The Co-Occurrence of T-2 Toxin, Deoxynivalenol, and Fumonisin B1 Activated the Glutathione Redox System in the EU-Limiting Doses in Laying Hens


 , ,
, , 
Abstract
1. Introduction
2. Results
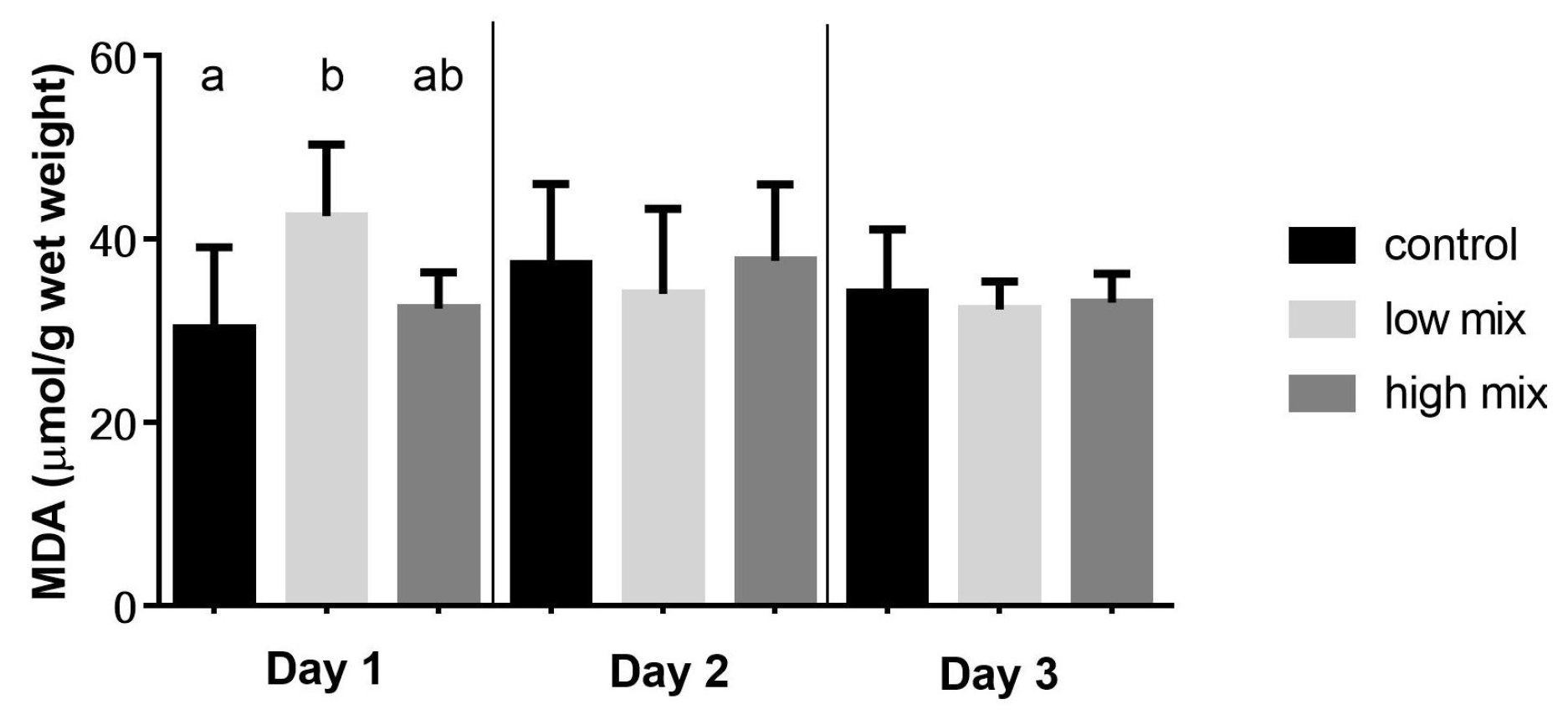
2.1. Results of the Lipid Peroxidation Parameters
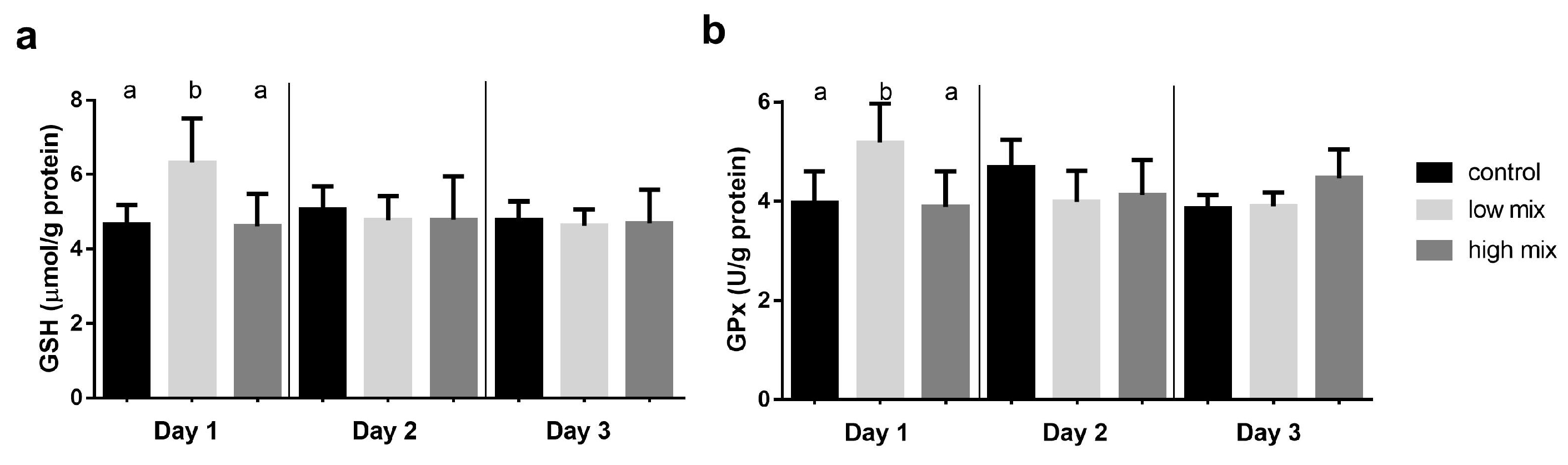
2.2. Results of the Glutathione Redox System Parameters
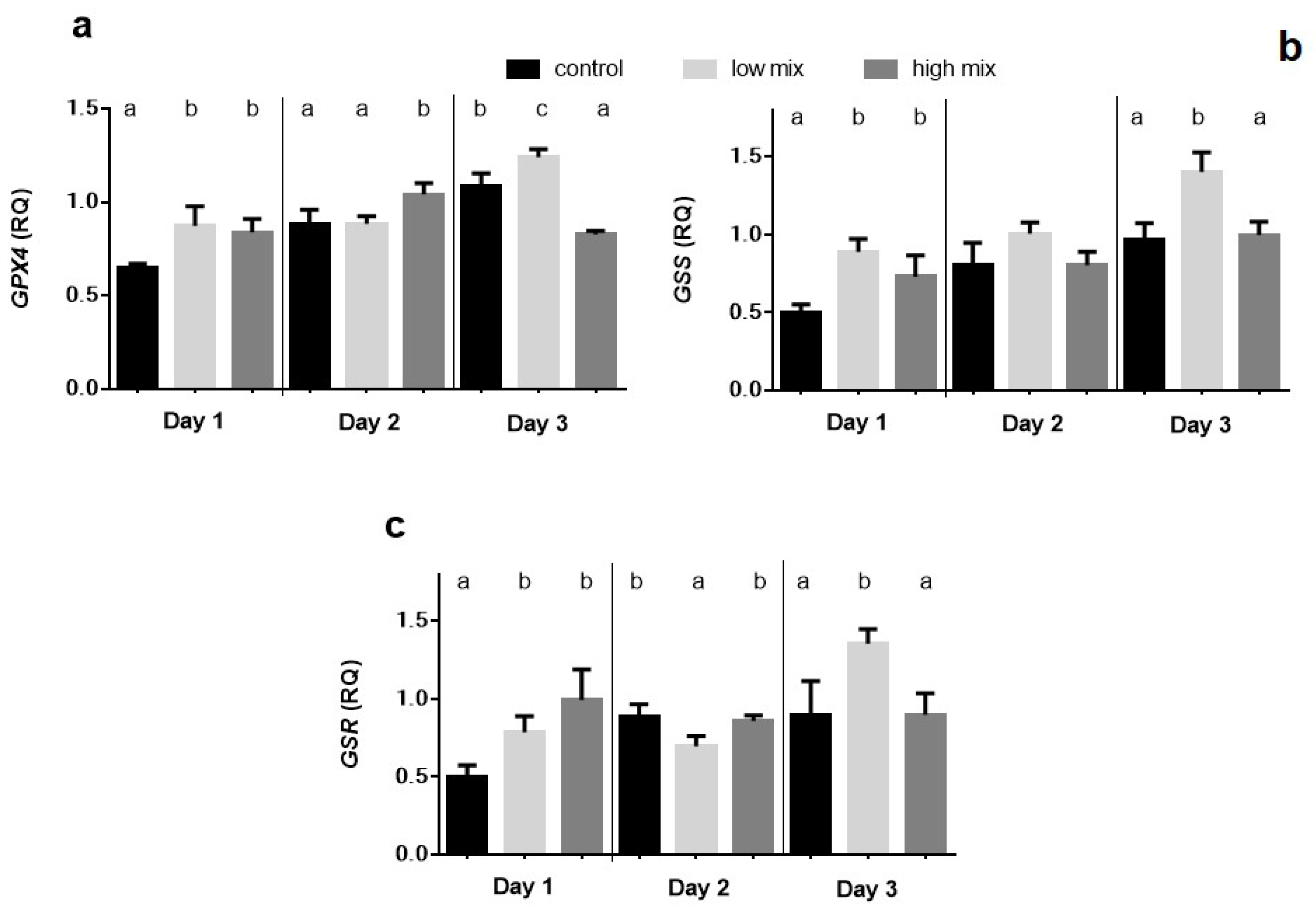
2.3. Relative Expression of GPX4, GSS, and GSR Genes
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Experimental design
5.2. Mycotoxin production
5.3. Biochemical and Gene Expression Analyses
5.4. Statistical Analysis
5.5. Ethical Issues
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tolosa, J.; Rodríguez-Carrasco, Y.; Ruiz, M.J.; Vila-Donat, P. Multi-mycotoxin occurrence in feed, metabolism and carry-over to animal-derived food products: A review. Food Chem. Toxicol. 2021, 158, 112661. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Gupta, A.; Mahato, D.K.; Pandhi, S.; Pandey, A.K.; Kargwal, R.; Mishra, S.; Suhag, R.; Sharma, N.; Saurabh, V.; et al. Aflatoxins in Cereals and Cereal-Based Products: Occurrence, Toxicity, Impact on Human Health, and Their Detoxification and Management Strategies. Toxins 2022, 14, 687. [Google Scholar] [CrossRef] [PubMed]
- Sokolović, M.; Garaj-Vrhovac, V.; Šimpraga, B. T-2 Toxin: Incidence and Toxicity in Poultry. Arch. Ind. Hyg. Toxicol. 2008, 59, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Awad, W.A.; Ghareeb, K.; Böhm, J.; Zentek, J. The Toxicological Impacts of the Fusarium Mycotoxin, Deoxynivalenol, in Poultry Flocks with Special Reference to Immunotoxicity. Toxins 2013, 5, 912–925. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-J.; Zhang, G.; Xue, B.; Ding, Q.; Han, L.; Huang, J.-C.; Wu, F.; Li, C.; Yang, C. Toxicity and detoxification of T-2 toxin in poultry. Food Chem. Toxicol. 2022, 169, 113392. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Toxicity, mechanisms and animal health risks. Anim. Feed. Sci. Technol. 2007, 137, 283–298. [Google Scholar] [CrossRef]
- Awad, W.A.; Ghareeb, K.; Bohm, J.; Razzazi, E.; Hellweg, P.; Zentek, J. The Impact of the Fusarium Toxin Deoxynivalenol (DON) on Poultry. Int. J. Poult. Sci. 2008, 9, 827–842. [Google Scholar] [CrossRef]
- Awad, W.A.; Ghareeb, K.; Böhm, J. The toxicity of Fusarium mycotoxin deoxynivalenol in poultry feeding. World’s Poult. Sci. J. 2012, 68, 651–668. [Google Scholar] [CrossRef]
- Fæste, C.K.; Solhaug, A.; Gaborit, M.; Pierre, F.; Massotte, D. Neurotoxic Potential of Deoxynivalenol in Murine Brain Cell Lines and Primary Hippocampal Cultures. Toxins 2022, 14, 48. [Google Scholar] [CrossRef]
- Kępińska-Pacelik, J.; Biel, W. Alimentary Risk of Mycotoxins for Humans and Animals. Toxins 2021, 13, 822. [Google Scholar] [CrossRef]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef] [PubMed]
- Broomhead, J.N.; Ledoux, D.R.; Bermudez, A.J.; Rottinghaus, G.E. Chronic Effects of Fumonisin B1 in Broilers and Turkeys Fed Dietary Treatments to Market Age. Poult. Sci. 2002, 81, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Weibking, T.S.; Ledoux, D.R.; Bermudez, A.J.; Rottinghaus, G.E. Individual and combined effects of feeding Fusarium monili-forme culture material, containing known levels of fumonisin B1, and aflatoxin B1 in the young turkey poult. Poult. Sci. 1994, 73, 1517–1525. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Luo, K.; Zhu, Q.; Peng, J.; Liu, C.; Wang, X.; Li, S.; Zhang, H. The natural occurrence, toxicity mechanisms and management strategies of Fumonisin B1: A review. Environ. Pollut. 2023, 320, 121065. [Google Scholar] [CrossRef]
- Strasser, A.; Carra, M.; Ghareeb, K.; Awad, G.W.; Böhm, J. Protective effects of antioxidants on deoxynivalenol-induced damage in murine lymphoma cells. Mycotoxin Res. 2013, 29, 203–208. [Google Scholar] [CrossRef]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of mycotoxin on immune response and consequences for pig health. Anim. Nutr. 2016, 2, 63–68. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, F.; Zhou, X.; Liu, M.; Zang, H.; Liu, X.; Shan, A.; Feng, X. Alleviation of Oral Exposure to Aflatoxin B1-Induced Renal Dysfunction, Oxidative Stress, and Cell Apoptosis in Mice Kidney by Curcumin. Antioxidants 2022, 11, 1082. [Google Scholar] [CrossRef]
- Surai, P.F.; Dvorska, J.E.; Sparks, N.H.C.; Jaques, K.A. Impact of mycotoxins on the body’s antioxidant defence. In Nutritional Biotechnology in the Feed and Food Industries, 2nd ed.; Lyons, T.P., Jaques, K.A., Eds.; Nottingham University Press: Nottingham, UK, 2002; pp. 131–142. [Google Scholar]
- Weber, M.; Balogh, K.; Fodor, J.; Erdélyi, M.; Ancsin, Z.; Mézes, M. Effect of T-2 and HT-2 Toxin during the Growing Period on Body Weight, Lipid Peroxide and Glutathione Redox Status of Broiler Chickens. Acta Veter. Brno 2010, 79, 27–31. [Google Scholar] [CrossRef]
- Gloire, G.; Legrand-Poels, S.; Piette, J. NF-κB activation by reactive oxygen species: Fifteen years later. Biochem. Pharmacol. 2006, 72, 1493–1505. [Google Scholar] [CrossRef]
- Smith, M.-C.; Madec, S.; Coton, E.; Hymery, N. Natural Co-Occurrence of Mycotoxins in Foods and Feeds and Their in vitro Combined Toxicological Effects. Toxins 2016, 4, 94. [Google Scholar] [CrossRef]
- Pelyhe, C.; Kövesi, B.; Zándoki, E.; Kovács, B.; Erdélyi, M.; Kulcsár, S.; Mézes, M.; Balogh, K. Multi-trichothecene mycotoxin exposure activates glutathione-redox system in broiler chicken. Toxicon 2018, 153, 53–57. [Google Scholar] [CrossRef]
- Szabó-Fodor, J.; Szabó, A.; Kócsó, D.; Marosi, K.; Bóta, B.; Kachlek, M.; Mézes, M.; Balogh, K.; Kövér, G.; Nagy, I.; et al. Interaction between the three frequently co-occurring Fusarium mycotoxins in rats. J. Anim. Physiol. Anim. Nutr. 2019, 103, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Kulcsár, S.; Kövesi, B.; Balogh, K.; Zándoki, E.; Ancsin, Z.; Márta, B.E.; Mézes, M. Effects of Fusarium Mycotoxin Exposure on Lipid Peroxidation and Glutathione Redox System in the Liver of Laying Hens. Antioxidants 2021, 10, 1313. [Google Scholar] [CrossRef] [PubMed]
- Balogh, K.; Bócsai, A.; Pelyhe, C.; Zándoki, E.; Erdélyi, M.; Szabó-Fodor, J.; Mézes, M. Effects of Long-term Feeding of Graded Levels of T-2 Toxin-contaminated Diets on Performance, Some Lipid Peroxide and Glutathione Redox Status Parameters of Broiler Chickens. J. Poult. Sci. 2015, 52, 176–182. [Google Scholar] [CrossRef]
- Poersch, A.B.; Trombetta, F.; Braga, A.C.M.; Boeira, S.P.; Oliveira, M.S.; Dilkin, P.; Mallmann, C.A.; Fighera, M.R.; Royes, L.; Furian, A. Involvement of oxidative stress in subacute toxicity induced by fumonisin B1 in broiler chicks. Veter. Microbiol. 2014, 174, 180–185. [Google Scholar] [CrossRef]
- Antonissen, G.; Van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Janssens, G.P.J.; De Baere, S.; Mountzouris, K.; Su, S.; Wong, E.A.; De Meulenaer, B.; et al. Mycotoxins Deoxynivalenol and Fumonisins Alter the Extrinsic Component of Intestinal Barrier in Broiler Chickens. J. Agric. Food Chem. 2015, 63, 10846–10855. [Google Scholar] [CrossRef]
- European Commission Recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. J. Eur. Union. 2007, L 118M, 1111–1113.
- Biomin World Mycotoxin Survey. 2020. Available online: https://www.biomin.net/downloads/2020-biomin-world-mycotoxin-survey-report (accessed on 20 December 2020).
- Fodor, J.; Németh, M.; Kametler, L.; Pósa, R.; Kovács, M.; Horn, P. Novel methods of Fusarium toxins’ production for toxico-logical experiments. Acta Agrar. Kaposváriensis 2006, 10, 277–285. [Google Scholar]
- Trebstein, A.; Seefelder, W.; Lauber, U.; Humpf, H.-U. Determination of T-2 and HT-2 Toxins in Cereals Including Oats after Immunoaffinity Cleanup by Liquid Chromatography and Fluorescence Detection. J. Agric. Food Chem. 2008, 56, 4968–4975. [Google Scholar] [CrossRef]
- Pussemier, L.; Pierard, J.-Y.; Anselme, M.; Tangni, E.K.; Motte, J.-C.; Larondelle, Y. Development and application of analytical methods for the determination of mycotoxins in organic and conventional wheat. Food Addit. Contam. 2006, 23, 1208–1218. [Google Scholar] [CrossRef]
- Shephard, G.S.; Sydenham, E.W.; Thiel, P.G.; Gelderblom, W.C.A. Quantitative Determination of Fumonisins B1 and B1 by High-Performance Liquid Chromatography with Fluorescence Detection. J. Liq. Chromatogr. 1990, 13, 2077–2087. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis (28.054), 14th ed.; Association of Official Analytical Chemists: Arlington, TX, USA, 1984; p. 243. [Google Scholar]
- Botsoglou, N.A.; Fletouris, D.J.; Papageorgiou, G.E.; Vassilopoulos, V.N.; Mantis, A.J.; Trakatellis, A.G. Rapid, Sensitive, and Specific Thiobarbituric Acid Method for Measuring Lipid Peroxidation in Animal Tissue, Food, and Feedstuff Samples. J. Agric. Food Chem. 1994, 42, 1931–1937. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound and non-protein sulfhydryl groups in tissues with Ellmann’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, R.A.; Burk, R.F. Glutathione peroxidase activity in selenium-deficient rat liver. Biochem. Biophys. Res. Commun. 1976, 71, 952–956. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Kendziorski, C.M.; Zhang, Y.; Lan, H.; Attie, A.D. The efficiency of pooling mRNA in microarray experiments. Biostatistics 2003, 4, 465–477. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Conjugated Dienes (OD 232 nm) | ||||
| Day 0 | Day 1 | Day 2 | Day 3 | |
| Control | 0.61 ± 0.03 | 0.60 ± 0.07 | 0.60 ± 0.05 | 0.65 ± 0.07 |
| Low mix | 0.60 ± 0.05 | 0.63 ± 0.13 | 0.57 ± 0.04 | |
| High mix | 0.60 ± 0.07 | 0.61 ± 0.09 | 0.59 ± 0.05 | |
| Conjugated trienes (OD 268 nm) | ||||
| Day 0 | Day 1 | Day 2 | Day 3 | |
| Control | 0.23 ± 0.03 | 0.22 ± 0.02 | 0.22 ± 0.02 | 0.23 ± 0.02 |
| Low mix | 0.21 ± 0.02 | 0.23 ± 0.05 | 0.20 ± 0.02 | |
| High mix | 0.22 ± 0.02 | 0.22 ± 0.03 | 0.22 ± 0.02 | |
| Group | T-2/HT-2 | DON/3-AcDON/15-AcDON | FB1 |
|---|---|---|---|
| Control | <0.02 | <0.02 | <0.02 |
| Low mix | 0.22/0.04 | 4.25 | 21.51 |
| High mix | 0.62/0.08 | 11.02 | 39.05 |
| Gene | Forward (5′-3′) | Reverse (5′-3′) | GenBank Accession No. |
|---|---|---|---|
| GAPDH | TGACCTGCCGTCTGGAGAAA | TGTGTATCCTAGGATGCCCTTCAG | NM_204305.1 |
| GPX4 | AGTGCCATCAAGTGGAACTTCAC | TTCAAGGCAGGCCGTCAT | NM_001346448.1 |
| GSS | GTACTCACTGGATGTGGGTGAAGA | CGGCTCGATCTTGTCCATCAG | XM_425692.6 |
| GSR | CCACCAGAAAGGGGATCTACG | ACAGAGATGGCTTCATCTTCAGTG | XM_015276627.2 |
| Gene | MGM Dual-Labeled Probe | Fluorescent Dye |
|---|---|---|
| GAPDH | CCAGCCAAGTATGATGAT | VIC |
| GPX4 | CAGCCCAATGGAG | FAM |
| GSS | AGGAGGGAACAACCTG | FAM |
| GSR | CTGGCACTTCGGCTC | FAM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulcsár, S.; Kövesi, B.; Balogh, K.; Zándoki, E.; Ancsin, Z.; Erdélyi, M.; Mézes, M. The Co-Occurrence of T-2 Toxin, Deoxynivalenol, and Fumonisin B1 Activated the Glutathione Redox System in the EU-Limiting Doses in Laying Hens. Toxins 2023, 15, 305. https://doi.org/10.3390/toxins15050305
Kulcsár S, Kövesi B, Balogh K, Zándoki E, Ancsin Z, Erdélyi M, Mézes M. The Co-Occurrence of T-2 Toxin, Deoxynivalenol, and Fumonisin B1 Activated the Glutathione Redox System in the EU-Limiting Doses in Laying Hens. Toxins. 2023; 15(5):305. https://doi.org/10.3390/toxins15050305
Chicago/Turabian StyleKulcsár, Szabina, Benjámin Kövesi, Krisztián Balogh, Erika Zándoki, Zsolt Ancsin, Márta Erdélyi, and Miklós Mézes. 2023. "The Co-Occurrence of T-2 Toxin, Deoxynivalenol, and Fumonisin B1 Activated the Glutathione Redox System in the EU-Limiting Doses in Laying Hens" Toxins 15, no. 5: 305. https://doi.org/10.3390/toxins15050305
APA StyleKulcsár, S., Kövesi, B., Balogh, K., Zándoki, E., Ancsin, Z., Erdélyi, M., & Mézes, M. (2023). The Co-Occurrence of T-2 Toxin, Deoxynivalenol, and Fumonisin B1 Activated the Glutathione Redox System in the EU-Limiting Doses in Laying Hens. Toxins, 15(5), 305. https://doi.org/10.3390/toxins15050305