Sensitive Detection and Differentiation of Biologically Active Ricin and Abrin in Complex Matrices via Specific Neutralizing Antibody-Based Cytotoxicity Assay


Abstract
1. Introduction
2. Results and Discussion
2.1. HeLa Cells Exhibited the Highest Sensitivity to Both Ricin and Abrin Exposures
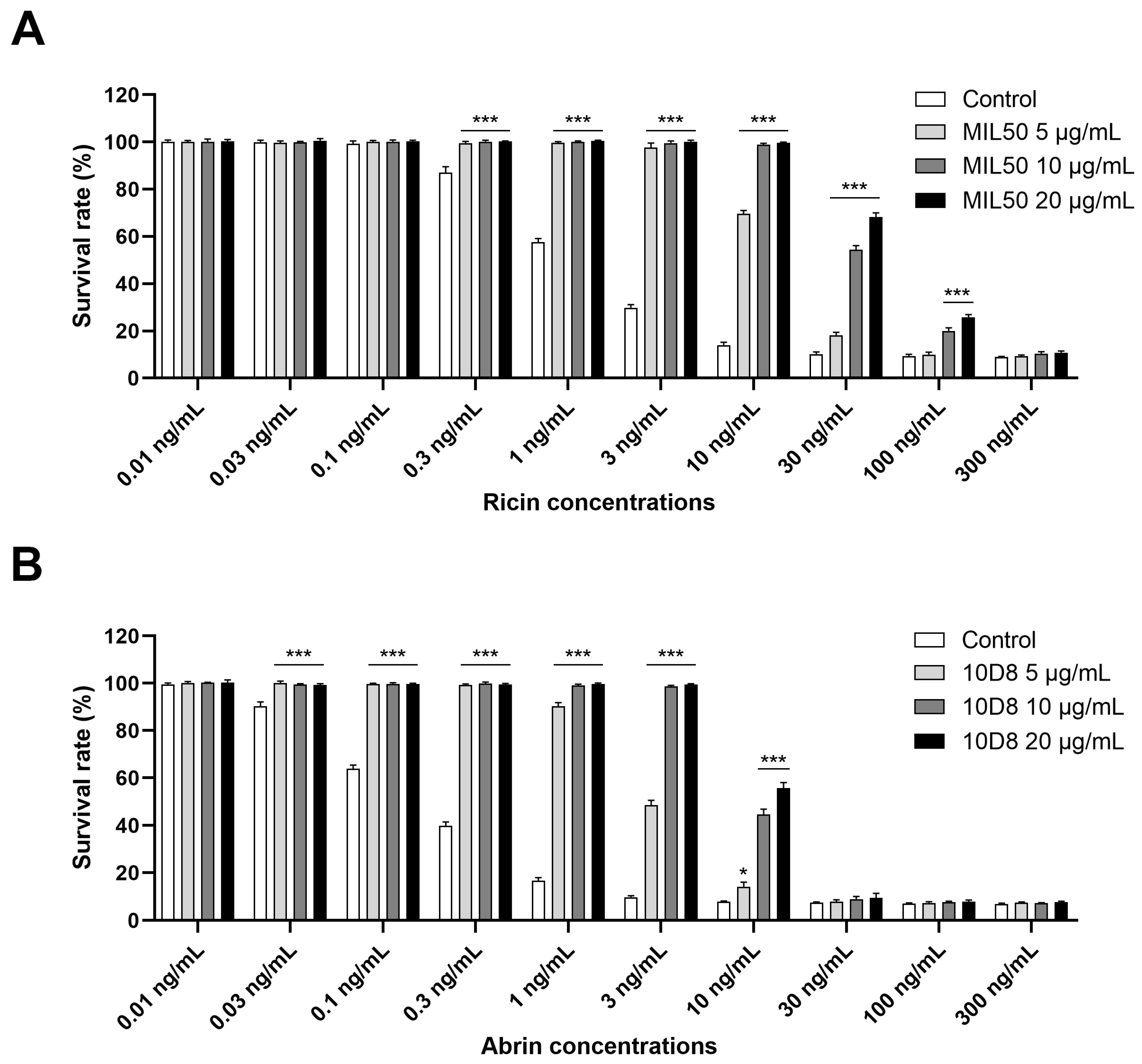
2.2. Optimization of Neutralizing Antibodies Concentrations for Ricin and Abrin-Specific Detection
2.3. Assessment of Neutralizing Antibodies’ Cross-Reactivity
2.4. Impact of Complex Matrices on Cell Growth and Toxin Detection
2.5. Detection of Ricin and Abrin from the Samples of OPCW Biotoxin Exercises
3. Conclusions
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Cytotoxicity Evaluation
4.4. Antibody Treatment
4.5. Analysis of Complex Matrix Samples
4.6. Analysis of OPCW Biotoxin Exercise Samples
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Felder, E.; Mossbrugger, I.; Lange, M.; Wolfel, R. Simultaneous detection of ricin and abrin DNA by real-time PCR (qPCR). Toxins 2012, 4, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Stirpe, F.; Battelli, M.G. Ribosome-inactivating proteins: Progress and problems. Cell Mol. Life Sci. 2006, 63, 1850–1866. [Google Scholar] [CrossRef] [PubMed]
- Janik, E.; Ceremuga, M.; Saluk-Bijak, J.; Bijak, M. Biological Toxins as the Potential Tools for Bioterrorism. Int. J. Mol. Sci. 2019, 20, 1181. [Google Scholar] [CrossRef] [PubMed]
- Bozza, W.P.; Tolleson, W.H.; Rivera Rosado, L.A.; Zhang, B. Ricin detection: Tracking active toxin. Biotechnol. Adv. 2015, 33, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Dorner, B.G.; Zeleny, R.; Harju, K.; Hennekinne, J.-A.; Vanninen, P.; Schimmel, H.; Rummel, A. Biological toxins of potential bioterrorism risk: Current status of detection and identification technology. TrAC Trends Anal. Chem. 2016, 85, 89–102. [Google Scholar] [CrossRef]
- Robb, C.S. The analysis of abrin in food and beverages. TrAC Trends Anal. Chem. 2015, 67, 100–106. [Google Scholar] [CrossRef]
- Kalb, S.R.; Schieltz, D.M.; Becher, F.; Astot, C.; Fredriksson, S.A.; Barr, J.R. Recommended Mass Spectrometry-Based Strategies to Identify Ricin-Containing Samples. Toxins 2015, 7, 4881–4894. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.H.; Yang, Y.; Geng, S.; Cheng, X.; Yu, H.L.; Liu, C.C.; Liu, S.L. Rapid Differential Detection of Abrin Isoforms by an Acetonitrile- and Ultrasound-Assisted On-Bead Trypsin Digestion Coupled with LC-MS/MS Analysis. Toxins 2021, 13, 358. [Google Scholar] [CrossRef]
- Simon, S.; Worbs, S.; Avondet, M.A.; Tracz, D.M.; Dano, J.; Schmidt, L.; Volland, H.; Dorner, B.G.; Corbett, C.R. Recommended Immunological Assays to Screen for Ricin-Containing Samples. Toxins 2015, 7, 4967–4986. [Google Scholar] [CrossRef]
- He, X.; Patfield, S.; Cheng, L.W.; Stanker, L.H.; Rasooly, R.; McKeon, T.A.; Zhang, Y.; Brandon, D.L. Detection of Abrin Holotoxin Using Novel Monoclonal Antibodies. Toxins 2017, 9, 386. [Google Scholar] [CrossRef]
- Worbs, S.; Kampa, B.; Skiba, M.; Hansbauer, E.M.; Stern, D.; Volland, H.; Becher, F.; Simon, S.; Dorner, M.B.; Dorner, B.G. Differentiation, Quantification and Identification of Abrin and Abrus precatorius Agglutinin. Toxins 2021, 13, 284. [Google Scholar] [CrossRef] [PubMed]
- Becher, F.; Duriez, E.; Volland, H.; Tabet, J.C.; Ezan, E. Detection of functional ricin by immunoaffinity and liquid chromatography-tandem mass spectrometry. Anal. Chem. 2007, 79, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Taisne, C.; Mahtal, N.; Forrester, A.; Lussignol, M.; Cintrat, J.C.; Esclatine, A.; Gillet, D.; Barbier, J. Autophagic Degradation Is Involved in Cell Protection against Ricin Toxin. Toxins 2023, 15, 304. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, V.L.; Nilles, J.M.; Rice, J.S.; Connell, T.R.; Schenning, A.M.; Reilly, L.M.; Durst, H.D. Ricin activity assay by direct analysis in real time mass spectrometry detection of adenine release. Anal. Chem. 2010, 82, 798–800. [Google Scholar] [CrossRef] [PubMed]
- Hale, M.L. Microtiter-based assay for evaluating the biological activity of ribosome-inactivating proteins. Pharmacol. Toxicol. 2001, 88, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Dawson, R.M.; Paddle, B.M.; Alderton, M.R. Characterization of the Asialofetuin microtitre plate-binding assay for evaluating inhibitors of ricin lectin activity. J. Appl. Toxicol. 1999, 19, 307–312. [Google Scholar] [CrossRef]
- Brzezinski, J.L.; Craft, D.L. Evaluation of an in vitro bioassay for the detection of purified ricin and castor bean in beverages and liquid food matrices. J. Food Prot. 2007, 70, 2377–2382. [Google Scholar] [CrossRef] [PubMed]
- Wahome, P.G.; Bai, Y.; Neal, L.M.; Robertus, J.D.; Mantis, N.J. Identification of small-molecule inhibitors of ricin and shiga toxin using a cell-based high-throughput screen. Toxicon 2010, 56, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Stechmann, B.; Bai, S.K.; Gobbo, E.; Lopez, R.; Merer, G.; Pinchard, S.; Panigai, L.; Tenza, D.; Raposo, G.; Beaumelle, B.; et al. Inhibition of retrograde transport protects mice from lethal ricin challenge. Cell 2010, 141, 231–242. [Google Scholar] [CrossRef]
- Pauly, D.; Worbs, S.; Kirchner, S.; Shatohina, O.; Dorner, M.B.; Dorner, B.G. Real-time cytotoxicity assay for rapid and sensitive detection of ricin from complex matrices. PLoS ONE 2012, 7, e35360. [Google Scholar] [CrossRef]
- Oda, T.; Komatsu, N.; Muramatsu, T. Cell lysis induced by ricin D and ricin E in various cell lines. Biosci. Biotechnol. Biochem. 1997, 61, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Saxena, N.; Phatak, P.; Chauhan, V. Differential toxicity of abrin in human cell lines of different organ origin. Toxicol. In Vitro 2022, 78, 105250. [Google Scholar] [CrossRef] [PubMed]
- Rasooly, R.; He, X. Sensitive bioassay for detection of biologically active ricin in food. J. Food Prot. 2012, 75, 951–954. [Google Scholar] [CrossRef] [PubMed]
- Pasetto, M.; Barison, E.; Castagna, M.; Della Cristina, P.; Anselmi, C.; Colombatti, M. Reductive activation of type 2 ribosome-inactivating proteins is promoted by transmembrane thioredoxin-related protein. J. Biol. Chem. 2012, 287, 7367–7373. [Google Scholar] [CrossRef]
- Tolleson, W.H.; Jackson, L.S.; Triplett, O.A.; Aluri, B.; Cappozzo, J.; Banaszewski, K.; Chang, C.W.; Nguyen, K.T. Chemical inactivation of protein toxins on food contact surfaces. J. Agric. Food Chem. 2012, 60, 6627–6640. [Google Scholar] [CrossRef] [PubMed]
- Sowa-Rogozinska, N.; Sominka, H.; Nowakowska-Golacka, J.; Sandvig, K.; Slominska-Wojewodzka, M. Intracellular Transport and Cytotoxicity of the Protein Toxin Ricin. Toxins 2019, 11, 350. [Google Scholar] [CrossRef]
- Makdasi, E.; Laskar, O.; Milrot, E.; Schuster, O.; Shmaya, S.; Yitzhaki, S. Whole-Cell Multiparameter Assay for Ricin and Abrin Activity-Based Digital Holographic Microscopy. Toxins 2019, 11, 174. [Google Scholar] [CrossRef] [PubMed]
- Sandvig, K.; Olsnes, S.; Pihl, A. Kinetics of binding of the toxic lectins abrin and ricin to surface receptors of human cells. J. Biol. Chem. 1976, 251, 3977–3984. [Google Scholar] [CrossRef] [PubMed]
- Leonard, J.E.; Grothaus, C.D.; Taetle, R. Ricin binding and protein synthesis inhibition in human hematopoietic cell lines. Blood 1988, 72, 1357–1363. [Google Scholar] [CrossRef]
- Chan, L.N.; Li, J.S.; Liu, S.Y. Differential effects of abrin on normal and tumor cells. J. Cell Physiol. 1985, 123, 132–138. [Google Scholar] [CrossRef]
- Nagatsuka, T.; Uzawa, H.; Ohsawa, I.; Seto, Y.; Nishida, Y. Use of lactose against the deadly biological toxin ricin. ACS Appl. Mater. Interfaces 2010, 2, 1081–1085. [Google Scholar] [CrossRef] [PubMed]
- Rasooly, R.; He, X.; Friedman, M. Milk inhibits the biological activity of ricin. J. Biol. Chem. 2012, 287, 27924–27929. [Google Scholar] [CrossRef]
- Lumor, S.E.; Deen, B.D.; Ronningen, I.; Smith, K.; Fredrickson, N.R.; Diez-Gonzalez, F.; Labuza, T.P. Assessment of the inhibition of ricin toxicity by lactose in milk. J. Food Prot. 2013, 76, 2037–2039. [Google Scholar] [CrossRef]
- Brandon, D.L.; Adams, L.M. Milk matrix effects on antibody binding analyzed by enzyme-linked immunosorbent assay and biolayer interferometry. J. Agric. Food Chem. 2015, 63, 3593–3598. [Google Scholar] [CrossRef] [PubMed]
- Hegde, R.; Maiti, T.K.; Podder, S.K. Purification and characterization of three toxins and two agglutinins from Abrus precatorius seed by using lactamyl-Sepharose affinity chromatography. Anal. Biochem. 1991, 194, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Xie, J.; Shao, N.; Yan, Y. The DNA aptamers that specifically recognize ricin toxin are selected by two in vitro selection methods. Electrophoresis 2006, 27, 1303–1311. [Google Scholar] [CrossRef]
- Luo, L.; Yang, J.; Li, Z.; Xu, H.; Guo, L.; Wang, L.; Wang, Y.; Luo, L.; Wang, J.; Zhang, P.; et al. Label-free differentiation and quantification of ricin, abrin from their agglutinin biotoxins by surface plasmon resonance. Talanta 2022, 238, 122860. [Google Scholar] [CrossRef]
- Li, Z.; Xu, H.; Ma, B.; Luo, L.; Guo, L.; Zhang, P.; Zhao, Y.; Wang, L.; Xie, J. Neutralizing Monoclonal Antibody, mAb 10D8, Is an Effective Detoxicant against Abrin-a Both In Vitro and In Vivo. Toxins 2022, 14, 164. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Ricin IC50 (ng/mL) | Ricin LOD (ng/mL) | Abrin IC50 (ng/mL) | Abrin LOD (ng/mL) |
|---|---|---|---|---|
| HeLa | 1.18 | 0.30 | 0.14 | 0.03 |
| HepG2 | 130.20 | 10.00 | 1.30 | 0.30 |
| A549 | >300.00 | 3.00 | 67.61 | 1.00 |
| HEK293 | 2.57 | 0.30 | 2.34 | 0.30 |
| HCT-8 | 74.99 | 1.00 | 36.52 | 1.00 |
| BGC-823 | 250.40 | 10.00 | 12.80 | 1.00 |
| HUVEC | 5.01 | 0.30 | 4.22 | 0.30 |
| AC16 | 3.16 | 0.30 | 2.70 | 0.30 |
| SH-SY5Y | >300.00 | 0.30 | 42.17 | 0.30 |
| No. | Sample Name | Spiking Chemicals (Concentration) | Matrices | Dilution Ratio | Detection Results |
|---|---|---|---|---|---|
| 3rd | BT18.1 | Abrin (150 μg/g) | Protein powder | 1:1000 | Abrin |
| BT18.2 | Ricin (15 μg/g) Abrus agglutinin (300 μg/g) * | Protein powder | 1:1000 | Ricin Abrin | |
| 4th | BT19.4 | Ricin (30 μg/mL) | Spray buffer | 1:20,000 | Ricin |
| 5th | BT20/PL01 | Ricin (16 μg/mL) | Human plasma | 1:1000 | Ricin |
| BT20/PL02 | Ricin (1 μg/mL) | Human plasma | 1:1000 | Ricin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Ma, B.; Gong, M.; Guo, L.; Wang, L.; Xu, H.; Xie, J. Sensitive Detection and Differentiation of Biologically Active Ricin and Abrin in Complex Matrices via Specific Neutralizing Antibody-Based Cytotoxicity Assay. Toxins 2024, 16, 237. https://doi.org/10.3390/toxins16060237
Li Z, Ma B, Gong M, Guo L, Wang L, Xu H, Xie J. Sensitive Detection and Differentiation of Biologically Active Ricin and Abrin in Complex Matrices via Specific Neutralizing Antibody-Based Cytotoxicity Assay. Toxins. 2024; 16(6):237. https://doi.org/10.3390/toxins16060237
Chicago/Turabian StyleLi, Zhi, Bo Ma, Mengqiang Gong, Lei Guo, Lili Wang, Hua Xu, and Jianwei Xie. 2024. "Sensitive Detection and Differentiation of Biologically Active Ricin and Abrin in Complex Matrices via Specific Neutralizing Antibody-Based Cytotoxicity Assay" Toxins 16, no. 6: 237. https://doi.org/10.3390/toxins16060237
APA StyleLi, Z., Ma, B., Gong, M., Guo, L., Wang, L., Xu, H., & Xie, J. (2024). Sensitive Detection and Differentiation of Biologically Active Ricin and Abrin in Complex Matrices via Specific Neutralizing Antibody-Based Cytotoxicity Assay. Toxins, 16(6), 237. https://doi.org/10.3390/toxins16060237