The Possible Role of Mycotoxins in the Pathogenesis of Endometrial Cancer

 ,
,  , , , and
, , , and
Abstract
1. Introduction
2. Results
2.1. Investigated Patient Groups
2.2. Biophysical, Hormonal, Clinical-Biochemical Parameters, and Endometrium Thickness
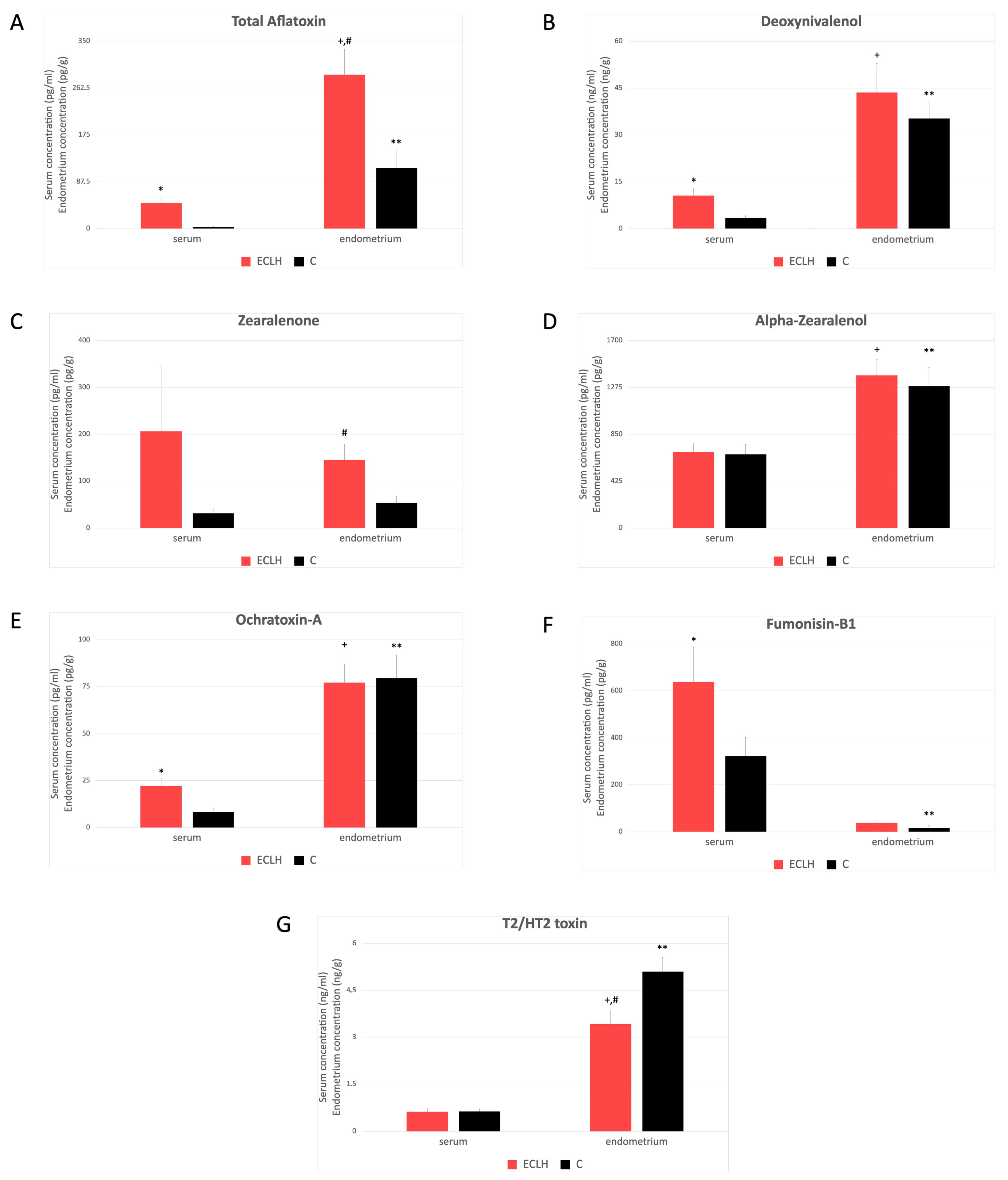
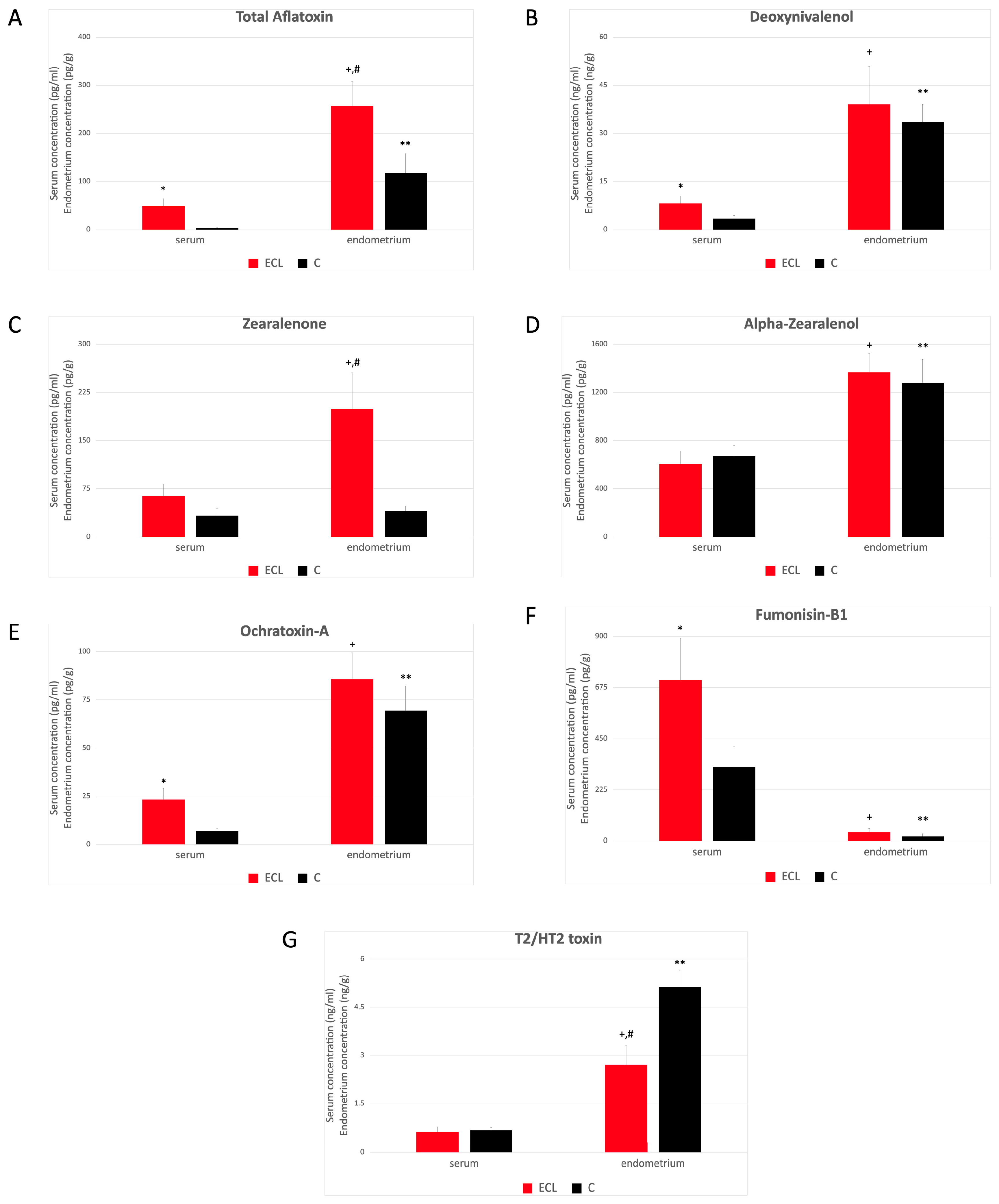
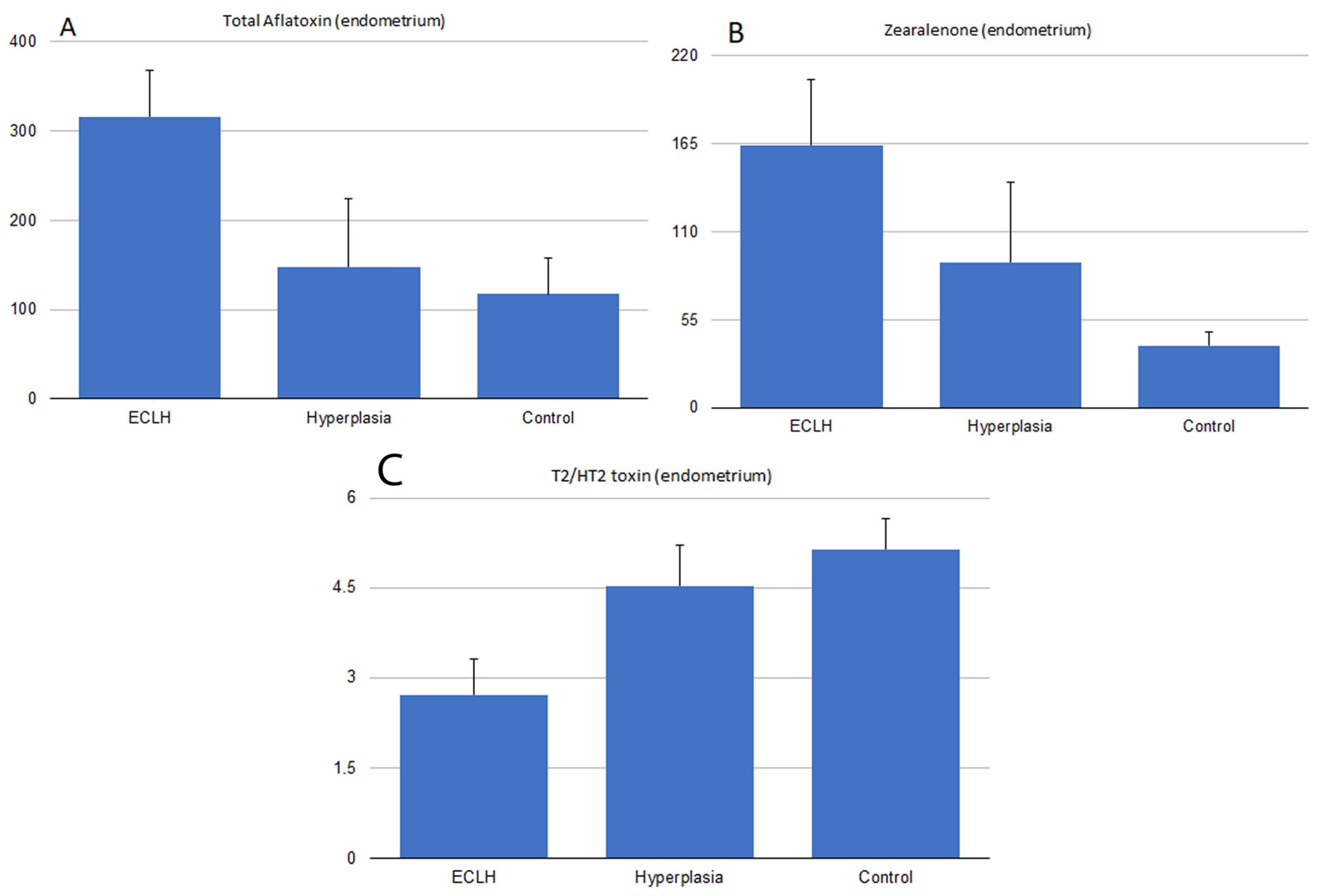
2.3. Serum and Endometrial Tissue Mycotoxin Levels
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Patients and Sample Collection
5.2. Experimental Setup on Samples
5.3. Mycotoxin Analyses
5.3.1. Zearalenone Assay
5.3.2. α-Zearalenol
5.3.3. Ochratoxin-A
5.3.4. Total Aflatoxin and Deoxynivalenol
5.3.5. Fumonisin B1
5.3.6. T2/HT2-Toxin Analyses
5.4. Hormone Analyses
5.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
References
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global Mycotoxin Occurrence in Feed: A Ten-Year Survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef] [PubMed]
- Cimbalo, A.; Alonso-Garrido, M.; Font, G.; Manyes, L. Toxicity of mycotoxins in vivo on vertebrate organisms: A review. Food Chem. Toxicol. 2020, 137, 111161. [Google Scholar] [CrossRef] [PubMed]
- Buszewska-Forajta, M. Mycotoxins, invisible danger of feedstuff with toxic effect on animals. Toxicon 2020, 182, 34–53. [Google Scholar] [CrossRef] [PubMed]
- Ekwomadu, T.; Mwanza, M.; Musekiwa, A. Mycotoxin-Linked Mutations and Cancer Risk: A Global Health Issue. Int. J. Environ. Res. Public Health 2022, 19, 7754. [Google Scholar] [CrossRef] [PubMed]
- Kobets, T.; Smith, B.P.C.; Williams, G.M. Food-Borne Chemical Carcinogens and the Evidence for Human Cancer Risk. Foods 2022, 11, 2828. [Google Scholar] [CrossRef] [PubMed]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens-the IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, S.F.; Bao, W. Molecular subtypes of endometrial cancer: Implications for adjuvant treatment strategies. Int. J. Gynaecol. Obstet. 2024, 164, 436–459. [Google Scholar] [CrossRef] [PubMed]
- Alexa, M.; Hasenburg, A.; Battista, M.J. The TCGA Molecular Classification of Endometrial Cancer and Its Possible Impact on Adjuvant Treatment Decisions. Cancers 2021, 13, 1478. [Google Scholar] [CrossRef] [PubMed]
- Bianco, B.; Barbosa, C.P.; Trevisan, C.M.; Laganà, A.S.; Montagna, E. Endometrial cancer: A genetic point of view. Transl. Cancer Res. 2020, 9, 7706–7715. [Google Scholar] [CrossRef]
- Kalinina, O.; Marchio, A.; Urbanskii, A.I.; Tarkova, A.B.; Rebbani, K.; Granov, D.A.; Dejean, A.; Generalov, M.I.; Pineau, P. Somatic changes in primary liver cancer in Russia: A pilot study. Mutat. Res. 2013, 755, 90–99. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Wu, R.Y.; Zhao, Y.; Xu, C.S.; Zhang, W.D.; Ge, W.; Liu, J.; Sun, Z.Y.; Zou, S.H.; Shen, W. Ochratoxin A exposure decreased sperm motility via the AMPK and PTEN signaling pathways. Toxicol. Appl. Pharmacol. 2018, 340, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Song, T.; Yang, W.; Huang, L.; Yang, Z.; Jiang, S. Zearalenone exposure affects the Wnt/β-catenin signaling pathway and related genes of porcine endometrial epithelial cells in vitro. Anim. Biosci. 2021, 34, 993–1005. [Google Scholar] [CrossRef] [PubMed]
- Vermij, L.; Léon-Castillo, A.; Singh, N.; Powell, M.E.; Edmondson, R.J.; Genestie, C.; Khaw, P.; Pyman, J.; McLachlin, C.M.; Ghatage, P.; et al. p53 immunohistochemistry in endometrial cancer: Clinical and molecular correlates in the PORTEC-3 trial. Mod. Pathol. 2022, 35, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Gouas, D.; Shi, H.; Hainaut, P. The aflatoxin-induced TP53 mutation at codon 249 (R249S): Biomarker of exposure, early detection and target for therapy. Cancer Lett. 2009, 286, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Van Vleet, T.R.; Watterson, T.L.; Klein, P.J.; Coulombe, R.A., Jr. Aflatoxin B1 Alters the Expression of p53 in Cytochrome P450-Expressing Human Lung Cells. Toxicol. Sci. 2005, 89, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Pajewska, M.; Łojko, M.; Cendrowski, K.; Sawicki, W.; Kowalkowski, T.; Buszewski, B.; Gadzała-Kopciuch, R. The determination of zearalenone and its major metabolites in endometrial cancer tissues. Anal. Bioanal. Chem. 2018, 410, 1571–1582. [Google Scholar] [CrossRef] [PubMed]
- Shmukler, M.; Elert, G. Density of blood. Phys. Factb. 2004, 4, 177–181. [Google Scholar]
- Guerre, P. Mycotoxin and Gut Microbiota Interactions. Toxins 2020, 12, 769. [Google Scholar] [CrossRef] [PubMed]
- Abu-Rustum, N.; Yashar, C.; Arend, R.; Barber, E.; Bradley, K.; Brooks, R.; Campos, S.M.; Chino, J.; Chon, H.S.; Chu, C.; et al. Uterine Neoplasms, Version 1.2023, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2023, 21, 181–209. [Google Scholar] [CrossRef]
- Cheymol, G. Drug pharmacokinetics in the obese. Fundam. Clin. Pharmacol. 1988, 2, 239–256. [Google Scholar] [CrossRef]
- Lootens, O.; De Boevre, M.; Ning, J.; Gasthuys, E.; Van Bocxlaer, J.; De Saeger, S.; Vermeulen, A. Building a Human Physiologically Based Pharmacokinetic Model for Aflatoxin B1 to Simulate Interactions with Drugs. Pharmaceutics 2023, 15, 894. [Google Scholar] [CrossRef] [PubMed]
- Fodor, J.; Balogh, K.; Weber, M.; Mézes, M.; Kametler, L.; Pósa, R.; Mamet, R.; Bauer, J.; Horn, P.; Kovács, F.; et al. Absorption, distribution, and elimination of fumonisin B1 metabolites in weaned piglets. Food Addit. Contam. 2008, 25, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Utsunomiya, H.; Yaegashi, N.; Sasano, H. Biological roles of estrogen and progesterone in human endometrial carcinoma–new developments in potential endocrine therapy for endometrial cancer. Endocr. J. 2007, 54, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Berstein, L.M.; Tchernobrovkina, A.E.; Gamajunova, V.B.; Kovalevskij, A.J.; Vasilyev, D.A.; Chepik, O.F.; Turkevitch, E.A.; Tsyrlina, E.V.; Maximov, S.J.; Ashrafian, L.A.; et al. Tumor estrogen content and clinico-morphological and endocrine features of endometrial cancer. J. Cancer Res. Clin. Oncol. 2003, 129, 245–249. [Google Scholar] [CrossRef]
- Poutanen, M.; Penning, T.M. Biology and clinical relevance of Hydroxysteroid (17beta) dehydrogenase enzymes. Mol. Cell. Endocrinol. 2019, 489, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Aljazzar, A.; El-Ghareeb, W.R.; Darwish, W.S.; Abdel-Raheem, S.M.; Ibrahim, A.M. Content of total aflatoxin, lead, and cadmium in the bovine meat and edible offal: Study of their human dietary intake, health risk assessment, and molecular biomarkers. Environ. Sci. Pollut. Res. Int. 2021, 28, 61225–61234. [Google Scholar] [CrossRef]
- Gramantieri, L.; Gnudi, F.; Vasuri, F.; Mandrioli, D.; Fornari, F.; Tovoli, F.; Suzzi, F.; Vornoli, A.; D’Errico, A.; Piscaglia, F.; et al. Aflatoxin B1 DNA-Adducts in Hepatocellular Carcinoma from a Low Exposure Area. Nutrients 2022, 14, 1652. [Google Scholar] [CrossRef]
- Amuzie, C.J.; Harkema, J.R.; Pestka, J.J. Tissue distribution and proinflammatory cytokine induction by the trichothecene deoxynivalenol in the mouse: Comparison of nasal vs. oral exposure. Toxicology 2008, 248, 39–44. [Google Scholar] [CrossRef]
- Soto, J.B.; Ruiz, M.J.; Manyes, L.; Juan-García, A. Blood, breast milk and urine: Potential biomarkers of exposure and estimated daily intake of ochratoxin A: A review. Food Addit. Contam. Part A 2016, 33, 313–328. [Google Scholar] [CrossRef]
- Schneweis, I.; Meyer, K.; Ritzmann, M.; Hoffmann, P.; Dempfle, L.; Bauer, J. Influence of organically or conventionally produced wheat on health, performance and mycotoxin residues in tissues and bile of growing pigs. Arch. Anim. Nutr. 2005, 59, 155–163. [Google Scholar] [CrossRef]
- Matsumoto, H.; Ito, T.; Ueno, Y. Toxicological approaches to the metabolities of fusaria. XII. Fate and distribution of T-2 toxin in mice. Jpn. J. Exp. Med. 1978, 48, 393–399. [Google Scholar] [PubMed]
- Yang, L.; Zhao, Z.; Deng, Y.; Zhou, Z.; Hou, J. Toxicity induced by F. poae-contaminated feed and the protective effect of Montmorillonite supplementation in broilers. Food Chem. Toxicol. 2014, 74, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Gadzała-Kopciuch, R.; Cendrowski, K.; Cesarz, A.; Kiełbasa, P.; Buszewski, B. Determination of zearalenone and its metabolites in endometrial cancer by coupled separation techniques. Anal. Bioanal. Chem. 2011, 401, 2069–2078. [Google Scholar] [CrossRef] [PubMed]
- Stockmann-Juvala, H.; Savolainen, K. A review of the toxic effects and mechanisms of action of fumonisin B1. Hum. Exp. Toxicol. 2008, 27, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, Y.; Wakai, T.; Kubota, M.; Osawa, M.; Sanpei, A.; Fujimaki, S. Mycotoxins are conventional and novel risk biomarkers for hepatocellular carcinoma. World J. Gastroenterol. 2013, 19, 2587–2590. [Google Scholar] [CrossRef] [PubMed]
- Renzulli, C.; Galvano, F.; Pierdomenico, L.; Speroni, E.; Guerra, M.C. Effects of rosmarinic acid against aflatoxin B1 and ochratoxin-A-induced cell damage in a human hepatoma cell line (Hep G2). J. Appl. Toxicol. 2004, 24, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, V.; Darroudi, F.; Uhl, M.; Steinkellner, H.; Gann, M.; Majer, B.J.; Eisenbauer, M.; Knasmüller, S. Genotoxic effects of ochratoxin A in human-derived hepatoma (HepG2) cells. Food Chem. Toxicol. 2002, 40, 1085–1090. [Google Scholar] [CrossRef] [PubMed]
- Bouaziz, C.; Sharaf El Dein, O.; El Golli, E.; Abid-Essefi, S.; Brenner, C.; Lemaire, C.; Bacha, H. Different apoptotic pathways induced by zearalenone, T-2 toxin and ochratoxin A in human hepatoma cells. Toxicology 2008, 254, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Grosso, F.; Saíd, S.; Mabrouk, I.; Fremy, J.M.; Castegnaro, M.; Jemmali, M.; Dragacci, S. New data on the occurrence of ochratoxin A in human sera from patients affected or not by renal diseases in Tunisia. Food Chem. Toxicol. 2003, 41, 1133–1140. [Google Scholar] [CrossRef]
- Aslam, M.; Rivzi, S.A.; Beg, A.E.; Blaszkewicz, M.; Golka, K.; Degen, G.H. Analysis of ochratoxin A blood levels in bladder cancer cases and healthy persons from Pakistan. J. Toxicol. Environ. Health A 2012, 75, 1176–1184. [Google Scholar] [CrossRef]
- Tomaszewski, J.; Miturski, R.; Semczuk, A.; Kotarski, J.; Jakowicki, J. Tissue zearalenone concentration in normal, hyperplastic and neoplastic human endometrium. Ginekol. Pol. 1998, 69, 363–366. [Google Scholar]
- Mousavi Khaneghah, A.; Fakhri, Y.; Raeisi, S.; Armoon, B.; Sant’Ana, A.S. Prevalence and concentration of ochratoxin A, zearalenone, deoxynivalenol and total aflatoxin in cereal-based products: A systematic review and meta-analysis. Food Chem. Toxicol. 2018, 118, 830–848. [Google Scholar] [CrossRef]
- Flasch, M.; Bueschl, C.; Del Favero, G.; Adam, G.; Schuhmacher, R.; Marko, D.; Warth, B. Elucidation of xenoestrogen metabolism by non-targeted, stable isotope-assisted mass spectrometry in breast cancer cells. Environ. Int. 2022, 158, 106940. [Google Scholar] [CrossRef]
- Rasooly, R.; Hernlem, B.; He, X.; Friedman, M. Non-linear relationships between aflatoxin B1 levels and the biological response of monkey kidney vero cells. Toxins 2013, 5, 1447–1461. [Google Scholar] [CrossRef]
- Wang, J.S.; Groopman, J.D. DNA damage by mycotoxins. Mutat. Res. 1999, 424, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, D.; Zhang, Z.; Zhu, W.; Li, W.; Zhao, J.; Xing, X.; He, Z.; Wang, S.; Wang, F.; et al. Persistent phosphorylation at specific H3 serine residues involved in chemical carcinogen-induced cell transformation. Mol. Carcinog. 2017, 56, 1449–1460. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Wang, J.; Huang, S.; Jiang, X.; Li, Y.; Wu, W.; Zhang, X. The G(2) phase arrest induced by sterigmatocystin is dependent on hMLH1- ERK/p38-p53 pathway in human esophageal epithelium cells in vitro. Food Chem. Toxicol. 2018, 115, 205–211. [Google Scholar] [CrossRef]
- Dong, L.; Lu, D.; Chen, R.; Lin, Y.; Zhu, H.; Zhang, Z.; Cai, S.; Cui, P.; Song, G.; Rao, D.; et al. Proteogenomic characterization identifies clinically relevant subgroups of intrahepatic cholangiocarcinoma. Cancer Cell 2022, 40, 70–87.e15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, J.; Cui, J.; Xing, L.; Wang, J.; Yan, X.; Zhang, X. ERK and p38 MAPK signaling pathways are involved in ochratoxin A-induced G2 phase arrest in human gastric epithelium cells. Toxicol. Lett. 2012, 209, 186–192. [Google Scholar] [CrossRef]
- Szőke, Z.; Babarczi, B.; Mézes, M.; Lakatos, I.; Poór, M.; Fliszár-Nyúl, E.; Oldal, M.; Czéh, Á.; Bodó, K.; Nagyéri, G.; et al. Analysis and Comparison of Rapid Methods for the Determination of Ochratoxin a Levels in Organs and Body Fluids Obtained from Exposed Mice. Toxins 2022, 14, 634. [Google Scholar] [CrossRef]
- Falkauskas, R.; Bakutis, B.; Jovaišienė, J.; Vaičiulienė, G.; Gerulis, G.; Kerzienė, S.; Jacevičienė, I.; Jacevičius, E.; Baliukonienė, V. Zearalenone and Its Metabolites in Blood Serum, Urine, and Milk of Dairy Cows. Animals 2022, 12, 1651. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| ECL (n = 12) | ECLH (n = 18) | Control (n = 21) | |
|---|---|---|---|
| BMI (Mean ± SEM, kg/m2) | 34.25 ± 2.141 | 33.97 ± 1.701 | 30.43 ± 1.415 |
| ALT (GPT, Mean ± SEM, IU/L) | 19.45 ± 2.977 | 18.36 ± 2.486 | 25.07 ± 2.958 |
| TSH (Mean ± SEM, mIU/L) | 2.139 ± 0.7526 | 2.21 ± 0.5885 | 1.33 ± 0.5 |
| Progesterone (Mean ± SEM, ng/mL) | 0.27 ± 0.0932 | 0.41 ± 0.1639 | 0.41 ± 0.1544 |
| Estrone (Mean ± SEM, pg/mL) | 45.30 ± 6.075 ab | 59.12 ± 13.9 b,* | 27.90 ± 6.209 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Unicsovics, M.; Molnár, Z.; Mézes, M.; Posta, K.; Nagyéri, G.; Várbíró, S.; Ács, N.; Sára, L.; Szőke, Z. The Possible Role of Mycotoxins in the Pathogenesis of Endometrial Cancer. Toxins 2024, 16, 236. https://doi.org/10.3390/toxins16060236
Unicsovics M, Molnár Z, Mézes M, Posta K, Nagyéri G, Várbíró S, Ács N, Sára L, Szőke Z. The Possible Role of Mycotoxins in the Pathogenesis of Endometrial Cancer. Toxins. 2024; 16(6):236. https://doi.org/10.3390/toxins16060236
Chicago/Turabian StyleUnicsovics, Márkó, Zsófia Molnár, Miklós Mézes, Katalin Posta, György Nagyéri, Szabolcs Várbíró, Nándor Ács, Levente Sára, and Zsuzsanna Szőke. 2024. "The Possible Role of Mycotoxins in the Pathogenesis of Endometrial Cancer" Toxins 16, no. 6: 236. https://doi.org/10.3390/toxins16060236
APA StyleUnicsovics, M., Molnár, Z., Mézes, M., Posta, K., Nagyéri, G., Várbíró, S., Ács, N., Sára, L., & Szőke, Z. (2024). The Possible Role of Mycotoxins in the Pathogenesis of Endometrial Cancer. Toxins, 16(6), 236. https://doi.org/10.3390/toxins16060236