dimeric naphthoquinone metabolite of Fusarium graminearum, on fatty acid profiles and antioxidant composition of quail eggs. Toxicology 2001, 164, 176. [Google Scholar]
- Dvorska, J.E.; Surai, P.F.; Speake, B.K.; Sparks, N.H.C. Effect of the mycotoxin aurofusarin on the antioxidant composition and fatty acid profile of quail eggs. Br. Poult. Sci. 2001, 42, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, J.L.; Nielsen, K.F.; Sondergaard, T.E. Redirection of pigment biosynthesis to isocoumarins in Fusarium. Fungal Genet. Biol. 2012, 49, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Busman, M.; Butchko, R.A.E.; Proctor, R.H. LC-MS/MS method for the determination of the fungal pigment bikaverin in maize kernels as an indicator of ear rot. Food Addit. Contam. Part A-Chem. 2012, 29, 1736–1742. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, S.; Eriksen, G.S.; Hofgaard, I.S.; Krska, R.; Beltran, E.; Sulyok, M. Faces of a changing climate: Semi-quantitative multi-mycotoxin analysis of grain grown in exceptional climatic conditions in Norway. Toxins 2013, 5, 1682–1697. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, J.L.; Phipps, R.K.; Nielsen, K.F.; Schroers, H.J.; Frank, J.; Thrane, U. Analysis of Fusarium avenaceum metabolites produced during wet apple core rot. J. Agric. Food Chem. 2009, 57, 1632–1639. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.; Mathew, A.; Purohit, H.J. Characterization of antibacterial activity of bikaverin from Fusarium sp. HKF15. J. Biosci. Bioeng. 2014, 117, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Medentsev, A.G.; Akimenko, V.K. Naphthoquinone metabolites of the fungi. Phytochemistry 1998, 47, 935–959. [Google Scholar] [CrossRef]
- Baker, R.A.; Tatum, J.H.; Nemec, S., Jr. Antimicrobial activity of naphthoquinones from Fusaria. Mycopathologia 1990, 111, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Brown, M. Modes of action of probiotics: Recent developments. J. Anim. Vet. Adv. 2011, 10, 1895–1900. [Google Scholar] [CrossRef]
- Ohashi, Y.; Ushida, K. Health-beneficial effects of probiotics: Its mode of action. Anim. Sci. J. 2009, 80, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Fairbrother, J.M.; Nadeau, E.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Sunaga, R.; Furihata, K.; Morisaki, N.; Iwasaki, S. Isolation and structures of an antifungal antibiotic, Fusarielin A, and related compounds produced by a Fusarium sp. J. Antibiot. 1995, 48, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, J.L.; Hansen, F.T.; Sondergaard, T.E.; Staerk, D.; Lee, T.V.; Wimmer, R.; Klitgaard, L.G.; Purup, S.; Giese, H.; Frandsen, R.J. Production of novel fusarielins by ectopic activation of the polyketide synthase 9 cluster in Fusarium graminearum. Environ. Microbiol. 2012, 14, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Sondergaard, T.E.; Hansen, F.T.; Purup, S.; Nielsen, A.K.; Bonefeld-Jorgensen, E.C.; Giese, H.; Sørensen, J.L. Fusarin C acts like an estrogenic agonist and stimulates breast cancer cells in vitro. Toxicol. Lett. 2011, 205, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Holdeman, L.V.; Cato, E.P.; Moore, W.E.C. Anaerobe Laboratory Manual, 4th ed.; Virginia Polytechnic Institute and State University: Blacksburg, VA, USA, 1977. [Google Scholar]
- Frisvad, J.C. Media and growth conditions for induction of secondary metabolite production. Meth. Mol. Biol. 2012, 944, 47–58. [Google Scholar]
- Fredborg, M.; Rosenvinge, F.S.; Spillum, E.; Kroghsbo, S.; Wang, M.; Sondergaard, T.E. Automated image analysis for quantification of filamentous bacteria. BMC Microbiol. 2015, 15, 255. [Google Scholar] [CrossRef] [PubMed]
- Uggerhoj, L.E.; Poulsen, T.J.; Munk, J.K.; Fredborg, M.; Sondergaard, T.E.; Frimodt-Moller, N.; Hansen, P.R.; Wimmer, R. Rational design of alpha-helical antimicrobial peptides: Do’s and don’ts. ChemBioChem 2015, 16, 242–253. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| L. acidophilus | E. coli | Staph. aureus | Salm. typhimurium | |
|---|---|---|---|---|
| Antibiotic Y | - | - | 32 μM | - |
| Beauvericin | 32 μM | - | 32 μM | - |
| Enniatin mix | 32 μM | - | 64 μM | 128 μM |
| Fusaric acid | 64 μM | 64 μM | - | - |
| Aurofusarin | 8 μM | - | - | - |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sondergaard, T.E.; Fredborg, M.; Oppenhagen Christensen, A.-M.; Damsgaard, S.K.; Kramer, N.F.; Giese, H.; Sørensen, J.L. Fast Screening of Antibacterial Compounds from Fusaria. Toxins 2016, 8, 355. https://doi.org/10.3390/toxins8120355
Sondergaard TE, Fredborg M, Oppenhagen Christensen A-M, Damsgaard SK, Kramer NF, Giese H, Sørensen JL. Fast Screening of Antibacterial Compounds from Fusaria. Toxins. 2016; 8(12):355. https://doi.org/10.3390/toxins8120355
Chicago/Turabian StyleSondergaard, Teis Esben, Marlene Fredborg, Ann-Maria Oppenhagen Christensen, Sofie K. Damsgaard, Nikoline F. Kramer, Henriette Giese, and Jens Laurids Sørensen. 2016. "Fast Screening of Antibacterial Compounds from Fusaria" Toxins 8, no. 12: 355. https://doi.org/10.3390/toxins8120355
APA StyleSondergaard, T. E., Fredborg, M., Oppenhagen Christensen, A.-M., Damsgaard, S. K., Kramer, N. F., Giese, H., & Sørensen, J. L. (2016). Fast Screening of Antibacterial Compounds from Fusaria. Toxins, 8(12), 355. https://doi.org/10.3390/toxins8120355