Fungal Species and Multi-Mycotoxin Associated with Post-Harvest Sorghum (Sorghum bicolor (L.) Moench) Grain in Eastern Ethiopia



Abstract
1. Introduction
2. Result and Discussions
2.1. Analysis of Fungal Species in Sorghum Grain Samples
2.2. Contamination of Mycotoxin Groups in Sorghum Grain
2.3. Multi-Mycotoxins Detected in Sorghum Samples
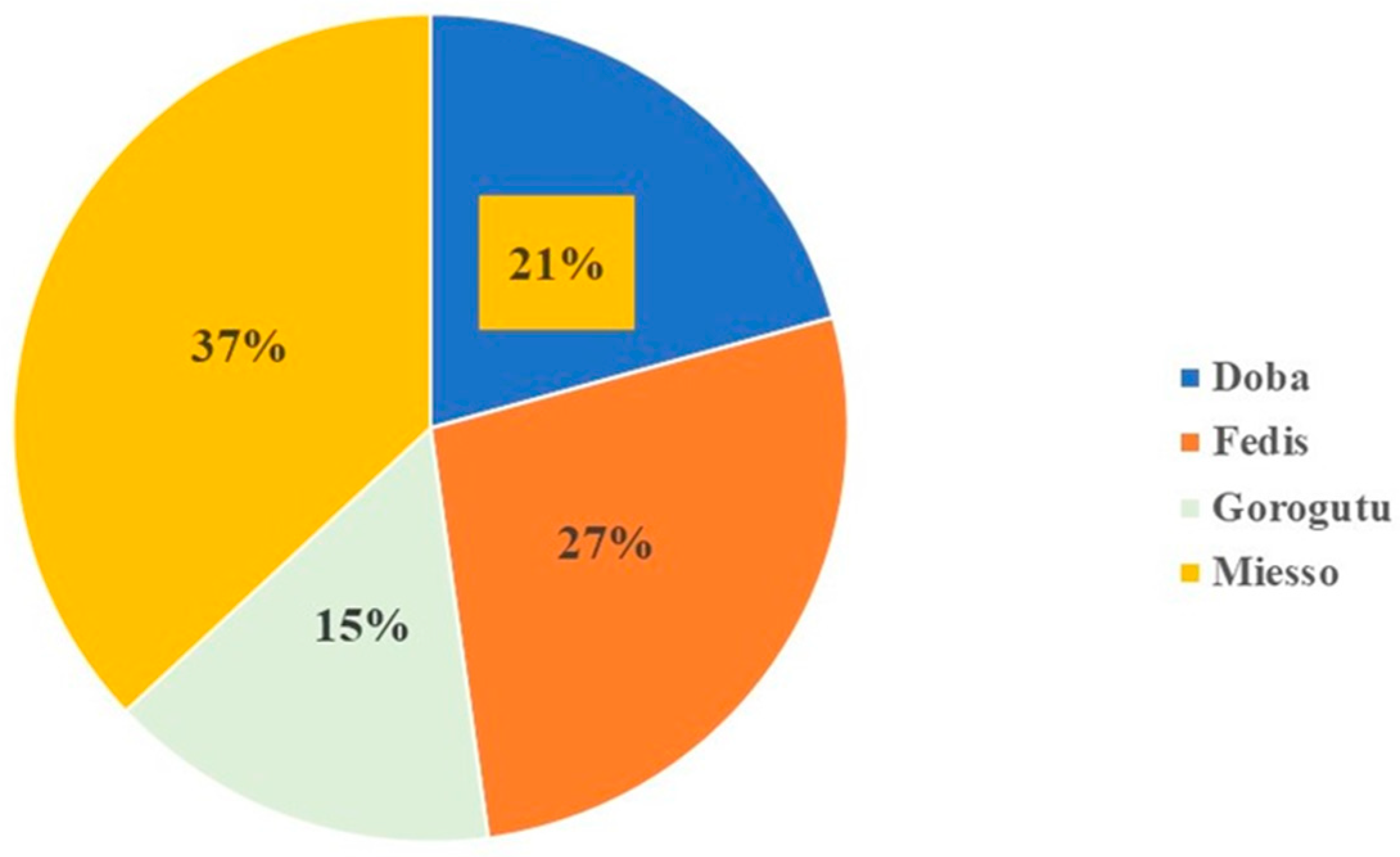
2.4. Multi-Mycotoxins in Sorghum Samples across the Targeted Districts
3. Conclusions
4. Materials and Methods
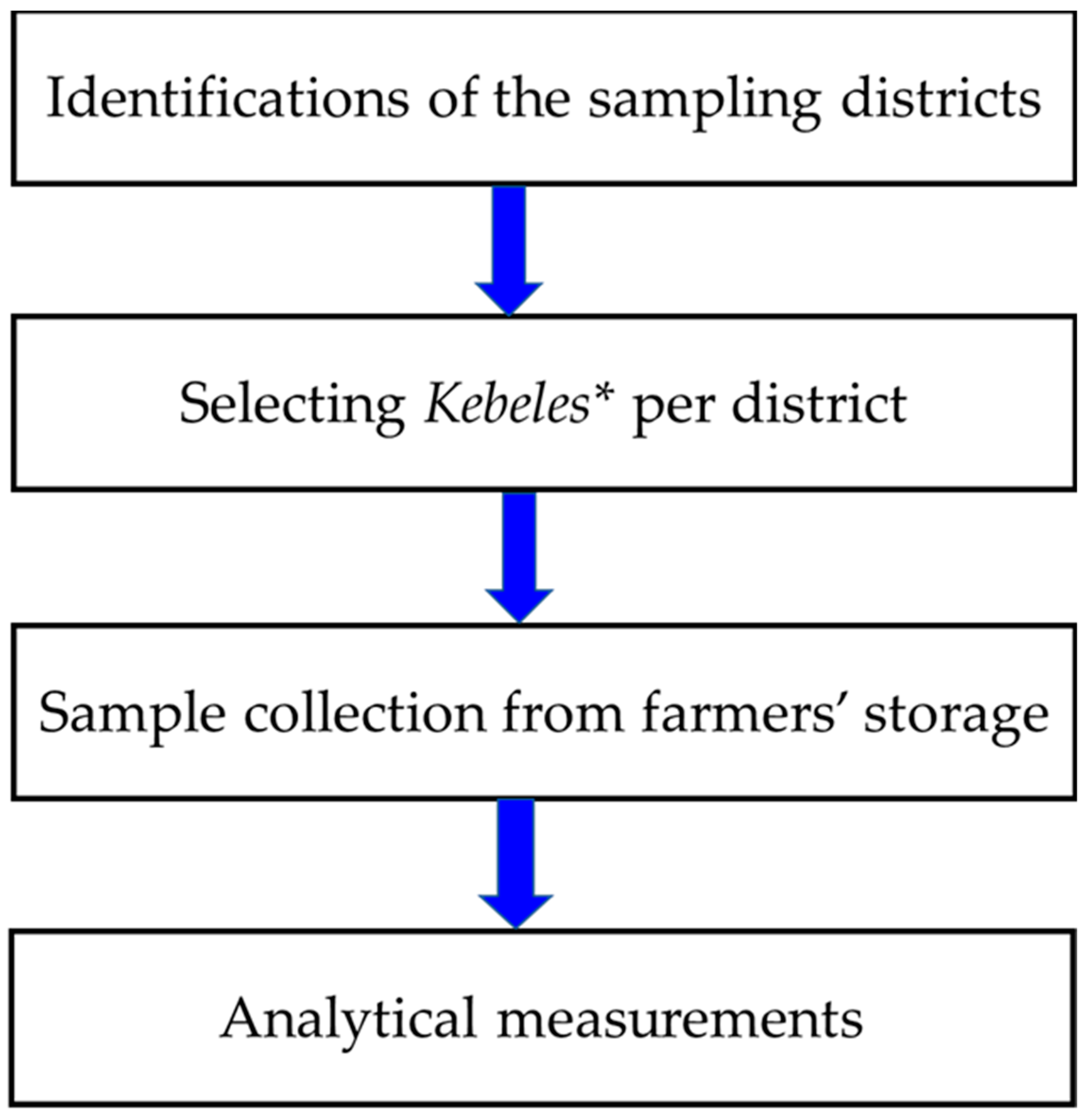
4.1. Schematic Overview of the Study
4.2. Sorghum Sample Collection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Districts | Altitude (m.a.s.l) | Min and Maximum Annual Rainfall (mm) |
|---|---|---|
| Doba | 1400–2500 | 550–800 |
| Fedis | 1200–1500 | 400–804 |
| Goro Gutu | 1250–2575 | 600–900 |
| Miesso | 1100–1400 | 400–750 |
4.3. Analysis of Fungal Species Associated with Sorghum Grain Samples
4.4. Multi-Mycotoxin Analysis in Sorghum Grain Samples
4.5. Data Evaluations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization Corporate Statistical Database. FAOSTAT Food and Agriculture Organization of the United Nations Statistical Pocketbook; World Food and Agriculture: Rome, Italy, 2018; Available online: http://www.fao.org/3/CA1796EN/ca1796en.pdf (accessed on 23 April 2022).
- Mayer, J.E.; Pfeiffer, W.H.; Beyer, P. Biofortified crops to alleviate micronutrient malnutrition. Curr. Opin. Plant Biol. 2008, 11, 166–170. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAO Food and Agriculture Data. 2021. Available online: https://www.fao.org/faostat/en/#data/QI (accessed on 24 May 2022).
- Central Statistical Agency. CSA Agricultural Sample Survey Report on Area and Production of Major Crops (Private Peasant Holdings ‘Meher’ Season); Statistical Bulletin 585; Central Statistical Agency: Addis Ababa, Ethiopia, 2020. [Google Scholar]
- Legesse, T. Physical measurements and improvement methods of protein and other nutritional quality traits of sorghum [Sorghum bicolor (L.) Moench]. Int. J. Food Eng. 2018, 2, 10–16. [Google Scholar]
- Rembold, F.; Hodges, R.; Bernard, M.; Knipschild, H.; Leo, O. The African postharvest losses information system (APHLIS). In EUR 24712 EN-Joint Research Centre-Institute for Environment and Sustainability; Publications Office of the European Union: Luxembourg, 2011. [Google Scholar]
- Tesfaye, W.; Tirivayi, N. The impacts of postharvest storage innovations on food security and welfare in Ethiopia. Food Policy 2018, 75, 52–67. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. FAO Post-harvest Loss Assessment of Maize, Wheat, Sorghum and Haricot Bean: A Study Conducted in Fourteen Elected Woredas of Ethiopia under the Project–GCP/ETH/084/SWI; Food and Agriculture Organization: Addis Ababa, Ethiopia, 2017; Available online: https://www.fao.org/fileadmin/user_upload/food-loss-reduction/Ethiopia/Ethiopia-Baseline_PHT_food_loss_assessment_report-V25jan18__003_.pdf (accessed on 25 June 2022).
- Chulze, S.N. Strategies to reduce mycotoxin levels in maize during storage: A review. Food Addit. Contam. 2010, 27, 651–657. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, J.P.F.; Placinta, C.M.; Macdonald, A.M.C. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed Sci. Technol. 1999, 80, 183–205. [Google Scholar] [CrossRef]
- Pestka, J.J.; Smolinski, A.T. Deoxynivalenol: Toxicology and potential effects on humans. J. Toxicol. Environ. Health. Part B 2005, 8, 39–69. [Google Scholar] [CrossRef]
- Ukwuru, M.U.; Ohaegbu, C.G.; Muritala, A. An overview of mycotoxin contamination of foods and feeds. J. Biochem. Microb. Toxicol. 2017, 1, 101. [Google Scholar]
- De Saeger, S. Determining Mycotoxins and Mycotoxigenic Fungi in Food and Feed; Woodhead Publishers: Cambridge, UK, 2011; ISBN 978-1-84569-674-0. [Google Scholar]
- Wild, C.P.; Turner, P.C. The toxicology of aflatoxins as basis for public health decisions. Mutagenesis 2002, 17, 471–482. [Google Scholar] [CrossRef]
- Ecker, D.J.; Sampath, R.; Willett, P.; Wyatt, J.R.; Samant, V.; Massire, C. The microbial Rosetta Stone Database: A compilation of global and emerging infectious microorganisms and bioterrorist threat agents. BMC Microbiol. 2005, 19, 19. [Google Scholar] [CrossRef]
- Ben Taheur, F.; Kouidhi, B.; Al Qurashi, Y.M.A.; Ben Salah-Abbès, J.; Chaieb, K. Review: Biotechnology of mycotoxins detoxification using microorganisms and enzymes. Toxicon 2019, 160, 12–22. [Google Scholar] [CrossRef]
- Chala, A.; Taye, W.; Ayalew, A.; Krska, R.; Sulyok, M.; Logrieco, A. Multi-mycotoxin analysis of sorghum (Sorghum bicolor L. Moench) and finger millet (Eleusine coracana L. Garten) from Ethiopia. Food Control 2014, 45, 29–35. [Google Scholar] [CrossRef]
- Weledesemayat, G.T.; Gezmu, T.B.; Woldegiorgis, A.Z.; Gemede, H.F. Study on Aspergillus species and aflatoxin levels in sorghum (Sorghum bicolor L.) stored for different period and storage system in Kewet districts, Northern Shewa, Ethiopia. J. Food Sci. Nutr. 2016, 3, 010. [Google Scholar] [CrossRef]
- Wondimeneh, T.; Amare, A.; Mashilla, D.; Alemayehu, C. Fungal invasion and mycotoxin contamination of stored sorghum grain as influenced by threshing methods. Int. J. Pest Manag. 2017, 64, 66–76. [Google Scholar] [CrossRef]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Malir, J.; Toman, J. Ochratoxin A, 50 years of research. Toxins 2016, 8, 191. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef]
- van Egmond, H.P.; Schothorst, R.C.; Jonker, M.A. Regulations relating to mycotoxins in food: Perspectives in a global and European context. Anal. Bioanal. Chem. 2007, 389, 147–157. [Google Scholar] [CrossRef]
- Jestoi, M. Emerging Fusarium mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin—A review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef]
- EFSA-European Food Safety Authority. EFSA Scientific opinion on the risks to human and animal health related to the presence of beauvericin and enniatins in food and feed: Beauvericin and enniatins in food and feed. EFSA J. 2014, 12, 3802. [Google Scholar]
- European Food Safety Authority. EFSA Scientific opinion on the risks for animal and public health related to the presence of T-2 and HT-2 toxin in food and feed. EFSA J. 2011, 9, 187. [Google Scholar] [CrossRef]
- EFSA-European Food Safety Authority. EFSA Scientific opinion on risks for animal and public health related to the presence of nivalenol in food and feed. EFSA J. 2013, 11, 3262. [Google Scholar] [CrossRef]
- EFSA-European Food Safety Authority. EFSA Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, e04718. [Google Scholar] [CrossRef]
- Motarjemi, Y. Public health measures: Modern approach to food safety management: An overview. Encycl. Food Saf. 2013, 4, 1–12. [Google Scholar] [CrossRef]
- Ssepuuya, G.; Van Poucke, C.; Ediage, E.N.; Mulholland, C.; Tritscher, A.; Verger, P.; Kenny, M.; Bessy, C.; De Saeger, S. Mycotoxin contamination of sorghum and its contribution to human dietary exposure in four sub-Saharan countries. Food Addit. Contam. Part A 2018, 35, 1384–1393. [Google Scholar] [CrossRef] [PubMed]
- Mamo, F.T.; Abate, B.A.; Tesfaye, K.; Nie, C.; Wang, G.; Liu, Y. Mycotoxins in Ethiopia: A review on prevalence, economic and health impacts. Toxins 2020, 12, 648. [Google Scholar] [CrossRef]
- Cuevas, H.E.; Prom, L.K. Evaluation of genetic diversity, agronomic traits, and anthracnose resistance in the NPGS Sudan Sorghum Core collection. BMC Genom. 2020, 21, 88. [Google Scholar] [CrossRef] [PubMed]
- Osman, M.A.; Salama, A.; Naguib, K.H.M.; Abdel-Wahhab, M.A.; Sherif, S.R. Fungi and mycotoxins associated with Egyptian sorghum grains. MOJ Toxicol. 2017, 3, 51–56. [Google Scholar] [CrossRef][Green Version]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin occurrence in feed and feed raw materials worldwide: Long-term analysis with special focus on Europe and Asia. J. Sci. Food Agric. 2013, 93, 2892–2899. [Google Scholar] [CrossRef]
- Tebele, S.M.; Gbashi, S.; Adebo, O.; Changwa, R.; Naidu, K.; Njobeh, P.B. Quantification of multi-mycotoxin in cereals (maize, maize porridge, sorghum and wheat) from Limpopo province of South Africa. Food Addit. Contam. Part A 2020, 37, 1922–1938. [Google Scholar] [CrossRef]
- European Commission. EU Regulation (1126/2007) amending Regulation (EC) No 1881/2006 setting maximum levels for certain contaminants in foodstuffs as regards Fusarium toxins in maize and maize products. Off. J. Eur. Union. 2007, 255, 14–17. [Google Scholar]
- Hanvi, D.M.; Lawson-Evi, P.; De Boevre, M.; Goto, C.E.; De Saeger, S.; Eklu-Gadegbeku, K. Natural occurrence of mycotoxins in maize and sorghum in Togo. Mycotoxin Res. 2019, 35, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Ayalew, A.; Fehrmann, H.; Lepschy, J.; Beck, R.; Abate, D. Natural occurrence of mycotoxins in staple cereals from Ethiopia. Mycopathologia 2006, 162, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Wageh, S.D.; Yoshinori, I.; Shouta, M.M.N.; Mayumi, I. An overview on mycotoxin contamination of foods in Africa. J. Vet. Med. Sci. 2014, 76, 789–797. [Google Scholar] [CrossRef]
- Bandyopadhyay, R.; Kumar, M.; Leslie, J.F. Relative severity of aflatoxin contamination of cereal crops in West Africa. Food Addit. Contam. 2007, 24, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Ediage, E.N.; Van Poucke, C.; De Saeger, S. A multi-analyte LC–MS/ MS method for the analysis of 23 mycotoxins in different sorghum varieties: The forgotten sample matrix. Food Chem. 2015, 177, 397–404. [Google Scholar] [CrossRef]
- Ghali, R.; Belouaer, I.; Hdiri, S.; Ghorbel, H.; Maaroufi, K.; Hedilli, A. Simultaneous HPLC determination of aflatoxins B1, B2, G1 and G2 in Tunisian sorghum and pistachios. J. Food Compos. Anal. 2009, 22, 751–755. [Google Scholar] [CrossRef]
- Barber, M. The UK Code of Good Storage Practice to Reduce Ochratoxin A in Cereals. Food Standards Agency. 2007. Available online: https://www.food.gov.uk/sites/default/files/multimedia/pdfs/ochratoxinacop.pdf (accessed on 23 June 2022).
- Ekwomadu, T.I.; Akinola, S.A.; Mwanza, M. Fusarium Mycotoxins, their metabolites (Free, Emerging, and Masked), Food Safety Concerns, and Health Impacts. Int. J. Environ. Health Res. 2021, 18, 11741. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Novak, B.; Nagl, V.; Berthiller, F. Emerging mycotoxins: Beyond traditionally determined food contaminants. J. Agric. Food Chem. 2017, 65, 7052–7070. [Google Scholar] [CrossRef]
- Santini, A.; Meca, G.; Uhlig, S.; Ritieni, A. Fusaproliferin, beauvericin and enniatins: Occurrence in food—review. World Mycotoxin J. 2012, 5, 71–81. [Google Scholar] [CrossRef]
- Agriopoulou, S. Enniatins: An Emerging Food Safety Issue. EC Nutr. 2016, 3, 1142–1146. [Google Scholar]
- Sanhueza, C.E.P.; Degrossi, M.C. Moniliformin, a Fusarium mycotoxin. Rev. Mex. Micol. 2004, 19, 103–112. [Google Scholar]
- Thiel, P.G.; Meyer, C.J.; Marasas, W.F.O. Natural occurrence of moniliformin together with deoxynivalenol and zearalenone in Transkei corn. J. Agric. Food Chem. 1982, 30, 308–317. [Google Scholar] [CrossRef]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef]
- Nafuka, S.N.; Misihairabgwi, J.M.; Bock, R.; Ishola, A.; Sulyok, M.; Krska, R. Variation of fungal metabolites in sorghum malts used to prepare Namibian traditional fermented beverages Omalodu and Otombo. Toxins 2019, 11, 165. [Google Scholar] [CrossRef] [PubMed]
- Escrivá, L.; Oueslati, S.; Font, G.; Manyes, L. Alternaria mycotoxins in food and feed: An Overview. J. Food Qual. 2017, 2017, 1569748. [Google Scholar] [CrossRef]
- Ezekiel, C.N.; Bandyopadhyay, R.; Sulyok, M.; Warth, B.; Krska, R. Fungal and bacterial metabolites in commercial poultry feed from Nigeria. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2012, 29, 1288–1299. [Google Scholar] [CrossRef] [PubMed]
- Ayalew, A. Mycotoxins and surface and internal fungi of maize from Ethiopia. Afr. J. Food Agric. Nutr. Dev. 2010, 10, 4109–4123. [Google Scholar] [CrossRef]
- Tsehaye, H.; Brurberg, M.B.; Sundheim, L.; Assefa, D.; Tronsmo, A.; Tronsmo, A.M. Natural occurrence of Fusarium species and fumonisin on maize grains in Ethiopia. Eur. J. Plant Pathol. 2017, 147, 141–155. [Google Scholar] [CrossRef]
- Yilma, S.; Sadessa, K.; Kebede, D. Fungal Infections and Aflatoxin Contamination in Maize Grains Collected from West Showa and East Wallega. Int. J. Curr. Res. Rev. 2019, 11, 16–22. [Google Scholar] [CrossRef]
- Mesfin, A.; Tesfamariam, K.; Belachew, T.; De Saeger, S.; Lachat, C.; De Boevre, M. Multi-mycotoxin profiling in maize reveals prevalence of Fusarium mycotoxins in South and West Ethiopia. World Mycotoxin J. 2021, 15, 73–83. [Google Scholar] [CrossRef]
- Fern´andez-Cruz, M.; Mansilla, L.; Tadeo, J.L. Mycotoxins in fruits and their processed products: Analysis, occurrence and health implications. J. Adv. Res. 2010, 2, 113–122. [Google Scholar] [CrossRef]
- Ostry, V. Alternaria mycotoxins: An overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs. World Mycotoxin J. 2008, 1, 175–188. [Google Scholar] [CrossRef]
- Scott, P.M.; Zhao, W.; Feng, S.; Lau, B.P.Y. Alternaria toxins alternariol and alternariol monomethyl ether in grain foods in Canada. Mycotoxin Res. 2012, 28, 261–266. [Google Scholar] [CrossRef]
- Muluken, G.; Yoseph, A.; Augustine, N.; Chemeda, F. Cereal stem borer management practices in subsistence farms of eastern Ethiopia. Int. J. Pest Manag. 2017, 63, 289–298. [Google Scholar]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi. Laboratory Manual Series; Fungal Biodiversity Center: Utrecht, The Netherlands, 2010. [Google Scholar]
- Sulyok, M.; Stadler, D.; Steiner, D.; Krska, R. Validation of an LC-MS/MS-based dilute-and-shoot approach for the quantification of > 500 mycotoxins and other secondary metabolites in food crops: Challenges and solutions. Anal. Bioanal. Chem. 2020, 412, 2607–2620. [Google Scholar] [CrossRef]
- Laatsch, H. AntiBase, a Data Base for Rapid Dereplication and Structure Determination of Microbial Natural Products; Wiley-VCH: Weinheim, Germany, 2009; Available online: http://www.user.gwdg.de/nucoc/laatsch/AnitBase.htm (accessed on 24 June 2022).
- Frisvad, J.C.; Filtenborg, O.; Thrane, U. Analysis and screening for mycotoxins and other metabolites in fungal cultures by thin-layer chromatography and high-performance liquid chromatography. Arch. Environ. Contam. Toxicol. 1989, 18, 331–335. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Andersen, B.; Thrane, U. The use of secondary metabolite profiling in chemotaxonomy of filamentous fungi. Mycol. Res. 2007, 112, 231–240. [Google Scholar] [CrossRef]




| Mycotoxins Group | Doba | Fedis | Goro Gutu | Miesso |
|---|---|---|---|---|
| Major mycotoxins and derivatives | 40.0 | 15.0 | 40.0 | 65.0 |
| Fusarium metabolites | 95.0 | 100 | 100 | 100 |
| Aspergillus metabolites | 100 | 60.0 | 95.0 | 90.0 |
| Penicillium metabolites | 100 | 100 | 100 | 100 |
| Alternaria metabolites | 90.0 | 100 | 95.0 | 100 |
| Metabolites from other fungal genera | 100 | 100 | 100 | 100 |
| Bacterial metabolites | 60.0 | 55.0 | 80.0 | 95.0 |
| Unspecified metabolites | 100 | 100 | 100 | 100 |
| Mycotoxins Group | Specific Metabolites | % Positive Samples | Concentration (μg/kg) | ||
|---|---|---|---|---|---|
| Mean | Median | Max | |||
| Major mycotoxins and derivatives | Aflatoxin B1 | 1 (1.25) | 0.29 | <LOD | 23.6 |
| Aflatoxin B2 | 1 (1.25) | 0.01 | <LOD | 0.79 | |
| Aflatoxin G1 | 1 (1.25) | 0.01 | <LOD | 0.62 | |
| Ochratoxin A | 4 (5.0) | 2.89 | <LOD | 115 | |
| Ochratoxin B | 4 (5.0) | 0.28 | <LOD | 13.8 | |
| Fumonisin B1 | 5 (6.25) | 2.78 | <LOD | 112 | |
| Fumonisin B2 | 2 (2.5) | 0.52 | <LOD | 30.3 | |
| Deoxynivalenol | 5 (6.25) | 2.70 | <LOD | 137 | |
| Nivalenol | 13 (16) | 1.66 | <LOD | 22.3 | |
| Monoacetoxyscirpenol | 8 (10) | 2.38 | <LOD | 59.1 | |
| Diacetoxyscirpenol | 13 (16) | 0.88 | <LOD | 15.3 | |
| Zearalenone | 19 (24) | 3.62 | <LOD | 121 | |
| alpha-Zearalenol | 1 (1.25) | 0.08 | <LOD | 6.52 | |
| Fusarium metabolites | Moniliformin | 74 (93) | 58.0 | 25.62 | 437 |
| Beauvericin | 17 (21) | 0.57 | <LOD | 10.2 | |
| Enniatin A1 | 2 (2.50) | 0.02 | <LOD | 0.95 | |
| Enniatin B | 8 (10) | 0.02 | <LOD | 0.65 | |
| Enniatin B1 | 3 (3.75) | 0.03 | <LOD | 1.14 | |
| Antibiotic Y | 3 (3.75) | 0.15 | <LOD | 8.29 | |
| Aurofusarin | 15 (19) | 4.79 | <LOD | 116 | |
| Bikaverin | 67 (84) | 80.9 | 42.3 | 468 | |
| Chrysogin | 20 (25) | 1.72 | <LOD | 34 | |
| Deoxyfusapyron | 39 (49) | 14.8 | <LOD | 306 | |
| Epiequisetin | 4 (5.00) | 0.02 | <LOD | 0.61 | |
| Equisetin | 23 (29) | 2.42 | <LOD | 32.9 | |
| Fusapyron | 7 (8.80) | 0.99 | <LOD | 27.3 | |
| Fusaric acid | 43 (54) | 204 | 36.6 | 2790 | |
| Gibepyron D | 2 (2.50) | 1.18 | <LOD | 50.9 | |
| Sambutoxin | 6 (7.50) | 0.02 | <LOD | 0.71 | |
| Siccanol | 70 (88) | 240 | 110 | 2374 | |
| W493 | 6 (7.50) | 1.25 | <LOD | 31.9 | |
| Aspergillus metabolites | Sterigmatocystin | 24 (30) | 2.80 | <LOD | 50.5 |
| Methoxysterigmatocystin | 5 (6.25) | 0.80 | <LOD | 29.7 | |
| Versicolorin C | 8 (10) | 0.19 | <LOD | 6.08 | |
| Averufin | 23 (29) | 0.66 | <LOD | 11.7 | |
| 8-O-Methylaverufin | 12 (15) | 0.32 | <LOD | 8.54 | |
| Kojic acid | 8 (10) | 197 | <LOD | 4580 | |
| 3-Nitropropionic acid | 49 (61) | 67.8 | 9.07 | 817 | |
| Asperflavine | 27 (34) | 3.83 | <LOD | 63 | |
| Asperfuran | 28 (35) | 218 | <LOD | 2630 | |
| Asperloxine A | 1 (1.25) | 0.04 | <LOD | 3.05 | |
| Aspochracin | 8 (10) | 6.42 | <LOD | 226 | |
| Sydonic acid | 2 (2.50) | 0.04 | <LOD | 2.77 | |
| Viomellein | 2 (2.50) | 21.2 | <LOD | 1410 | |
| Penicillium metabolites | Mycophenolic acid | 48 (60) | 242 | 8.40 | 8300 |
| Mycophenolic acid IV | 16 (20) | 1.13 | <LOD | 47 | |
| 1-Deoxypebrolide | 38 (48) | 5.46 | <LOD | 127 | |
| 7-Hydroxypestalotin | 6 (7.50) | 0.44 | <LOD | 13.4 | |
| Barceloneic acid | 13 (16) | 1.27 | <LOD | 32.8 | |
| Chanoclavin | 4 (5.00) | 0.06 | <LOD | 1.71 | |
| Cycloaspeptide A | 5 (6.25) | 1.28 | <LOD | 90.5 | |
| Cyclopenin | 4 (5.00) | 0.08 | <LOD | 2.47 | |
| Cyclopenol | 5 (6.25) | 0.93 | <LOD | 33.5 | |
| Dechlorogriseofulvin | 6 (7.50) | 0.52 | <LOD | 23.5 | |
| Dehydrogriseofulvin | 2 (2.50) | 0.01 | <LOD | 0.47 | |
| F01 1358-A | 1 (1.25) | 0.10 | <LOD | 8.38 | |
| Flavoglaucin | 80 (100) | 128 | 12.8 | 1870 | |
| Griseofulvin | 15 (19) | 0.96 | <LOD | 25.3 | |
| NP1793 | 5 (6.25) | 3.23 | <LOD | 129 | |
| O-Methylviridicatin | 4 (5.00) | 0.08 | <LOD | 5.29 | |
| Penicillic acid | 3 (3.75) | 0.66 | <LOD | 27.9 | |
| Quinolactacin A | 4 (5.00) | 0.07 | <LOD | 3.78 | |
| Quinolactacin B | 1 (1.25) | 0.01 | <LOD | 0.41 | |
| PF 1163A | 2 (2.50) | 0.02 | <LOD | 1.33 | |
| Rugulovasine A | 5 (6.25) | 0.58 | <LOD | 24.8 | |
| Alternaria metabolites | Tenuazonic acid | 74 (93) | 155 | 70.5 | 1520 |
| Alternariol | 47 (59) | 3.39 | 0.73 | 46.4 | |
| Alternariolmethylether | 40 (50) | 2.38 | 0.26 | 35.2 | |
| Tentoxin | 46 (58 | 1.07 | 0.45 | 9.33 | |
| Macrosporin | 57 (71) | 5.61 | 2.32 | 77.1 | |
| Radicinin | 13 (16) | 6.19 | <LOD | 126 | |
| Metabolites from other fungal genera | Abscisic acid | 80 (100) | 17.5 | 15.8 | 46.5 |
| Cytochalasin B | 2 (2.50) | 1.45 | <LOD | 109 | |
| Destruxin A | 3 (3.75) | 0.06 | <LOD | 2.37 | |
| Monocerin | 50 (63) | 6.05 | 0.69 | 119 | |
| Preussin | 5 (6.25) | 0.61 | <LOD | 28.3 | |
| Terphenyllin | 26 (33) | 61.6 | <LOD | 1980 | |
| Terrein | 1 (1.25) | 1.49 | <LOD | 120 | |
| Trichodermamide C | 4 (5.00) | 0.40 | <LOD | 11.1 | |
| Bacterial metabolites | Chloramphenicol | 55 (69) | 0.39 | 0.24 | 4.98 |
| Monactin | 2 (2.50) | 0.02 | <LOD | 1.06 | |
| Nonactin | 4 (5.00) | 0.12 | <LOD | 5.4 | |
| Puromycin | 3 (3.75) | 0.35 | <LOD | 14.1 | |
| Unspecific metabolites | Asperglaucide | 32 (40) | 76.52 | <LOD | 1583 |
| Brevianamid F | 26 (33) | 2.86 | <LOD | 110 | |
| cyclo(L-Pro-L-Tyr) | 80 (100) | 33.58 | 20.90 | 574 | |
| cyclo(L-Pro-L-Val) | 8 (10) | 5.71 | <LOD | 410 | |
| Emodin | 30 (38) | 1.13 | <LOD | 24.6 | |
| Fallacinol | 22 (28) | 3.32 | <LOD | 46 | |
| N-Benzoyl-Phenylalanine | 25 (31) | 2.56 | <LOD | 51 | |
| Neoechinulin A | 44 (55) | 172 | 7.85 | 2000 | |
| Neoechinulin D | 54 (68) | 27.5 | 0.94 | 329 | |
| Physcion | 18 (23) | 21.2 | <LOD | 309 | |
| Tryptophol | 11 (14) | 5.08 | <LOD | 210 | |
| Mycotoxin Groups | Specific Metabolites | Doba | Fedis | Goro Gutu | Miesso |
|---|---|---|---|---|---|
| Means ± SD | Means ± SD | Means ± SD | Means ± SD | ||
| Major mycotoxins and derivatives | Aflatoxin B1 | <LOD | <LOD | 1.18 ± 5.28 | <LOD |
| Aflatoxin B2 | <LOD | <LOD | 0.04 ± 0.18 | <LOD | |
| Aflatoxin G1 | <LOD | <LOD | 0.03 ± 0.14 | <LOD | |
| Ochratoxin A | 7.86 ± 26.2 | <LOD | 3.71 ± 16.57 | <LOD | |
| Ochratoxin B | 0.43 ± 1.10 | <LOD | 0.69 ± 3.08 | <LOD | |
| Fumonisin B1 | 7.00 ± 25.45 | <LOD | <LOD | 4.12 ± 9.50 | |
| Fumonisin B2 | 1.52 ± 6.78 | <LOD | <LOD | 0.58 ± 2.59 | |
| Deoxynivalenol | 8.45 ± 30.74 | <LOD | <LOD | 2.36 ± 7.96 | |
| Nivalenol | <LOD | 0.51 ± 2.28 | 1.08 ± 2.77 | 5.05 ± 7.13 | |
| Monoacetoxyscirpenol | 1.08 ± 4.83 | 2.96 ± 13.22 | <LOD | 5.49 ± 10.1 | |
| Diacetoxyscirpenol | 0.70 ± 3.13 | 0.76 ± 3.42 | 0.24 ± 0.92 | 1.84 ± 3.47 | |
| Zearalenone | 2.15 ± 7.65 | 0.08 ± 0.38 | 9.47 ± 27.97 | 2.77 ± 5.98 | |
| alpha-Zearalenol | <LOD | <LOD | 0.33 ± 1.46 | <LOD | |
| Fusarium metabolites | Moniliformin | 26.5 ± 38.1 | 27.1 ± 22.3 | 29.5 ± 27.9 | 149 ±99.1 |
| Beauvericin | 0.14 ± 047 | 0.06 ± 0.15 | 0.56 ± 2.19 | 1.53 ± 2.95 | |
| Enniatin A1 | <LOD | <LOD | 0.09 ± 0.28 | <LOD | |
| Enniatin B | 0.02 ± 0.01 | 0.01 ± 0.02 | 0.07 ± 0.17 | 0.01 ± 0.03 | |
| Enniatin B1 | <LOD | 0.02 ± 0.07 | 0.09 ± 0.31 | <LOD | |
| Antibiotic Y | <LOD | 0.15 ± 0.65 | <LOD | 0.47 ± 1.86 | |
| Aurofusarin | 1.37 ± 5.54 | 2.76 ± 12.36 | 5.13 ± 16.8 | 9.92 ± 26.14 | |
| Bikaverin | 23.1 ± 51.1 | 68.3 ± 41.8 | 50.2 ± 98.6 | 182 ± 111 | |
| Chrysogin | 2.53 ± 7.73 | <LOD | 2.48 ± 3.84 | 1.85 ± 2.62 | |
| Deoxyfusapyron | 3.53 ± 13.9 | 10.5 ± 20.5 | 4.66 ± 11.5 | 40.4 ± 66.1 | |
| Epiequisetin | 0.02 ± 0.08 | 0.02 ± 0.07 | <LOD | 0.05 ± 0.15 | |
| Equisetin | 0.97 ± 3.11 | 0.97 ± 2.26 | 2.92 ± 6.63 | 4.79 ± 8.53 | |
| Fusapyron | <LOD | <LOD | 0.50 ± 2.26 | 3.49 ± 6.83 | |
| Fusaric acid | 41.8 ± 115 | 65.4 ± 80.8 | 82.4 ± 184 | 628 ± 649 | |
| Gibepyron D | <LOD | <LOD | <LOD | 4.73 ± 14.6 | |
| Sambutoxin | 0.01 ± 0.02 | <LOD | <LOD | 0.06 ± 0.17 | |
| Siccanol | 46.5 ± 47.9 | 274 ± 502 | 135 ± 243 | 506 ± 544 | |
| W493 | 0.76 ± 3.39 | 1.55 ± 6.93 | 2.53 ± 7.49 | 0.15 ± 0.67 | |
| Aspergillus metabolites | Sterigmatocystin | 2.86 ± 11.27 | 2.18 ± 4.74 | 1.66 ± 4.88 | 4.49 ± 11.0 |
| Methoxysterigmatocystin | 1.78 ± 6.70 | 0.06 ± 0.27 | 1.38 ± 4.33 | <LOD | |
| Versicolorin C | 0.34 ± 1.36 | 0.11 ± 0.37 | 0.29 ± 0.72 | 0.04 ± 0.19 | |
| Averufin | 0.73 ± 2.64 | 0.48 ± 0.82 | 0.82 ± 2.29 | 0.59 ± 1.44 | |
| 8-O-Methylaverufin | 0.13 ± 0.52 | 0.07 ± 0.22 | 0.37 ± 1.51 | 0.73 ± 1.98 | |
| Kojic acid | 235 ± 1024 | <LOD | 546 ± 1301 | 9.31 ± 34.0 | |
| 3-Nitropropionic acid | 85.0 ± 115 | 9.47 ± 40.36 | 84.2 ± 124 | 92.7 ± 199 | |
| Asperflavine | 4.41 ± 10.23 | 9.55 ± 18.35 | 1.01 ± 1.66 | 0.34 ± 1.00 | |
| Asperfuran | 307 ± 639 | 92.2 ±149 | 235 ± 419 | 238 ± 466 | |
| Asperloxine A | 0.15 ± 0.68 | <LOD | <LOD | <LOD | |
| Aspochracin | 1.77 ± 5.54 | 11.3 ± 50.6 | 4.12 ± 15.1 | 8.49 ± 36.2 | |
| Sydonic acid | <LOD | 0.14 ± 0.62 | 0.03 ± 0.11 | <LOD | |
| Viomellein | 70.6 ± 316 | <LOD | 14.1 ± 63.1 | <LOD | |
| Penicillium metabolites | Mycophenolic acid | 167 ± 273 | 33.8 ±138 | 763 ±1801 | 3.07 ± 5.57 |
| Mycophenolic acid IV | 0.70 ± 1.37 | 0.32 ± 1.45 | 3.51 ± 10.4 | <LOD | |
| 1-Deoxypebrolide | 8.50 ± 8.53 | 0.12 ± 0.53 | 13.21 ± 27.6 | <LOD | |
| 7-Hydroxypestalotin | <LOD | <LOD | 1.75 ± 3.34 | <LOD | |
| Barceloneic acid | <LOD | 0.75 ± 2.37 | <LOD | 4.31 ± 7.34 | |
| Chanoclavin | 0.10 ± 0.31 | <LOD | 0.05 ± 0.24 | 0.08 ± 0.38 | |
| Cycloaspeptide A | <LOD | 4.55 ± 20.2 | <LOD | 0.60 ± 2.23 | |
| Cyclopenin | 0.03 ± 0.16 | 0.12 ± 0.55 | 0.16 ± 0.55 | <LOD | |
| Cyclopenol | 1.04 ± 4.67 | 0.22 ± 0.97 | 2.48 ± 7.73 | <LOD | |
| Dechlorogriseofulvin | 1.57 ± 5.33 | 0.27 ± 1.19 | 0.11 ± 0.48 | 0.14 ± 0.63 | |
| Dehydrogriseofulvin | 0.01 ± 0.03 | <LOD | 0.02 ± 0.11 | <LOD | |
| F01 1358-A | <LOD | <LOD | 0.42 ± 1.87 | <LOD | |
| Flavoglaucin | 267 ± 471 | 118 ± 181 | 91.3 ± 148 | 34.5 ± 64.5 | |
| Griseofulvin | 2.51 ± 6.68 | 0.52 ± 1.00 | 0.57 ± 1.13 | 0.23 ± 1.02 | |
| NP1793 | 0.92 ± 2.86 | 6.45 ± 28.9 | 1.85 ± 8.29 | 3.71 ± 16.6 | |
| O-Methylviridicatin | <LOD | 0.33 ± 1.19 | 0.01 ± 0.04 | <LOD | |
| Penicillic acid | 1.56 ± 6.24 | <LOD | 1.09 ± 4.90 | <LOD | |
| Quinolactacin A | 0.21 ± 0.84 | <LOD | 0.05 ± 0.18 | <LOD | |
| Quinolactacin B | 0.02 ± 0.09 | <LOD | <LOD | <LOD | |
| PF 1163A | <LOD | <LOD | 0.09 ± 0.03 | <LOD | |
| Rugulovasine A | 1.41 ± 5.55 | 0.76 ± 2.35 | 0.17 ± 0.77 | <LOD | |
| Alternaria metabolites | Tenuazonic acid | 47.9 ± 43.5 | 180 ± 195 | 79.4 ± 96.1 | 312 ± 321 |
| Alternariol | 0.58 ± 1.09 | 3.45 ± 7.22 | 3.55 ± 10.32 | 5.99 ± 10.1 | |
| Alternariolmethylether | 0.39 ± 1.04 | 2.64 ± 7.76 | 2.72 ± 7.30 | 3.78 ± 6.82 | |
| Tentoxin | 0.17 ± 0.30 | 1.46 ± 1.57 | 0.22 ± 0.64 | 2.45 ± 2.51 | |
| Macrosporin | 0.88 ± 1.89 | 11.0 ± 17.1 | 0.75 ± 0.94 | 9.78 ± 5.73 | |
| Radicinin | 0.46 ± 2.05 | 12.2 ± 29.4 | 9.34 ± 23.13 | 2.82 ± 10.23 | |
| Metabolites from other fungal genera | Abscisic acid | 16.6 ± 6.66 | 19.4 ± 8.42 | 14.5 ± 4.58 | 19.49 ± 4.56 |
| Cytochalasin B | <LOD | 0.36 ± 1.61 | 5.43 ± 24.3 | <LOD | |
| Destruxin A | 0.19 ± 0.61 | 0.08 ± 0.35 | <LOD | <LOD | |
| Monocerin | 8.68 ± 27.2 | 7.16 ± 7.92 | 4.68 ± 9.50 | 3.68 ± 7.17 | |
| Preussin | 0.63 ± 1.95 | 1.42 ± 6.33 | 0.25 ± 1.13 | 0.16 ± 0.69 | |
| Terphenyllin | 13.2 ± 25.4 | 136 ± 438 | 10.9 ±21.9 | 86.3 ± 210 | |
| Terrein | 5.99 ± 26.8 | <LOD | <LOD | <LOD | |
| Trichodermamide C | 0.56 ± 1.75 | 0.55 ± 2.47 | 0.49 ± 2.23 | <LOD | |
| Bacterial metabolites | Chloramphenicol | 0.15 ± 0.19 | 0.12 ± 0.16 | 0.32 ± 0.31 | 1.00 ± 1.09 |
| Monactin | <LOD | 0.03 ± 0.14 | <LOD | 0.05 ± 0.23 | |
| Nonactin | <LOD | 0.27 ± 1.21 | <LOD | 0.20 ± 0.76 | |
| Puromycin | 0.32 ± 1.41 | 0.70 ± 3.14 | <LOD | 0.39 ± 1.75 | |
| Unspecific metabolites | Asperglaucide | 58.0 ± 135 | 63.1 ±130 | 1.74 ± 3.86 | 183 ± 420 |
| Brevianamid F | 1.01 ± 2.14 | 6.22 ± 24.4 | 1.51 ± 2.18 | 2.71 ± 2.79 | |
| cyclo(L-Pro-L-Tyr) | 19.9 ± 12.4 | 48.9 ± 124 | 22.4 ± 12.3 | 43.1 ±27.8 | |
| cyclo(L-Pro-L-Val) | <LOD | 20.5 ± 91.6 | <LOD | 2.34 ± 3.31 | |
| Emodin | 0.62 ± 1.33 | 2.99 ± 5.81 | 0.67 ± 1.39 | 0.26 ± 0.46 | |
| Fallacinol | 3.18 ± 7.23 | 6.98 ± 12.6 | 2.64 ± 8.62 | 0.51 ± 1.66 | |
| N-Benzoyl-Phenylalanine | 3.39 ± 5.64 | 0.32 ± 0.60 | 4.70 ± 11.8 | 1.84 ± 4.08 | |
| Neoechinulin A | 250 ± 480 | 267 ± 451 | 109 ± 176 | 59.9 ± 136 | |
| Neoechinulin D | 33.4 ± 76.2 | 49.6 ± 93.4 | 16.9 ± 27.2 | 9.99 ± 21.2 | |
| Physcion | 28.7 ± 69.4 | 47.0 ± 78.0 | 6.37 ± 19.6 | 2.85 ± 12.8 | |
| Tryptophol | 5.15 ± 10.2 | 11.5 ± 47.0 | 3.73 ± 7.78 | <LOD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammed, A.; Bekeko, Z.; Yusufe, M.; Sulyok, M.; Krska, R. Fungal Species and Multi-Mycotoxin Associated with Post-Harvest Sorghum (Sorghum bicolor (L.) Moench) Grain in Eastern Ethiopia. Toxins 2022, 14, 473. https://doi.org/10.3390/toxins14070473
Mohammed A, Bekeko Z, Yusufe M, Sulyok M, Krska R. Fungal Species and Multi-Mycotoxin Associated with Post-Harvest Sorghum (Sorghum bicolor (L.) Moench) Grain in Eastern Ethiopia. Toxins. 2022; 14(7):473. https://doi.org/10.3390/toxins14070473
Chicago/Turabian StyleMohammed, Abdi, Zelalem Bekeko, Mawardi Yusufe, Michael Sulyok, and Rudolf Krska. 2022. "Fungal Species and Multi-Mycotoxin Associated with Post-Harvest Sorghum (Sorghum bicolor (L.) Moench) Grain in Eastern Ethiopia" Toxins 14, no. 7: 473. https://doi.org/10.3390/toxins14070473
APA StyleMohammed, A., Bekeko, Z., Yusufe, M., Sulyok, M., & Krska, R. (2022). Fungal Species and Multi-Mycotoxin Associated with Post-Harvest Sorghum (Sorghum bicolor (L.) Moench) Grain in Eastern Ethiopia. Toxins, 14(7), 473. https://doi.org/10.3390/toxins14070473