The Fate of Microcystins in the Environment and Challenges for Monitoring


Abstract
:1. Introduction
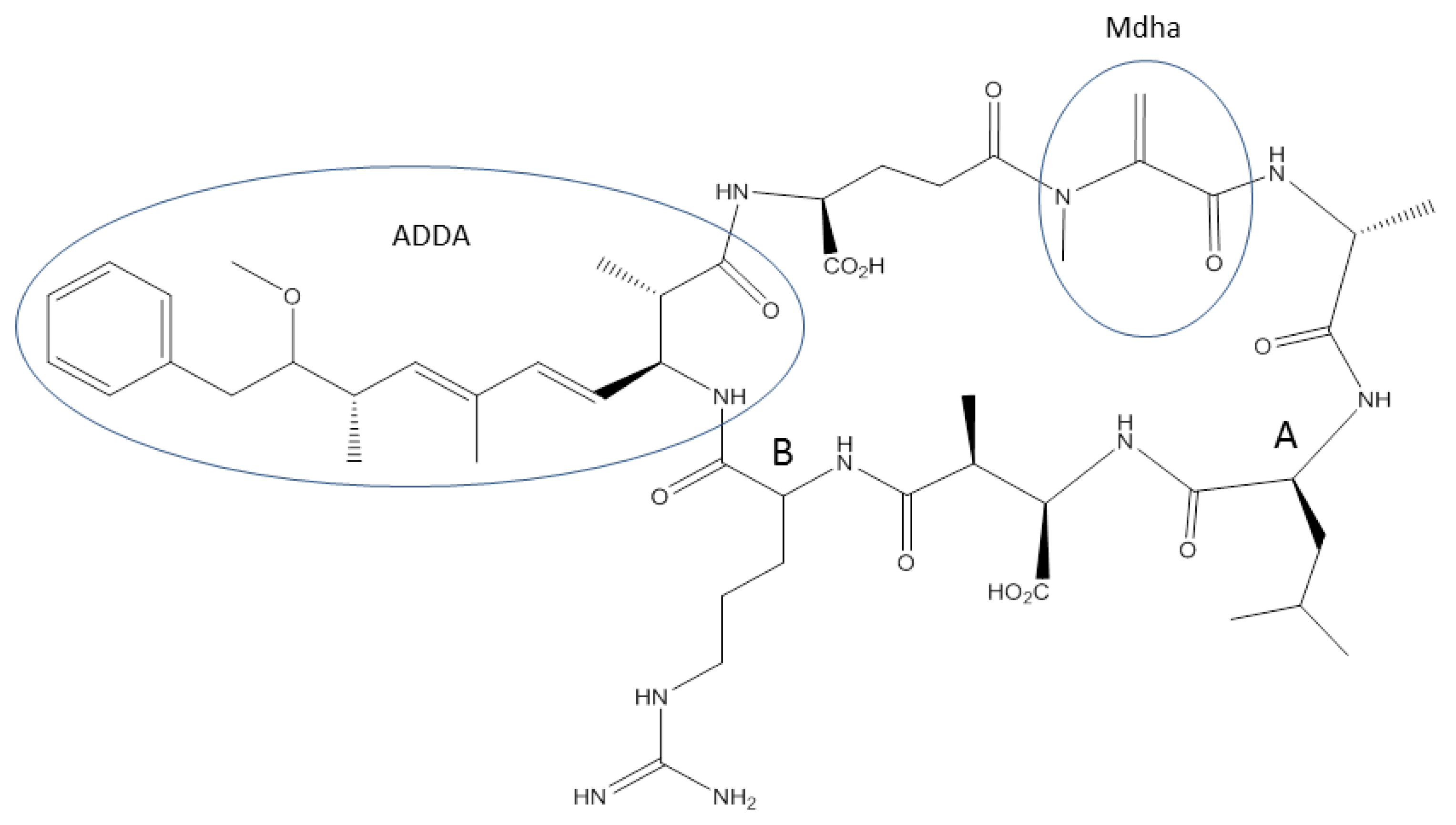
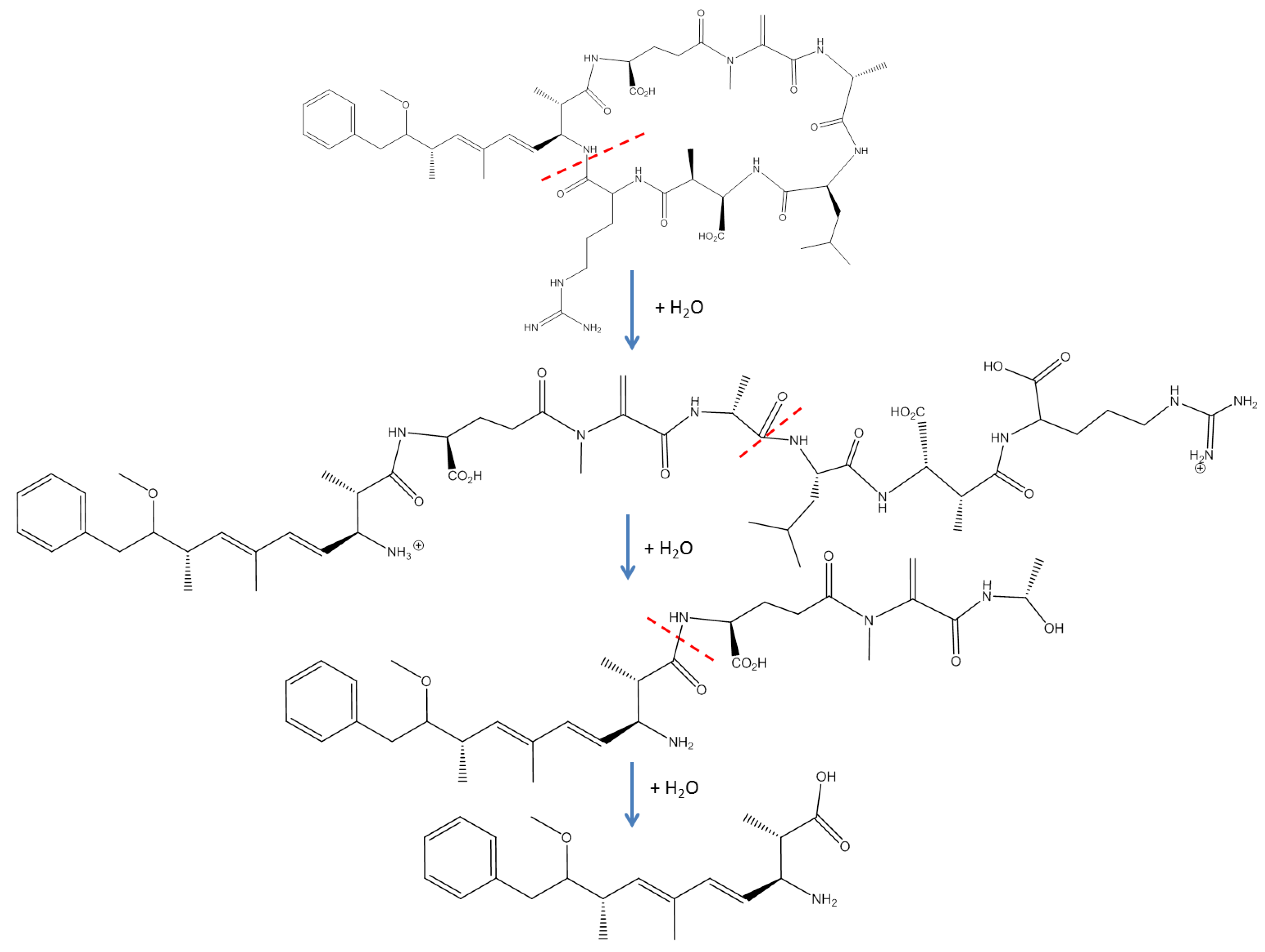
1.1. Abiotic Transformations
1.2. Biological Processes
1.2.1. Microbial Degradation
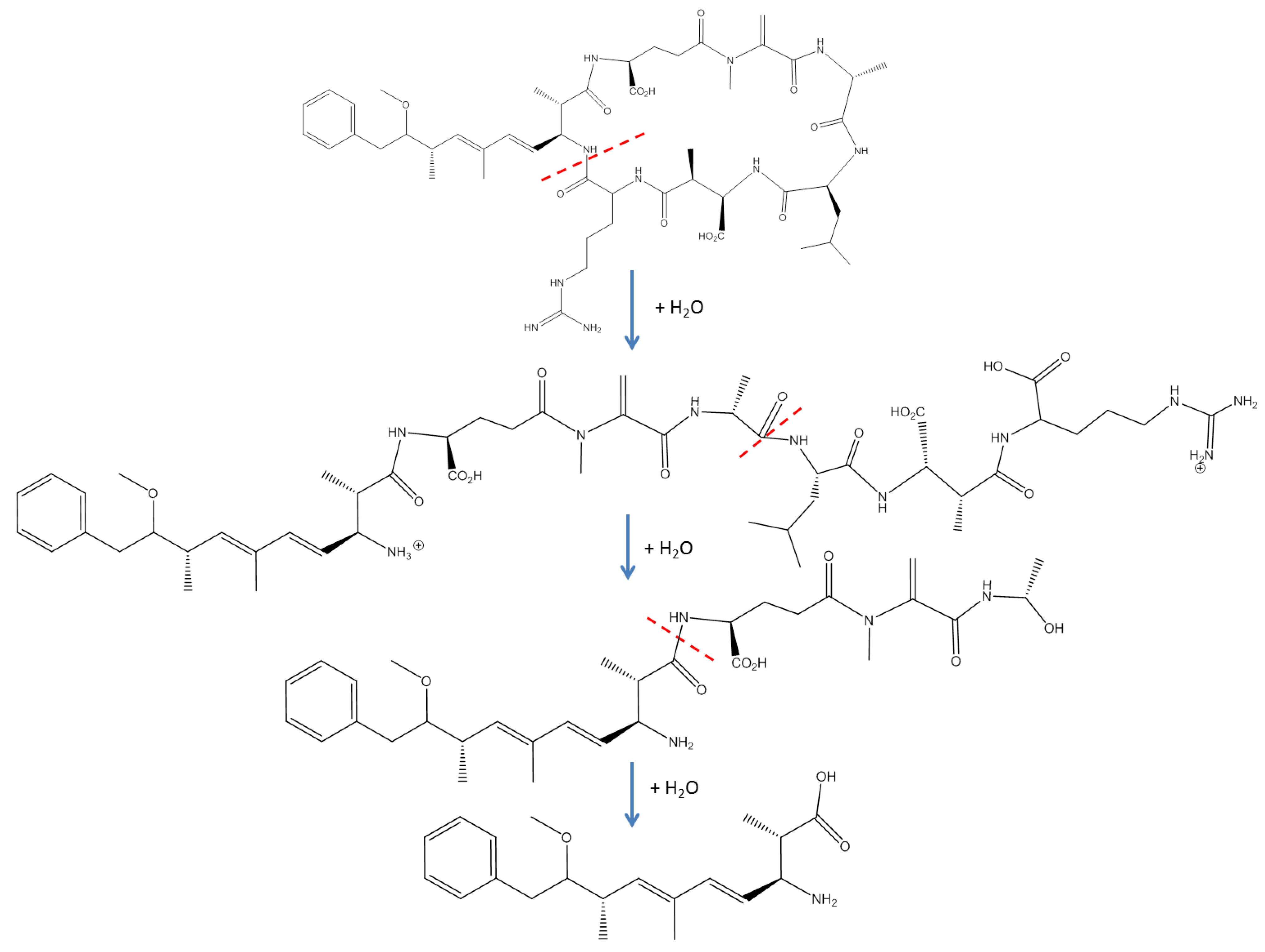
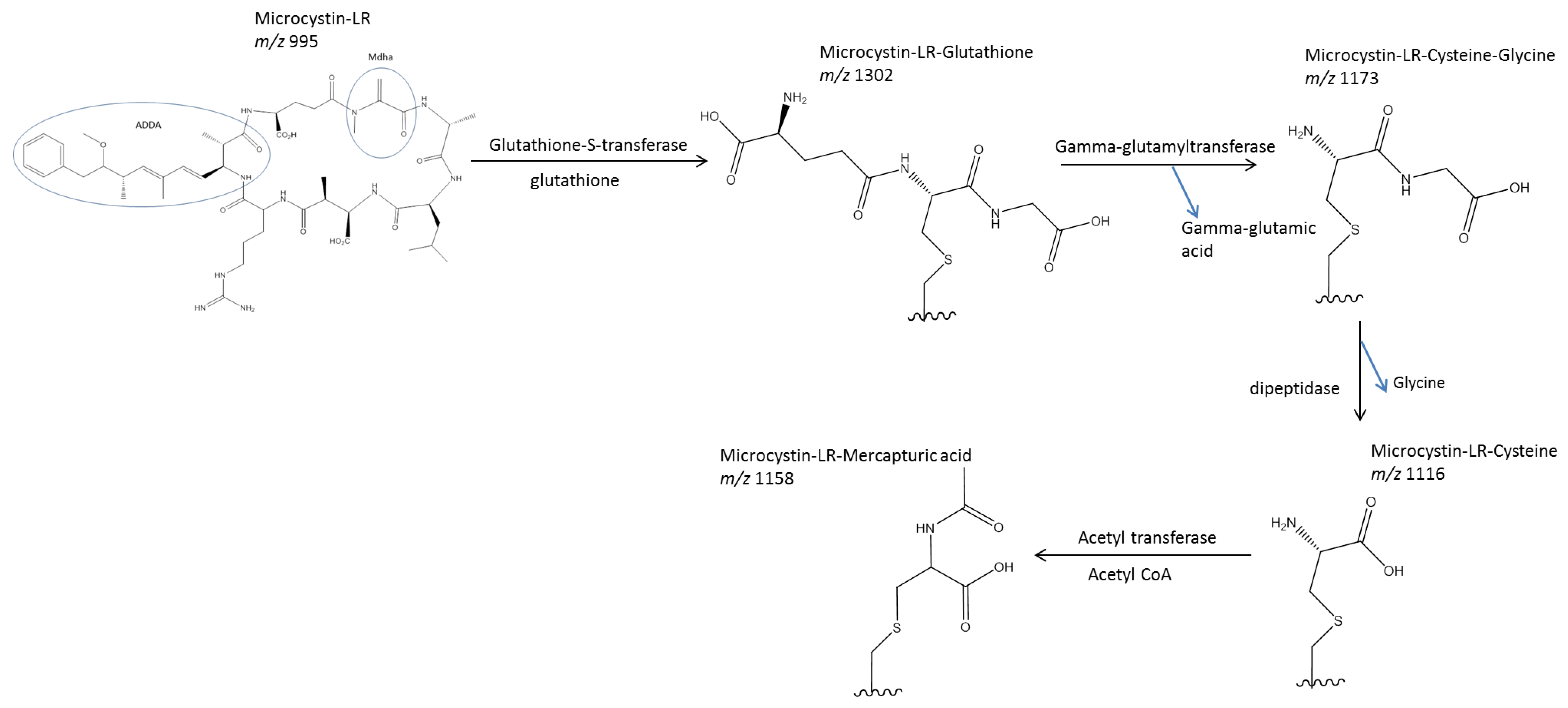
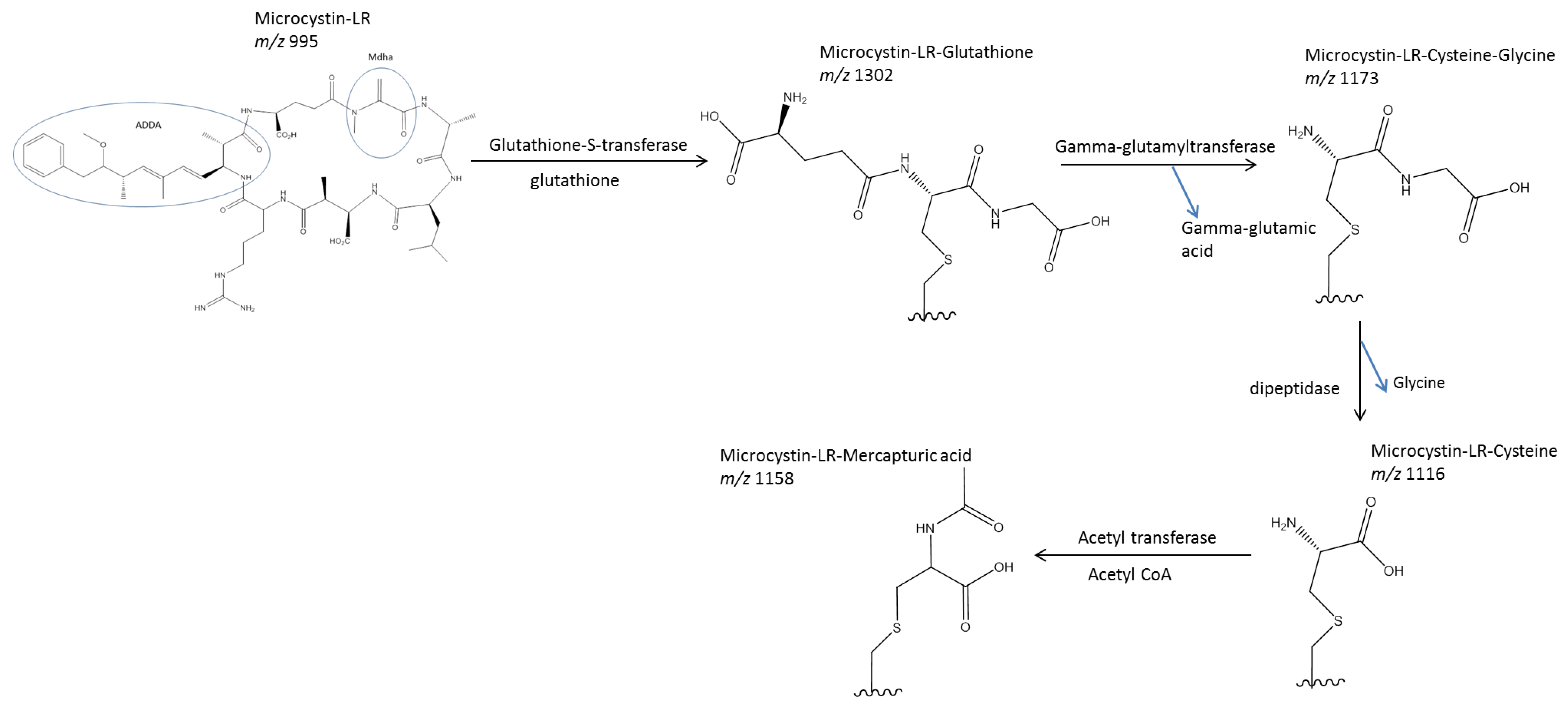
1.2.2. Metabolism and Conjugation
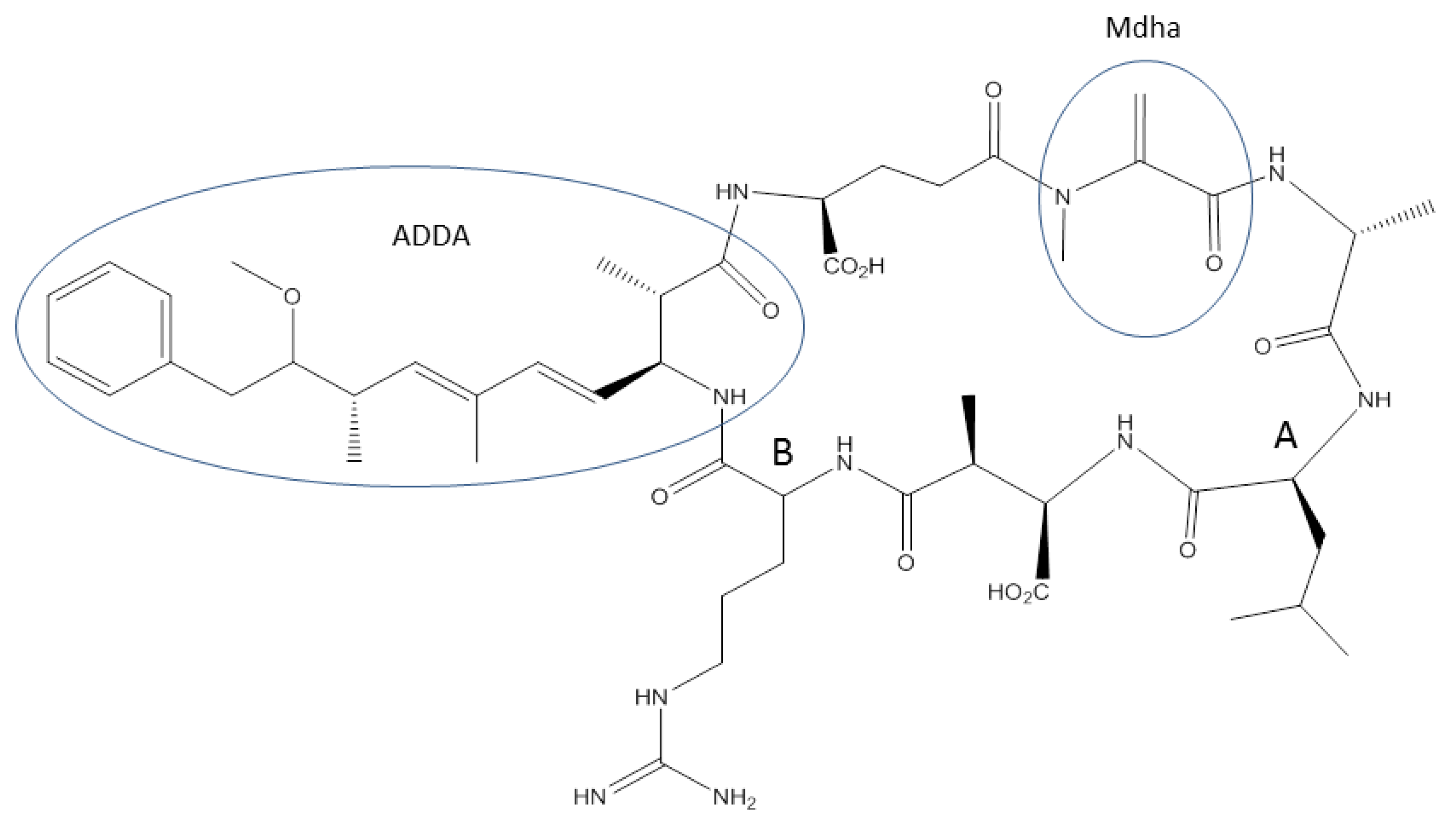
2. Degree of Resistance to Microcystin Toxicity
2.1. Terrestrial Organisms vs. Aquatic Organisms
2.2. Bivalves
2.3. Fish
2.4. Zooplankton
2.5. Detection of Microcystins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Year | Cleanup method | Analysis | Recovery of microcystin-LR |
|---|---|---|---|---|
| [172] | 2005 | C18 solid phase extraction | LC-MS | ~57% (estimated from Figure 3 of reference [172]) |
| [129] | 2005 | C18 solid phase extraction | LC-MS and ELISA | 68%–96% |
| [173] | 2005 | C18 solid phase extraction | LC-MS and ELISA | 44%–101% |
| [174] | 2007 | - | ELISA | >25% |
| [105] | 2007 | Waters Oasis solid phase extraction | LC-Photodiode array | >85% |
| [175] | 2008 | Waters Oasis solid phase extraction, Silica gel | LC-MS | >90% |
| [171] | 2009 | 500 mg C18 solid phase extraction | LC-MS/MS | 74%–125% |
| [170] | 2009 | - | LC-MS | 80%–99% |
| [114] | 2009 | - | LC-MS | 68%–73% |
| [95] | 2013 | Charcoal solid phase extraction | LC-MS/MS | 54%–106% |
3. Toxicokinetics
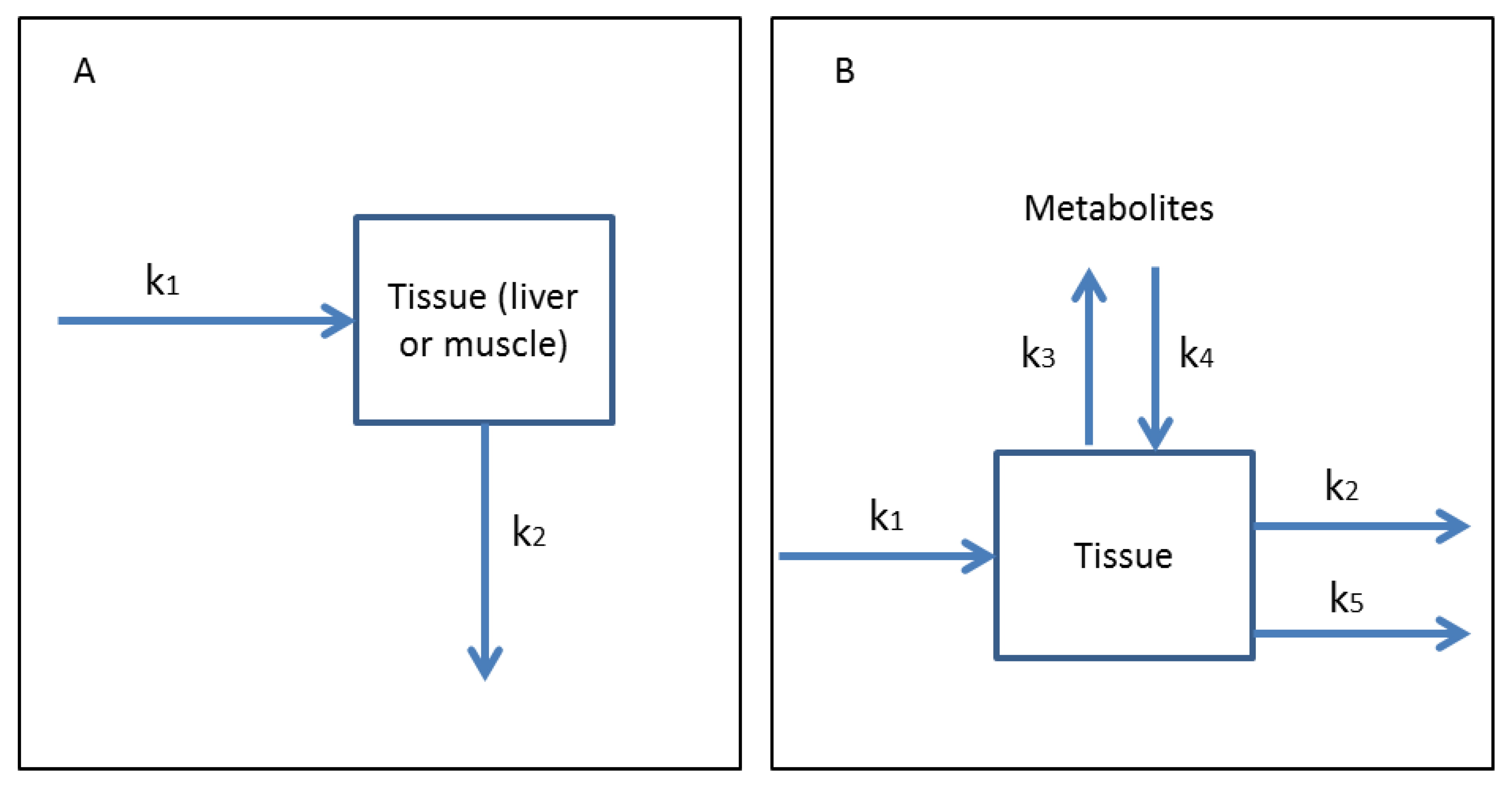
3.1. One-Compartment Models
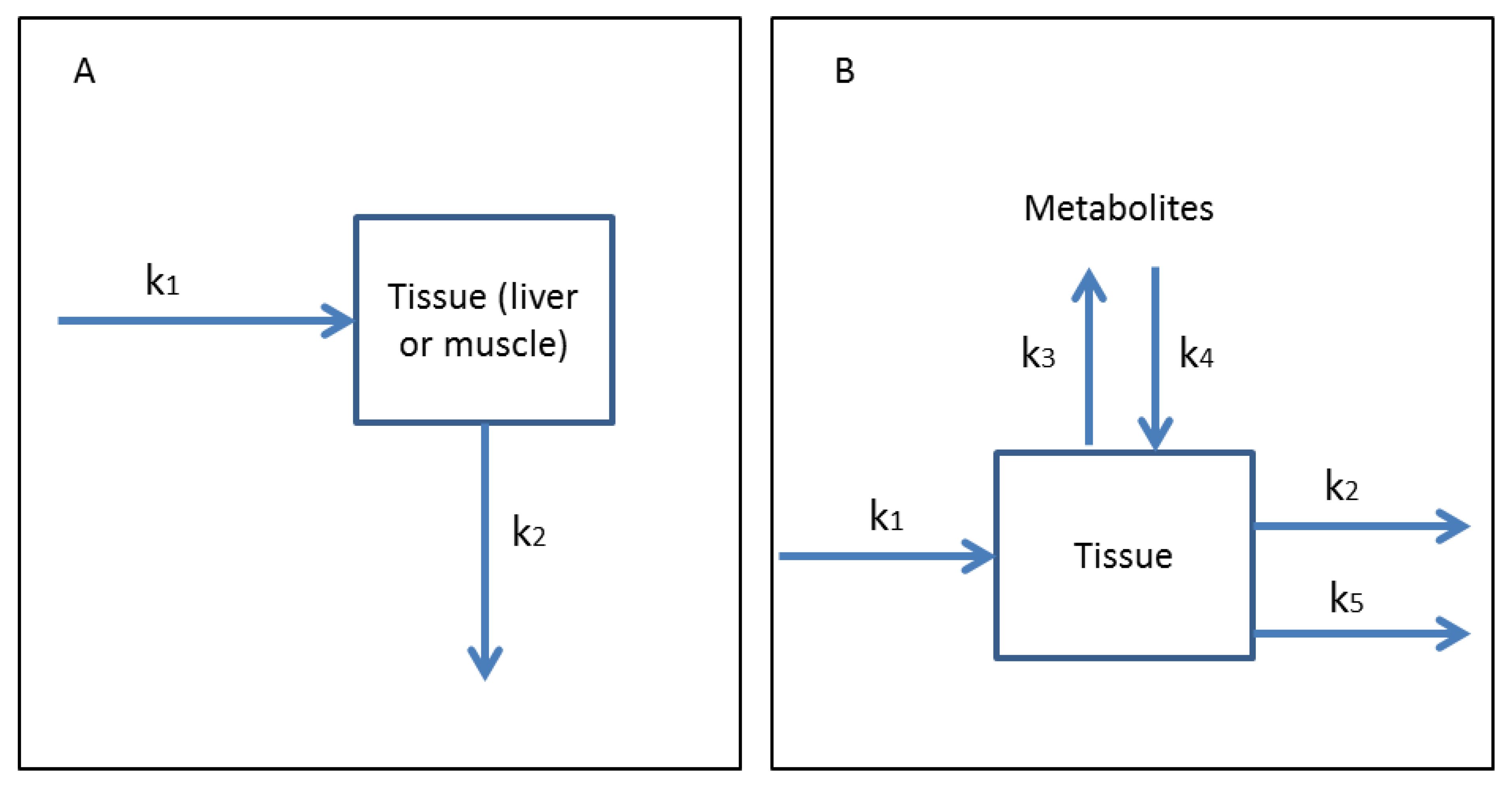
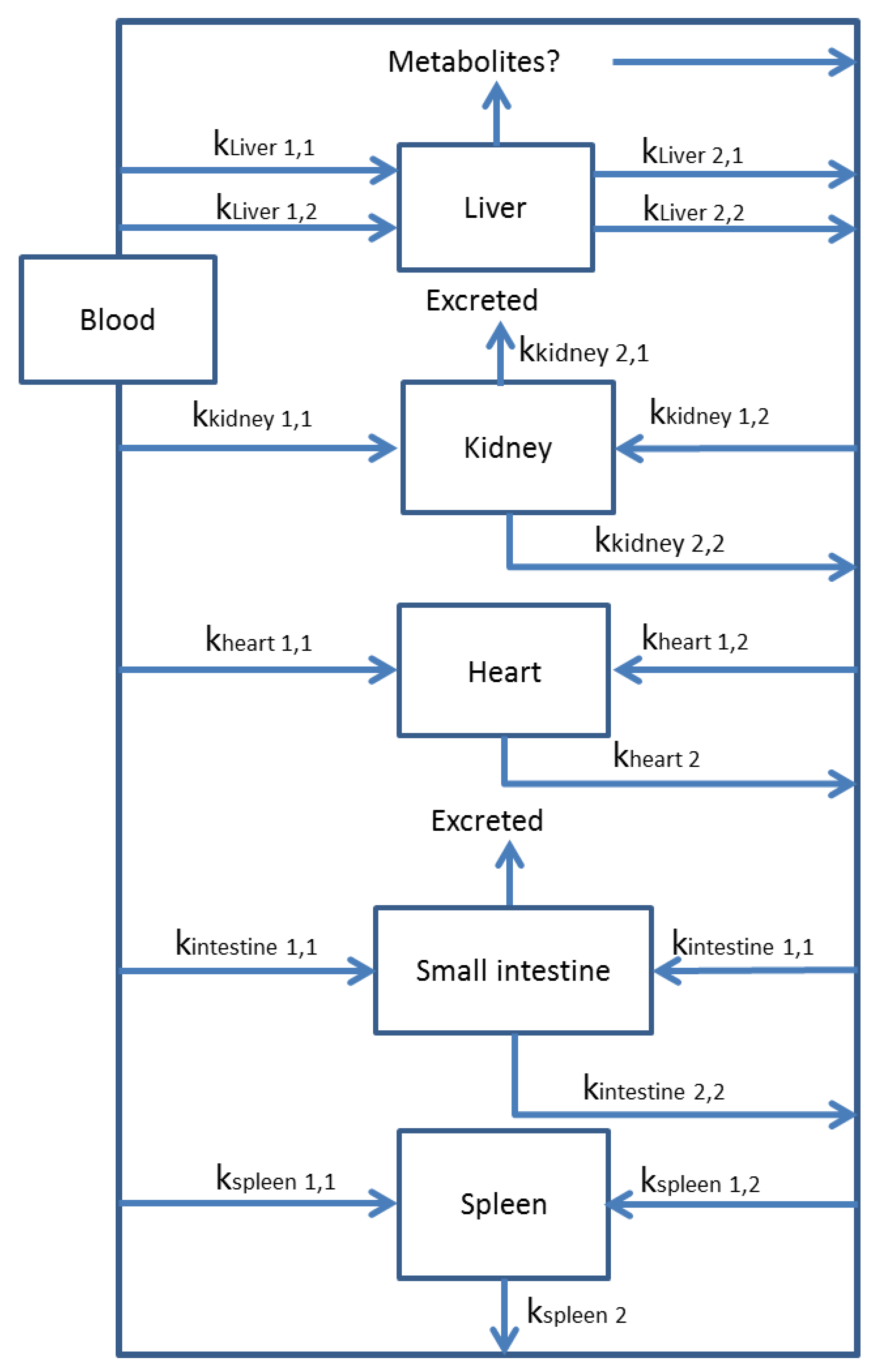
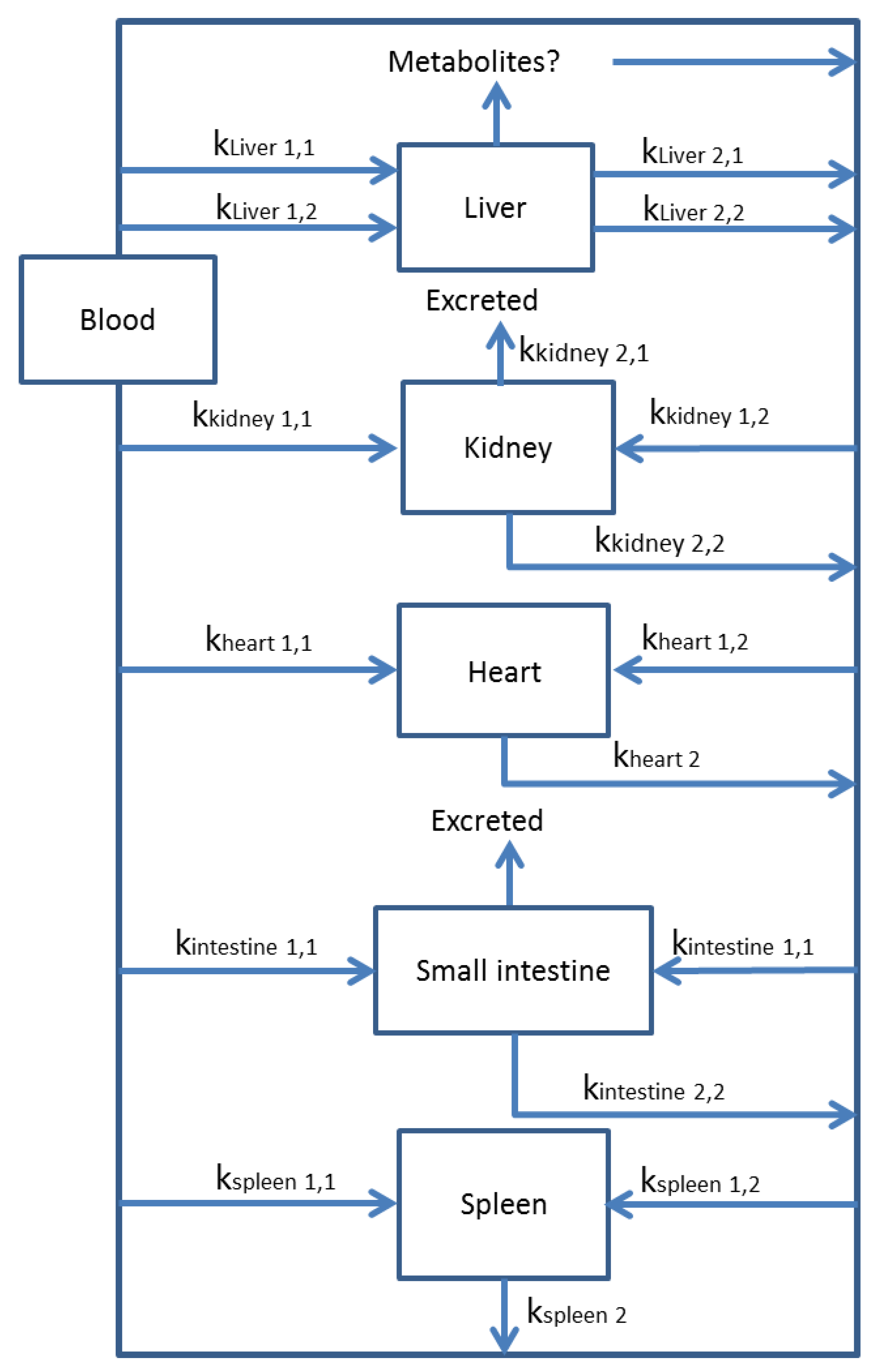
3.2. Multi-Compartmental Models
3.3. Microcystin Conjugate Toxicokinetics
4. Conclusions and Future Direction
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wehr, J.D.; Sheath, R.G. Freshwater habitats of algae. In Freshwater Algae of North America; Wehr, J.D., Sheath, R.G., Eds.; Academic Press: San Diego, CA, USA, 2003; pp. 11–57. [Google Scholar]
- Smith, J.L.; Boyer, G.L.; Zimba, P.V. A Review of Cyanobacterial Odorous and Bioactive Metabolites: Impacts and Management Alternatives in Aquaculture. Aquaculture 2008, 280, 5–20. [Google Scholar] [CrossRef]
- Lampert, W. Laboratory Studies on Zooplankton-Cyanobacteria Interactions. N. Z. J. Mar. Fresh. 1987, 21, 483–490. [Google Scholar] [CrossRef]
- Carmichael, W. Cyanobacteria Secondary Metabolites—The Cyanotoxins. J. Appl. Bacteriol. 1992, 72, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W. Health Effects of Toxin-Producing Cyanobacteria: “The CyanoHABs”. Hum. Ecol. Risk Assess. 2001, 7, 1393–1407. [Google Scholar] [CrossRef]
- Dittman, E.; Wiegand, C. Cyanobacterial Toxins—Occurrence, Biosynthesis and Impact on Human Affairs. Mol. Nutr. Food Res. 2006, 50, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B. On the Chemistry, Toxicology and Genetics of the Cyanobacterial Toxins, Microcystin, Nodularin, Saxitoxin and Cylindrospermopsin. Mar. Drugs 2010, 10, 1650–1680. [Google Scholar] [CrossRef]
- Newcombe, G.; Chorus, I.; Falconer, I.; Lin, T. Cyanobacteria: Impacts of Climate Change on Occurrence, Toxicity and Water Quality Management. Water Res. 2012, 46, 1347–1348. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Hall, N.S.; Calandrino, E.S. Controlling Harmful Cyanobacterial Blooms in a World Experiencing Anthropogenic and Climate-induced Change. Sci. Total Environ. 2011, 409, 1739–1745. [Google Scholar] [CrossRef] [PubMed]
- De Figueiredo, D.R.; Azeiteiro, U.M.; Esteves, S.M.; Gonçalves, F.J.M.; Pereira, M.J. Microcystin-producing blooms—A serious global public health issue. Ecotox. Environ. Saf. 2004, 59, 151–163. [Google Scholar] [CrossRef]
- Svrcek, C.; Smith, D.W. An Overview of the Cyanobacteria Toxins and the Current State of Knowledge on Water Treatment Options. J. Environ. Eng. 2004, 3, 155–185. [Google Scholar] [CrossRef]
- Zurawell, R.W.; Chen, H.; Burke, J.M.; Prepas, E.E. Hepatotoxic Cyanobacteria: A Review of the Biological Importance of Microcystins in Freshwater Environments. J. Toxicol. Environ. Health 2005, 8, 1–37. [Google Scholar] [CrossRef]
- Niedermeyer, T.H.J.; Daily, A.; Swiatecka-Hagenbruch, M.; Moscow, J.A. Selectivity and Potency of Microcystin Congeners against OATP1B1 and OATP1B3 Expressing Cancer Cells. PLoS One 2014, 9, e91476. [Google Scholar] [CrossRef] [PubMed]
- Runnegar, M.T.C.; Berndt, N.; Kaplowitz, N. Microcystin Uptake and Inhibition of Protein Phosphatases: Effects of Chemoprotectants and Self-inhibition in Relation to Known Hepatic Transporters. Toxicol. Appl. Pharmacol. 1995, 134, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Pant, S.C.; Vilayaraghavan, R.; Rao, P.V. Comparative Toxicity Evaluation of Cyanobacterial Cyclic Peptide Toxin Microcystin Variants (LR, RR, YR) in Mice. Toxicology 2003, 188, 285–296. [Google Scholar] [CrossRef] [PubMed]
- De Maagd, P.G.; Hendriks, A.J.; Seinen, W.; Sijm, D. pH-dependent Hydrophobicity of the Cyanobacteria Toxin Microcystin-LR. Water Res. 1999, 33, 677–680. [Google Scholar] [CrossRef]
- Eriksson, J.E.; Grönberg, L.; Nygård, S.; Slotte, P.; Meriluoto, J.A.O. Hepatocellular Uptake of 3H-dihydromicrocystin-LR, a Cyclic Peptide Toxin. Biochim. Biophys. Acta 1990, 1025, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Falconer, I.R. Cyanobacterial Toxins of Drinking Water Supplies; CRC Press: Boca Raton, FL, USA, 2005; pp. 1–9. [Google Scholar]
- Wolkoff, A.W.; Cohen, D.E. Hepatic Transport of Bile Acids. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, 175–179. [Google Scholar]
- MacKintosh, R.W.; Dalby, K.N.; Campbell, D.G.; Cohen, P.T.W.; Cohen, P.; MacKintosh, C. The Cyanobacterial Toxin Microcystin Binds Covalently to Cysteine-273 on Protein Phosphatase 1. FEBS Lett. 1995, 371, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Puerto, M.; Pichardo, S.; Jos, Á.; Cameán, A.M. Comparison of the Toxicity Induced by Microcystin-RR and Microcystin-YR in Differentiated and Undifferentiated Caco-2 Cells. Toxicon 2009, 54, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Fawell, J.K.; James, C.P.; James, H.A. Toxins from Blue-Green Algae: Toxicological Assessment of micRocystin-LR and a Method for Its Determination in Water; Foundation for Water Research: Medmenham, Marlow, Bucks, UK, 1994; pp. 1–46. [Google Scholar]
- World Health Organization. Toxic Cyanobacteria in Water. A Guide to Public Health Consequence, Monitoring and Management; E and FN Spon: London, UK, 1999. [Google Scholar]
- Ballantyne, B. Toxicology and Hazard Eevaluation of Cyanide Fumigation Powders. Fund. Appl. Toxicol. 1988, 26, 325–335. [Google Scholar]
- Kotak, B.G.; Kenefick, S.L.; Fritz, D.L.; Rousseaux, C.G.; Prepas, E.E.; Hrudey, S.E. Occurrence and Toxicological Evaluation of Cyanobacterial Toxins in Alberta Lakes and Farm Dugouts. Water Res. 1993, 27, 495–506. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Namikoshi, M.; Choi, B.W. Structure and Biosynthesis of Toxins from Blue-green Algae (cyanobacteria). J. Appl. Phycol. 1994, 6, 159–176. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Cyanobacterial Toxins: Microcystin-LR in Drinking Water. In Guidelines for Drinking Water Quality; WHO: Geneva, Switzerland, 1998. [Google Scholar]
- Corbel, S.; Mougin, C.; Bouaïcha, N. Cyanobacterial toxins: Modes of Actions, Fate in Aquatic and Soil Ecosystems, Phytotoxicity and Bioaccumulation in Agricultural crops. Chemosphere 2014, 96, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Sano, T.; Kubota, R.; Kobayashi, N.; Tahara, M.; Obama, T.; Sugimoto, N.; Nishimura, T.; Ikarashi, Y. Effects of the Amino Acid Constituents of Microcystin Variants on Cytotoxicity to Primary Cultured Rat Hepatocytes. Toxins 2013, 6, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Bláha, L.; Babica, P.; Maršálek, B. Toxins Produced in Cyanobacterial Water Blooms-toxicity and Risks. Interdiscip. Toxicol. 2009, 2, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Pouria, S.; de Andrade, A.; Barbosa, J.; Cavalcanti, R.L.; Barreto, V.T.; Ward, C.J.; Preiser, W.; Poon, G.K.; Neild, G.H.; Codd, G.A. Fatal Microcystin Intoxication in Haemodialysis Unit in Caruaru, Brazil. Lancet 1998, 352, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Nishiwaki-Matsushima, R.; Ohta, T.; Nishiwaki, S.; Suganuma, M.; Kohyama, K.; Ishikawa, T.; Carmichael, W.W.; Fujiki, H. Liver Tumor Promotion by the Cyanobacterial Cyclic Peptide Toxin Microcystin-LR. J. Cancer Res. Clin. Oncol. 1992, 118, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.Z. Primary Prevention of Hepatocellular Carcinoma. J. Gastroen. Hepatol. 1995, 10, 674–682. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Chorus, I. Accumulation of Cyanobacterial Toxins in Freshwater “Seafood” and its Consequence for Public Health: A review. Environ. Pollut. 2007, 150, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Mulvenna, V.; Dale, K.; Priestly, B.; Mueller, U.; Humpage, A.; Shaw, G.; Allinson, G.; Falconer, I. Health Risk Assessment for Cyanobacterial Toxins in Seafood. Int. J. Environ. Res. Public Health 2012, 9, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Harada, K. Chemistry and detection of microcystins. In Toxic Microcystis, 1st ed.; Watanabe, M.F., Harada, K., Carmichael, W.W., Fujiki, H., Eds.; CRC Press: Boca Raton, FL, USA, 1995; pp. 110–114. [Google Scholar]
- Tsuji, K.; Masui, H.; Uemura, H.; Mori, Y.; Harada, K. Analysis of Microcystins in Sediments Using MMPB Method. Toxicon 2001, 39, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Carey, C.C.; Haney, J.F.; Cottingham, K.L. First Report of Microcystin-LR in the Cyanobacterium Gloeotrichia echinulata. Environ. Toxicol. 2007, 22, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.I.; Tsuji, K.; Watanabe, M.F.; Kondo, F. Stability of Microcystins from Cyanobacteria—III. Effect of pH and Temperature. Phycol. 1996, 35, 83–88. [Google Scholar] [CrossRef]
- Morris, R.J.; Williams, D.E.; Luu, H.A.; Holmes, C.F.B.; Andersen, R.J.; Calvert, S.E. The adsorption of microcystin-LR by natural clay particles. Toxicon 2000, 38, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Qian, Y.; Dai, S.; Feng, N. Adsorption of microcystin-LR and -LW on suspended particular matter (SPM) at different pH. Water Air Soil Pollut. 2008, 192, 67–76. [Google Scholar] [CrossRef]
- Hakanson, L. Suspended Particulate Matter in Lakes, Rivers, and Marine Systems; Blackburn Press: West Caldwell, NJ, USA, 2006. [Google Scholar]
- Munusamy, T.; Hu, Y.; Lee, J. Adsorption and Photodegradation of Microcystin-LR onto Sediments Collected from Reservoirs and Rivers in Taiwan: A Laboratory Study to Investigate the Fate, Transfer, and Degradation of Microcystin-LR. Environ. Sci. Pollut. Res. 2012, 19, 2390–2399. [Google Scholar] [CrossRef]
- Rudolph-BohÈhner, S.; Mierke, D.F.; Moroder, L. Molecular Structure of the Cyanobacterial Tumor-Producing Microcystins. FEBS Lett. 1994, 349, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Seitzinger, S.P. The Effect of pH on the Release of Phosphorus from Potomac Estuary Sediments: Implications for Blue-green Algal Blooms. Estuar. Shelf Coast. Sci. 1991, 33, 409–418. [Google Scholar] [CrossRef]
- Codd, G.A. Cyanobacterial Toxins: Occurrence, Properties, and Biological Significance. Water Sci. Technol. 1995, 32, 149–156. [Google Scholar] [CrossRef]
- Tsuji, K.; Naito, S.; Kondo, F.; Ishikawa, N.; Watanabe, M.F.; Suzuki, M.; Harada, K. Stability of Microcystins from Cyanobacteria—I. Effect of Light on Decomposition and Isomerization. Environ. Sci. Technol. 1994, 28, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; Steinberg, C. Rates of Humic Substance Photosensitized Degradation of Microcystin-LR in Natural Waters. Environ. Sci. Technol. 2000, 34, 3415–3419. [Google Scholar] [CrossRef]
- Tsuji, K.; Watanuki, T.; Kondo, F.; Watanabe, M.F.; Suzuki, S.; Nakazawa, H.; Suzuki, M.; Uchida, H.; Harada, K. Stability of Microcystins from Cyanobacteria—II. Effect of UV Light on Decomposition and Isomerization. Toxicon 1995, 33, 1619–1631. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, W.L.; Peng, P.G.; Sheng, G.Y.; Fu, J.M. Chemical and Molecular Heterogeneity of Humic Acids Repetitively Extracted from a Peat. Soil Sci. Soc. Am. J. 2003, 67, 740–746. [Google Scholar] [CrossRef]
- Song, W.; Bardowell, A.; O’Shea, K. Mechanistic Study of the Influence of Oxygen on the Photosensitized Transformations of Microcystins (Cyanotoxins). Environ. Sci. Technol. 2007, 41, 5336–5341. [Google Scholar] [CrossRef] [PubMed]
- Wörmer, L.; Huerta-Fontela, M.; Cirés, S.; Carrasco, D.; Quesada, A. Natural Photodegradation of the Cyanobacterial Toxins Microcystins and Cylindrospermopsin. Environ. Sci. Technol. 2010, 44, 3002–3007. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, M.G.; Shoemaker, J.A.; De La Cruz, A.A.; Dionysiou, D. Unveiling New Degradation Intermediates/Pathways from the Photocatalytic Degradation of Microcystin-LR. Environ. Sci. Technol. 2008, 42, 8877–8883. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Chen, D.; Huang, Y.; Yang, J.; Chen, G. Heterogeneous Fenton Photodegradation of Microcystin-LR with Visible Light Irradiation. Chinese J. Anal. Chem. 2011, 39, 540–543. [Google Scholar] [CrossRef]
- Antoniou, M.G.; Shoemaker, J.A.; De La Cruz, A.A.; Dionysiou, D.D. LC/MS/MS Structure Elucidation of Reaction Intermediates Formed during the TiO2 Photocatalysis of Microcystin-LR. Toxicon 2008, 51, 1103–1118. [Google Scholar] [CrossRef] [PubMed]
- Liu, I.; Lawton, L.A.; Cornish, B.; Robertson, P.K.J. Mechanistic and Toxicity Studies of the Photocatalytic Oxidation of Microcystin-LR. J. Photochem. Photobiol. A 2002, 148, 349–354. [Google Scholar] [CrossRef]
- Choi, H.; Antoniou, M.G.; Pelaez, M.; De La Cruz, A.A.; Shoemaker, J.A.; Dionysiou, D.D. Mesoporous Nitrogen-doped TiO2 for the Photocatalytic Destruction of the Cyanobacterial Toxin Microcystin-LR under Visible Light Irradiation. Environ. Sci. Technol. 2007, 41, 7530–7535. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Su, M.F.; Zhu, W.Q.; Li, X.N.; Jia, Y.; Guo, P.; Chen, Z.H.; Jiang, W.X. Growth Inhibition of Microcystis Aeruginosa by White-rot Fungus Lopharia Spadicea. Water Sci. Technol. 2010, 62, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Du, J.; Song, F.; Zhao, G.; Tian, X. A Fungus Capable of Degrading Microcystin-LR in the Algal Culture of Microcystis Aeruginosa PCC7806. Appl. Biochem. Biotechnol. 2012, 166, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Bourne, D.G.; Jones, G.J.; Blakeley, R.L.; Jones, A.; Negri, A.P.; Riddles, P. Enzymatic Pathway for the Bacterial Degradation of the Cyanobacterial Cyclic Peptide Toxin Microcystin-LR. Appl. Environ. Microbiol. 1996, 62, 4086–4094. [Google Scholar] [PubMed]
- Bourne, D.G.; Riddles, P.; Jones, G.J.; Smith, W.; Blakeley, R.L. Characterisation of a Gene Cluster Involved in Bacterial Degradation of the Cyanobacterial Degradation of the Cyanobacterial Toxin Microcystin-LR. Environ. Toxicol. 2001, 16, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Imanishi, S.; Kato, H.; Mizuno, M.; Tsuji, K.; Harada, K. Bacterial Degradation of Microcystins and Nodularin. Chem. Res. Toxicol. 2005, 18, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Imanishi, S.; Kato, H.; Mizuno, M.; Ito, E.; Tsuji, K. Isolation of ADDA from Microcystin-LR by Microbial Degradation. Toxicon. 2004, 44, 107–109. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.; Gaudieux, A.L.; Fanok, S.; Newcombe, G.; Humpage, A.R. Bacterial Degradation of Microcystin Toxins in Drinking Water Eliminates their Toxicity. Toxicon 2007, 50, 438–441. [Google Scholar] [CrossRef] [PubMed]
- Okano, K.; Shimizu, K.; Kawauchi, Y.; Maseda, H.; Utsumi, M.; Zhang, Z.; Neilan, B.A.; Sugiura, N. Characteristics of a Microcystin-degrading Bacterium under Alkaline Environmental Conditions. J. Toxicol. 2009. [Google Scholar] [CrossRef]
- Christoffersen, K.; Lyck, S.; Winding, A. Microbial Activity and Bacterial Community Structure during Degradation of Microcystins. Aquat. Microb. Ecol. 2002, 27, 125–136. [Google Scholar] [CrossRef]
- Steffen, M.M.; Li, Z.; Effler, C.; Hauser, L.J.; Boyer, G.L.; Wilhelm, S.W. Comparative Metagenomics of Toxic Freshwater Cyanobacteria Bloom Communities on Two Continents. PLoS One 2012, 7, e44002. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.W.; Farnsley, S.E.; LeCleir, G.R.; Layton, A.C.; Satchwell, M.F.; DeBruyn, J.M.; Boyer, G.L.; Zhu, G.; Paerl, H.W. The Relationships between Nutrients, Cyanobacterial Toxins and the Microbial Community in Taihu (Lake Tai), China. Harmful Algae 2011, 10, 207–215. [Google Scholar] [CrossRef]
- Jones, G.J.; Orr, P.T. Release and Degradation of Microcystin Following Algicide Treatment of a Microcystis Aeruginosa Bloom in a Recreational Lake, as 248 Determined by HPLC and Protein Phosphatase Inhibition assay. Water Res. 1994, 28, 871–876. [Google Scholar] [CrossRef]
- Park, H.; Sasaki, Y.; Maruyami, T.; Yanagisawa, E.; Hiraishi, A.; Kato, K. Degradation of the Cyanobacterial Heptatotoxin Microcystin by a New Bacterium Isolated from a Hypertrophic Lake. Environ. Toxicol. 2001, 6, 337–343. [Google Scholar] [CrossRef]
- Saito, T.; Sugiura, N.; Itayama, T.; Inamori, Y.; Matsumura, M.J. Degradation Characteristics of Microcystins by Isolated Bacteria from Lake Kasumigaura. J. Water Supply Res. Technol. 2003, 52, 13–18. [Google Scholar]
- Rapala, J.; Berg, K.A.; Lyra, C.; Niemi, R.M.; Manz, W.; Suomalainen, S.; Paulin, L.; Lahti, K. Paucibacter toxinivorans gen. nov., sp. nov., a bacterium that degrades cyclic cyanobacterial hepatotoxins microcystins and nodularins. Int. J. Syst. Evol. Micr. 2005, 55, 1563–1568. [Google Scholar] [CrossRef]
- Maruyama, T.; Park, H.; Ozawa, K.; Tanaka, Y.; Sumino, T.; Hamana, K.; Hirashi, A.; Kato, K. Sphingosinicella microcystinivorans gen. nov., sp. nov., a microcystin degrading bacterium. Int. J. Syst. Evol. Micr. 2006, 56, 85–89. [Google Scholar] [CrossRef]
- Lahti, K.; Niemi, M.R.; Rapala, J.; Sivonen, K. Biodegradation of Cyanobacterial Hepatotoxins-Characterisation of Toxin Degrading Bacteria. In Harmful Algae; Reguera, B., Blanco, J., Fernández, M.L., Wyatt, T., Eds.; United Nations Educational, Scientific, and Cultural Organization: Vigo, Spain, 1988; pp. 363–365. [Google Scholar]
- Berg, K.A.; Lyra, C.; Sivonen, K.; Paulin, L.; Suomalainen, S.; Tuomi, P.; Rapala, J. High Diversity of Cultivable Heterotrophic Bacteria in Association with Cyanobacterial Water Blooms. ISME J. 2009, 3, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.; Huang, H.; Kwon, Y.; Greengard, P.; Nairn, A.C.; Kuriyan, J. Three Dimensional Structure of the Catalytic Subunit of Protein Serine/threonine Phosphatase-1. Nature 1995, 376, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, N.P.E. Role of Metabolism in Chemical Toxicity. In Cytochromes P450: Metabolic and Toxicological Aspects; Ioannides, C., Ed.; CRC Press: Boca Raton, FL, USA, 1996; pp. 29–49. [Google Scholar]
- Manahan, S.E. Toxicological Chemistry and Biochemistry, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2003; pp. 142–153. [Google Scholar]
- Townsend, D.M.; Tew, K.D. The role of glutathione-S-transferase in anti-cancer drug resistance. Oncogene 2003, 22, 7369–7375. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Wahlländer, A.; Waydhas, C.; Soboll, S.; Häberle, D. Functions of Intracellular Glutathione in Hepatic Hydroperoxide and Drug Metabolism and the Role of Extracellular Glutathione. Adv. Enzyme Regul. 1980, 18, 303–320. [Google Scholar] [CrossRef] [PubMed]
- Lindwall, G.; Boyer, T.D. Excretion of Glutathione Conjugates by Primary Cultured Rat Hepatocytes. J. Biol. Chem. 1987, 262, 5151–5158. [Google Scholar] [PubMed]
- Lilja, H.; Jeppsson, J.; Kristensson, H. Evaluation of Serum γ-Glutamyltransferase by Electrofocusing, and Variations in Isoform Patterns. Clin. Chem. 1983, 29, 1034–1037. [Google Scholar] [PubMed]
- Ishikawa, T. The ATP-Dependent Glutathione S-Conjugate Export Pump. Trends Biochem. Sci. 1992, 17, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Müeller, M.; Meijer, C.; Zaman, G.R.; Borst, P.; Scheper, R.J.; Mulder, N.H.; DeVries, E.E.; Jansen, P.L. Overexpression of the Gene Encoding the Multidrug Resistance-associated Protein Results in Increased ATP-dependent Glutathione S-conjugate Transport. J. Med. Sci. 1994, 91, 13033–13037. [Google Scholar]
- Ito, E.; Satake, M.; Yasumoto, T. Pathological Effects of Lyngbyatoxin A upon Mice. Toxicon 2002, 40, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.E.; Dawe, S.C.; Kent, M.; Andersen, R.J.; Craig, M.; Holmes, C.F.B. Bioaccumulation and Clearance of Microcystins from Salt Water Mussels, Mytilus edulis, and in vivo Evidence for Covalently Bound Microcystins in Mussel Tissues. Toxicon 1997, 35, 1617–1625. [Google Scholar] [CrossRef] [PubMed]
- Pflugmacher, S.; Wiegand, C.; Oberemm, A.; Beattie, K.A.; Krause, E.; Codd, G.A.; Steinberg, C.E.W. Identification of an Enzymatically Formed Glutathione Conjugate of the Cyanobacterial Hepatotoxin Microcystin-LR: The First Step of Detoxication. Biochim. Biophys. Acta Gen. Subj. 1998, 1425, 527–533. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Codd, G.A.; Steinberg, C.E.W. Effects of the Cyanobacterial Toxin Microcystin-LR on Detoxication Enzymes in Aquatic Plants. Environ. Toxicol. 1999, 14, 111–115. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Weigand, C.; Beattie, K.A.; Krause, E.; Steinberg, C.E.W.; Codd, G.A. Uptake, Effects, and Metabolism of Cyanobacterial Toxins in the Emergent Reed Plant Phragmites Australis (Cav.) Trin. Ex Steud. Environ. Toxicol. Chem. 2001, 20, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Pflugmacher, S. Promotion of Oxidative Stress in the Aquatic Macrophyte Ceratophyllum Demersum during Biotransformation of the Cyanobacterial Toxin Microcystin-LR. Aquat. Toxicol. 2004, 70, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Meissner, S.; Fastner, J.; Dittmann, E. Microcystin Production Revisited: Conjugate Formation Makes a Major Contribution. Environ. Microbiol. 2013, 15, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Schulz, K.L.; Zimba, P.V.; Boyer, G.L. Possible Mechanism for the Foodweb Transfer of Covalently Bound Microcystins. Ecotox. Environ. Saf. 2010, 73, 757–761. [Google Scholar] [CrossRef]
- Lance, E.; Petit, A.; Sanchez, W.; Paty, C.; Ge´rard, C.; Bormans, M. Evidence of Trophic Transfer of Microcystins from the Gastropod Lymnaea stagnalis to the Fish Gasterosteus Aculeatus. Harmful Algae 2014, 31, 9–17. [Google Scholar] [CrossRef]
- Zilliges, Y.; Kehr, J.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Borner, T.; Dittmann, E. The Cyanobacterial Hepatotoxin Microcystin Binds to Proteins and Increases the Fitness of Microcystis under Oxidative Stress Conditions. PLoS One 2013, 6, e17615. [Google Scholar] [CrossRef]
- Schmidt, J.R.; Shaskus, M.; Estenik, J.F.; Oesch, C.; Khidekel, R.; Boyer, G.L. Variations in the Microcystin Content of Different Fish Species Collected from a Eutrophic Lake. Toxins 2013, 5, 992–1009. [Google Scholar] [CrossRef] [PubMed]
- Robinson, N.A.; Pace, J.G.; Matson, C.F.; Miura, G.A.; Lawrence, W.B. Tissue Distribution, Excretion and Hepatic Biotransformation of Microcystin-LR in Mice. J. Pharmacol. Exp. Ther. 1991, 256, 176–182. [Google Scholar] [PubMed]
- Amorim, A.; Vasconcelos, V. Dynamics of Microcystins in the Mussel. Mytilus galloprovincialis. Toxicon 1999, 37, 1041–1052. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Liebig, J.R.; Carmichael, W.W.; Agy, M.A.; Johengen, T.H.; Fahnenstiel, G.L.; Nalepa, T.F. Zebra Mussel (Dreissena polymorpha) Selective Filtration Promoted Toxic Microcystis blooms in Saginaw Bay (Lake Huron) and Lake Erie. Can. J. Fish. Aquat. Sci. 2001, 58, 1208–1221. [Google Scholar] [CrossRef]
- Hooser, S.B.; Kuhlenschmidt, M.S.; Dahlem, A.M.; Beasley, V.R.; Carmichael, W.W.; Haschek, W.M. Uptake and Subcellular Localization of Tritiated Dihydro-microcystin-LR in Rat Liver. Toxicon 1991, 29, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Runnegar, M.T.C.; Gerdes, R.G.; Falconer, I.R. The Uptake of the Cyanobacterial Hepatotoxin Microcystin by Isolated Rat Hepatocytes. Toxicon 1991, 29, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Sahin, A.; Tencalla, F.G.; Dietrich, D.R.; Naegeli, H. Biliary Excretion of Biochemically Active Cyanobacteria (blue-green algae) Hepatotoxins in Fish. Toxicology 1996, 106, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Kondo, F.; Matsumoto, H.; Yamada, S.; Ishikawa, N.; Ito, E.; Nagata, S.; Ueno, Y.; Suzuki, M.; Harada, K. Detection and Identification of Metabolites of Microcystins formed in Vivo in Mouse and Rat Livers. Chem. Res. Toxicol. 1996, 9, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, S. Covalent Glutathione Conjugation to Cyanobacterial Hepatotoxin Microcystin-LR by F344 Rat Cytosolic and Microsomal Glutathione S-transferases. Environ. Toxicol. Phar. 2001, 9, 135–139. [Google Scholar] [CrossRef]
- Xie, L.; Xie, P.; Guo, L.; Li, L.; Miyabara, Y.; Park, H. Organ Distribution and Bioaccumulation of Microcystins in Freshwater Fish at Different Trophic Levels from the Eutrophic Lake Chaohu, China. Environ. Toxicol. 2005, 20, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Yokoyama, A.; Nakamura, K.; Park, H. Accumulation of Microcystins in Various Organs of the Freshwater Snail Sinotaia histrica and Three Fishes in a Temperate Lake, the Eutrophic Lake Suwa, Japan. Toxicon 2007, 49, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Deng, X.; Xie, P.; Yang, Q.; Chen, J.; Dai, M. Determination of Microcystin-LR and its Metabolites in Snail (Bellamya aeruginosa), Shrimp (Macrobrachium nipponensis) and Silver Carp (Hypophthalmichthys molitrix) from Lake Taihu, China. Chemosphere. 2009, 76, 974–981. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Chen, J.; Xie, P.; Zhang, D.; Li, G.; Wu, L.; Zhang, W.; Guo, X.; Li, S. Quantitatively Evaluating Detoxification of the Hepatotoxic Microcystins through the Glutathione and Cysteine Pathway in the Cyanobacterial-eating Bighead Carp. Aquat. Toxicol. 2012, 116–117, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Yang, Q.; Xie, P.; Deng, X.; Chen, J.; Dai, M. The Role of Cysteine Conjugation in the Detoxification of Microcystin-LR in Liver of Bighead Carp (Aristichthys nobilis): A Field and Laboratory Study. Ecotoxicology 2012, 21, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Buratti, F.M.; Scardala, S.; Funari, E.; Testai, E. Human Glutathione Transferases Catalyzing the Conjugation of the Hepatatoxin Microcystin-LR. Chem. Res. Toxicol. 2011, 24, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Beattie, K.A.; Pflugmacher, S.; Codd, G.A. Immuno-crossreactivity and toxicity assessment of conjugation products of the cyanobacterial toxin, microcystin-LR. FEMS Microbiol. Lett. 2000, 189, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Vasconcelos, V. Molecular Mechanisms of Microcystin Toxicity in Animal Cells. Int. J. Mol. Sci. 2010, 11, 268–287. [Google Scholar] [CrossRef] [PubMed]
- Ito, E.; Takai, A.; Kondo, F.; Masui, H.; Imanishi, S.; Harada, K. Comparison of protein phosphatase inhibitory activity and apparent toxicity of microcystins and related compounds. Toxicon 2001, 40, 1017–1025. [Google Scholar] [CrossRef]
- Smith, J.L.; Haney, J.F. Foodweb Transfer, Accumulation, and Depuration of Microcystins, a Cyanobacterial Toxin, in Pumpkinseed Sunfish (Lepomis gibbosus). Toxicon 2006, 48, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Moreno, I.M.; Molina, R.; Jos, A.; Picó, Y.; Cameán, A.M. Determination of Microcystins in Fish by Solvent Extraction and Liquid Chromatography. J. Chromatagr. A. 2005, 1080, 199–203. [Google Scholar] [CrossRef]
- Oberhaus, L.; Gelinas, M.; Pinel-Alloul, B.; Ghadouani, A.; Humbert, J. Grazing of Two Toxic Planktothrix Species by Daphnia pulicaria: Potential for Bloom Control and Transfer of Microcystins. J. Plankton Res. 2007, 29, 827–838. [Google Scholar] [CrossRef]
- Smith, J.A.; Witkowski, P.J.; Fusillo, T.V. Manmade Organic Compounds in the Surface Waters of the United States—A review of Current Understanding. U.S. Geolo. Surv. Circ. 1988, 1007, 92. [Google Scholar]
- Nowell, L.H.; Capel, P.D.; Dileanis, P.D. Pesticides in Stream Sediment and Aquatic Biota—Distribution, Trends, and Governing Factors; CRC Press: Boca Raton, FL, USA, 1999. [Google Scholar]
- Freitas de Magalhães, V.F.; Soares, R.M.; Azevedo, S.M.F.O. Microcystin Contamination in Fish from the Jacarepagua Lagoon (Rio de Janeiro, Brazil): Ecological Implication and Human Health Risk. Toxicon 2001, 39, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Prepas, E.E.; Kotak, B.G.; Campbell, L.M.; Evans, J.C.; Hrudey, S.E.; Holmes, C.F.B. Accumulation and Elimination of Cyanobacterial Hepatotoxins by the Freshwater Clam Anodonta grandis simpsoniana. Can. J. Fish. Aquat. Sci. 1997, 54, 41–46. [Google Scholar] [CrossRef]
- Zhang, D.; Xie, P.; Liu, Y.; Qiu, T. Transfer, Ddistribution and Bioaccumulation of Microcystins in the Aquatic Food Web in Lake Taihu, China, with Potential Risks to Human Health. Sci. Total Environ. 2013, 7, 2191–2199. [Google Scholar]
- Poste, A.; Hecky, R.E.; Guildford, S.J. Evaluating Microcystin Exposure Risk through Fish Consumption. Environ. Sci. Technol. 2011, 45, 5806–5811. [Google Scholar] [CrossRef] [PubMed]
- Amrani, A.; Nasri, H.; Azzouz, A.; Kadi, Y.; Bouaïcha, N. Variation in Cyanobacterial Hepatotoxin (microcystin) Content of Water Samples and two Species of Fishes Collected from a Shallow Lake in Algeria. Arch. Environ. Contam. Toxicol. 2014, 66, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Liu, Y.; Chen, W.; Liu, L.; Kent, M.; Song, L. Health Risks Associated with Consumption of Microcystin-contaminated Fish and Shellfish in three Chinese Lakes: Significance for Freshwater Aquacultures. Ecotox. Environ. Saf. 2010, 73, 1804–1811. [Google Scholar] [CrossRef]
- Jia, J.; Luo, W.; Lu, Y.; Giesy, J.P. Bioaccumulation of Microcystins (MCs) in Four Fish Species from Lake Taihu, China: Assessment of Risks to Humans. Sci. Total Environ. 2014, 487, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Leland, H.V.; Kuwabara, J.S. Trace metals. In Fundamentals of Aquatic Toxicology; Rand, G.M., Petrocelli, S.R., Eds.; CRC Press: New York, NY, USA, 1985; pp. 374–415. [Google Scholar]
- Suedel, B.C.; Boraczek, J.A.; Peddicord, R.K.; Clifford, P.A.; Dillon, T.M. Trophic Transfer and Biomagnification Potential of Contaminants in Aquatic Ecosystems. Rev. Environ. Contam. Toxicol. 1994, 136, 21–89. [Google Scholar] [PubMed]
- Kozlowsky-Suzuki, B.; Wilson, A.E.; Ferrão-Filho, A. Biomagnification or Biodilution of Microcystins in Aquatic Foodwebs? Meta-analyses of Laboratory and Field Studies. Harmful Algae 2012, 18, 47–55. [Google Scholar] [CrossRef]
- Papadimitriou, T.; Kagalou, I.; Stalikas, C.; Pilidis, G.; Leonardos, I.D. Assessment of Microcystin Distribution and Biomagnification in Tissues of Aquatic Food Web Compartments from a Shallow Lake and Evaluation of Potential Risks to Public Health. Ecotoxicology 2012, 21, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Ibelings, B.W.; Bruning, K.; de Jonge, J.; Wolfstein, K.; Dionisio Pires, L.M.; Postma, J.; Burger, T. Distribution of Microcystins in a Lake Foodweb: No Evidence for Biomagnification. Microbial Ecol. 2005, 49, 487–500. [Google Scholar] [CrossRef]
- Gustafsson, S.; Hanson, L. Development of Tolerance against Toxic Cyanobacteria in Daphnia. Aquat. Ecol. 2004, 38, 37–44. [Google Scholar] [CrossRef]
- Guo, N.; Xie, P. Development of Tolerance against Toxic Microcystis aeruginosa in Three Cladocerans and the Ecological Implications. Environ. Pollut. 2006, 143, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, C.; Wiegand, C.; Ame, M.V.; Nicklisch, A.; Wunderlin, D.; Pflugmacher, S. The Effects of a Cyanobacterial Crude Extract on Different Aquatic Organisms: Evidence for Cyanobacterial Toxin Modulating Factors. Environ. Toxicol. 2001, 16, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, A.; Park, H. Depuration Kinetics and Persistence of the Cyanobacterial Toxin Microcystin-LR in the Freshwater Bivalve Unio douglasiae. Environ. Toxicol. 2003, 18, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Pires, L.M.; Bones, B.M.; Van Donk, E.; Ibelings, B.W. Grazing on Colonial and Filamentous, Toxic and Non-toxic Cyanobacteria by the Zebra Mussel Dreissena polymorpha. J. Plankton Res. 2004, 27, 331–339. [Google Scholar] [CrossRef]
- Juhel, G.; Davenport, J.; O’Halloran, J.; Culloty, S.; Ramsay, R.; James, K.; Furey, A.; Allis, O. Pseudodiarrhoea in Zebra Mussels Dreissena polymorpha (Pallas) Exposed to Microcystins. J. Exp. Biol. 2006, 209, 810–816. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, V.M.; Wiegand, C.; Pflugmacher, S. Dynamics of Glutathione-S-transferases in Mytilus galloprovincialis Exposed to Toxic Microcystis Aeruginosa Cells, Extracts and Pure Toxins. Toxicon 2007, 50, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Contardo-Jara, V.; Pflugmacher, S.; Wiegand, C. Multi-xenobiotic Resistance a Possible Explanation for the Insensitivity of Bivalves towards Cyanobacterial Toxins. Toxicon 2008, 52, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Blanchette, M.; Haney, J.F. The Effect of Toxic Microcystis Aeruginosa on Four Different Populations of Daphnia. UNH Cent. Freshw. Biol. Res. 2002, 4, 1–10. [Google Scholar]
- Råbergh, C.M.I.; Bylund, G.; Eriksson, J.E. Histopathological Effects of Microcystin-LR, a Cyclic Peptide Toxin from the Cyanobacterium (blue-green alga) Microcystis aeruginosa, on Common Carp (Cyprinus carpio L.). Aquat. Toxicol. 1991, 20, 131–146. [Google Scholar] [CrossRef]
- Malbrouck, C.; Trausch, G.; Devos, P.; Kestemont, P. Hepatic Accumulation and Effects of Microcystin-LR on Juvenile Goldfish Carassius auratus L. Comp. Biochem. Phys. 2003, 135, 39–48. [Google Scholar] [CrossRef]
- Kotak, B.G.; Semalulu, S.; Fritz, D.L.; Prepas, E.E.; Hrudey, S.E.E.; Coppock, R.W. Hepatic and Renal Pathology of Intraperitoneally Administered Microcystin-LR in Rainbow Trout (Oncorhynchus mykiss). Toxicon 1996, 34, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liu, J.; Wu, K.; Qu, Q.; Fan, F.; Klaassen, C.D. Overexpression of Nrf2 Protects Against Microcystin-induced Hepatotoxicity in Mice. PLoS One 2014, 9, e93013. [Google Scholar] [CrossRef] [PubMed]
- Falconer, I.R.; Jackson, A.R.B.; Langley, J.; Runnegar, M.T. Liver Pathology in Mice in Poisoning by the Blue-green Alga Microcystis aeruginosa. Aust. J. Biol. Sci. 1981, 34, 179–187. [Google Scholar]
- Sivonen, K.; Jones, G.J. Cyanobacterial toxins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; E and FN Spon: London, UK, 1999; pp. 41–111. [Google Scholar]
- Smith, J.L.; Boyer, G.L.; Mills, E.; Schulz, K.L. Toxicity of Microcystin-LR, a Cyanobacterial Toxin, to Multiple Life Stages of the Burrowing Mayfly, Hexagenia, and Possible Implications for Recruitment. Environ. Toxicol. 2007, 23, 499–506. [Google Scholar] [CrossRef]
- Zhang, D.; Deng, X.; Xie, P.; Chen, J.; Guo, L. Risk Assessment of Microcystins in Silver Carp (Hypophthalmichthys molitrix) from Eight Eutrophic Lakes in China. Food Chem. 2013, 140, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xie, P.; Zhang, D.; Lei, H. In Situ Studies on the Distribution Parameters and Dynamics of Microcystins in Biomanipulation Fish-bighead Carp (Aristichthys nobilis). Environ. Pollut. 2007, 147, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Namikoshi, M.; Rinehart, K.I. Bioactive Compounds Produced by Cyanobacteria. J. Ind. Microbiol. 1996, 17, 373–384. [Google Scholar] [CrossRef]
- Lampert, W. Inhibitory and Toxic Effects on Blue-green Algae on Daphnia. Int. Rev. Gestamen Hydrobiol. 1981, 66, 285–298. [Google Scholar] [CrossRef]
- Nizan, S.; Dimentman, C.; Shilo, M. Acute Toxic Effects of Cyanobacterium Microcystis aeruginosa on Daphnia magna. Limnol. Oceanogr. 1986, 31, 497–502. [Google Scholar] [CrossRef]
- Fulton, R.S., III; Paerl, H.W. Effects of Colonial Morphology on Zooplankton Utilization of Algal Sources during Blue-green Algal (Microcystis aeruginosa) Blooms. Limnol. Oceanogr. 1987, 32, 634–644. [Google Scholar] [CrossRef]
- Fulton, R.S., III; Paerl, H.W. Toxic and Inhibitory Effects of the Blue-green Alga Microcystis aeruginosa on Herbivorous Zooplankton. J. Plankton Res. 1987, 9, 837–855. [Google Scholar] [CrossRef]
- Benndorf, J.; Henning, M. Daphnia and Toxic Blooms of Microcystis aeruginosa in Bautzen Reservoir (GDR). Int. Rev. Gestamen Hydrobiol. 1989, 74, 233–248. [Google Scholar] [CrossRef]
- Rohrlack, T.; Dittmann, E.; Henning, M.; Borner, T.; Kohl, J. Role of Microcystins in Poisoning and Food Ingestion Inhibition by Daphnia Galeata Caused by the Cyanobacterium Microcystis aeruginosa. Appl. Environ. Microbiol. 1999, 65, 737. [Google Scholar] [PubMed]
- Chislock, M.F.; Sarnelle, O.; Jernigan, L.M.; Wilson, A.E. Do High Concentrations of Microcystin Prevent Daphnia Control of Phytoplankton? Water Res. 2013, 47, 1961–1970. [Google Scholar] [CrossRef] [PubMed]
- Porter, K.G.; Orcutt, J.D. Nutritional Adequacy, Manageability, and Toxicity as Factors that Determine Food Quality of Green and Blue-green Algae for Daphnia. In Evolution and Ecology of Zooplankton Communities; Kerfoot, W.C., Ed.; University Press of New England: Hanover, New Hampshire, UK, 1980; pp. 268–281. [Google Scholar]
- Holm, N.P.; Shapiro, J.S. An Examination of Lipid Reserves and the Nutritional Status of Daphnia pulex Fed Aphanizomenon flos-aquae. Limnol. Oceanogr. 1984, 28, 677–687. [Google Scholar] [CrossRef]
- Ahlgren, G.; Lundstedt, L.; Brett, M.; Forsberg, C. Lipid Composition and Food Quality of Some Freshwater Phytoplankton for Cladoceran Zooplankters. J. Plankton Res. 1990, 12, 809–818. [Google Scholar] [CrossRef]
- Van Gremberghe, I.V.; Vanormelingren, P.; Vanelslander, B.; van der Gucht, K.; D’hondt, S.; de Meester, L.; Vyverman, W. Genotype-dependent Interactions among Sympatric Microcystis Strains Mediated by Daphnia Grazing. Oikos 2009, 118, 1647–1658. [Google Scholar] [CrossRef]
- DeMott, W.R.; Dhawale, S. Inhibition of in-vitro Protein Phosphatase Activity in Three Zooplankton Species by Microcystin-LR, a Toxin from Cyanobacteria. Arch. Hydrobiol. 1995, 134, 417–424. [Google Scholar]
- DeMott, W.R.; Zhang, Q.; Carmichael, W.W. Effects of Toxic Cyanobacteria and Purified Toxins on the Survival and Feeding of a Copepod and Three Species of Daphnia. Limnol. Oceanogr. 1991, 36, 1346–1357. [Google Scholar] [CrossRef]
- Reinikainen, M.; Lindvall, F.; Meriluoto, J.A.O.; Repka, S.; Sivonen, K.; Spoof, L.; Wahlsten, M. Effects of Dissolved Cyanobacterial Toxins on the Survival and Egg Hatching of Estuarine Calanoid Copepods. Mar. Biol. 2002, 140, 577–583. [Google Scholar] [CrossRef]
- Ger, K.A.; Teh, S.J.; Goldman, C.R. Microcystin-LR Toxicity on Dominant Copepods Eurytemora affinis and Pseudodiaptomus forbesi of the upper San Francisco Estuary. Sci. Total Environ. 2009, 407, 4852–4857. [Google Scholar] [CrossRef] [PubMed]
- DeMott, W.R.; Moxter, F. Foraging on Cyanobacteria by Copepods: Responses to Chemical Defenses and Resource Abundance. Ecology 1991, 72, 1820–1834. [Google Scholar] [CrossRef]
- DeMott, W.R. Foraging Strategies and Growth Inhibition in five Daphnids Feeding on Mixtures of a Toxic Cyanobacterium and a Green Alga. Freshwater Biol. 1999, 42, 263–274. [Google Scholar] [CrossRef]
- Ortiz-Rodríguez, R.; Wiegand, C. Age Related Acute Effects of Microcystin-LR on Daphnia Magna Biotransformation and Oxidative Stress. Toxicon 2010, 56, 1342–4349. [Google Scholar] [CrossRef] [PubMed]
- Hairston, N.G.; Lampert, W.; Cáceres, C.E.; Holtmeier, C.L.; Weider, L.J.; Gaedke, U.; Fischer, J.M.; Fox, J.A.; Post, D.M. Rapid Evolution Revealed by Dormant Eggs. Nature 1999, 401, 44. [Google Scholar] [CrossRef]
- Geis-Asteggiante, L.; Lehotay, S.J.; Fortis, L.L.; Paoli, G.; Wijey, C.; Heinzen, H. Development and Validation of a Rapid Method for Microcystins in Fish and Comparing LC-MS/MS Results with ELISA. Anal. Bioanal. Chem. 2011, 401, 2617–2630. [Google Scholar] [CrossRef] [PubMed]
- Kohoutek, J.; Adamovský, O.; Oravec, M.; Simek, Z.; Palíková, M.; Kopp, R.; Bláha, L. LC-MS Analyses of Microcystins in Fish Tissues Overestimate Toxin Levels-critical Comparison with LC-MS/MS. Anal. Bioanal. Chem. 2010, 398, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Boyer, G.L. Standardization of Microcystin Extraction from Fish Tissues: A Novel Internal Standard as a Surrogate for Polar and Non-polar Variants. Toxicon 2009, 53, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Mekebri, A.; Blondina, G.J.; Crane, D.B. Method Validation of Microcystins in Water and Tissue by Enhanced Liquid Chromatography Tandem Mass Spectrometry. J. Chromatogr. A 2009, 1216, 3147–3155. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, K.M.; Spoof, L.E.M.; Meriluoto, J.A.O. Quantitative LC-ESF-MS Analyses of Microcystins and Nodularin-R in Animal Tissue-matrix Effects and Method Validation. Environ. Toxicol. 2005, 20, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Ernst, B.; Bietz, L.; Hoeger, S.J.; Dietrich, D.R. Recovery of MC-LR in Fish Liver Tissue. Environ. Toxicol. 2005, 20, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Adamovský, O.; Kopp, R.; Hilscherová, K.; Babica, P.; Palíková, M.; Pašková, V.; Navrátil, S.; Maršálek, B.; Bláha, L. Microcystin Kinetics (bioaccumulation and elimination) and Biochemical Responses in Common Carp (Cyprinus carpio) and Silver Carp (Hypophthalmichthys molitrix) Exposed to Toxic Cyanobacterial Blooms. Environ. Toxicol. Chem. 2007, 26, 2687–2693. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Xie, P.; Liang, G.; Chen, J.; Lei, H. Simultaneous Determination of Microcystin-LR and its Glutathione Conjugate in Fish Tissues by Liquid Chromatography-tandem Mass Spectrometry. J. Chromatogr. B 2008, 862, 43–50. [Google Scholar] [CrossRef]
- Barron, M.G.; Stehly, G.R.; Hayton, W.L. Pharmacokinetic Modeling in Aquatic Animals I. Models and concepts. Aquatic Toxicol. 1990, 18, 61–86. [Google Scholar] [CrossRef]
- Dyble, J.; Gossiaux, D.; Landrum, P.; Kashian, D.R.; Pothoven, S. A Kinetic Study of Accumulation and Elimination of Microcystin-LR in Yellow Perch (Perca flavescens) Tissue and Implications for Human Fish Consumption. Mar. Drugs 2011, 9, 2553–2571. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.S.; Magnusson, B.M.; Burczynski, F.J.; Weiss, M. Enterohepatic Circulation: Physiological, Pharmacokinetic and Clinical Implications. Clin. Pharmacokinet. 2002, 41, 751–790. [Google Scholar] [CrossRef] [PubMed]
- Beasley, V.R.; Stotts, R.R. Toxicokinetics of Microcystin and Dihydro-microcystin in Swine. US Army report. 1994. Available online: http://www.dtic.mil/cgibin/GetTRDoc?AD=ADA280697&Location=U2&doc=GetTRDoc.pdf (accessed on 25 June 2014).
- Li, T.; Huang, P.; Liang, J.; Fu, W.; Guo, Z.; Xu, L. Microcystin-LR (MCLR) Induces a Compensation of PP2A Activity Mediated by α4 Protein in HEK293 Cells. Int. J. Biol. Sci. 2011, 7, 740–752. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Chu, F.S. Kinetics of Distribution of Microcystin-LR in Serum and Liver Cytosol of Mice: An Immunochemical Analysis. J. Agric. Food Chem. 1994, 42, 1035–1040. [Google Scholar] [CrossRef]
- Robinson, N.A.; Miura, G.A.; Matson, C.F.; Dinterman, R.E.; Pace, J.G. Characterization of Chemically Tritiated Microcystin-LR and its Distribution in Mouse. Toxicon 1990, 27, 1035–1042. [Google Scholar] [CrossRef]
- Brooks, W.P.; Codd, G.A. Distribution of Microcystis Aeruginosa Peptide Toxin and Interactions with Hepatic Microsomes in Mice. Pharmacol. Toxicol. 1987, 60, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Xie, P.; Chen, J.; Liang, G.; Yu, T.; Jiang, Y. Tissue Distribution and Depuration of the Extracted Hepatotoxic Cyanotoxin Microcystins in Crucian Carp (Carassius carassius) Intraperitoneally Injected at a Sublethal Dose. ScientificWorldJournal 2008, 8, 713–719. [Google Scholar] [CrossRef]
- Rohrlack, T.; Christofferson, K.; Dittmann, E.; Nogueira, I.; Vasconcelos, V.; Borner, T. Ingestion of Microcystins by Daphnia: Intestinal Uptake and Toxic Effects. Limnol. Oceanogr. 2005, 50, 440–448. [Google Scholar] [CrossRef]
- Sadler, T.; von Elert, E. Dietary Exposure of Daphnia to Microcystins: No in vivo Relevance of Biotransformation. Aquat. Toxicol. 2014, 150, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, J.; Xie, P.; He, J.; Guo, X.; Tuo, X.; Zhang, W.; Wu, L. Rapid Conversion and Reversible Conjugation of Glutathione Detoxification of Microcystins in Bighead Carp (Aristichthys nobilis). Aquat. Toxicol. 2014, 147, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Agha, R.; Quesada, A. Oligopeptides as Biomarkers of Cyanobacterial Subpopulations. Toward an Understanding of Their Biological Role. Toxins 2014, 6, 1929–1950. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidt, J.R.; Wilhelm, S.W.; Boyer, G.L. The Fate of Microcystins in the Environment and Challenges for Monitoring. Toxins 2014, 6, 3354-3387. https://doi.org/10.3390/toxins6123354
Schmidt JR, Wilhelm SW, Boyer GL. The Fate of Microcystins in the Environment and Challenges for Monitoring. Toxins. 2014; 6(12):3354-3387. https://doi.org/10.3390/toxins6123354
Chicago/Turabian StyleSchmidt, Justine R., Steven W. Wilhelm, and Gregory L. Boyer. 2014. "The Fate of Microcystins in the Environment and Challenges for Monitoring" Toxins 6, no. 12: 3354-3387. https://doi.org/10.3390/toxins6123354
APA StyleSchmidt, J. R., Wilhelm, S. W., & Boyer, G. L. (2014). The Fate of Microcystins in the Environment and Challenges for Monitoring. Toxins, 6(12), 3354-3387. https://doi.org/10.3390/toxins6123354