Phenotypic and Transcriptomics Analyses Reveal Underlying Mechanisms in a Mouse Model of Corneal Bee Sting

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
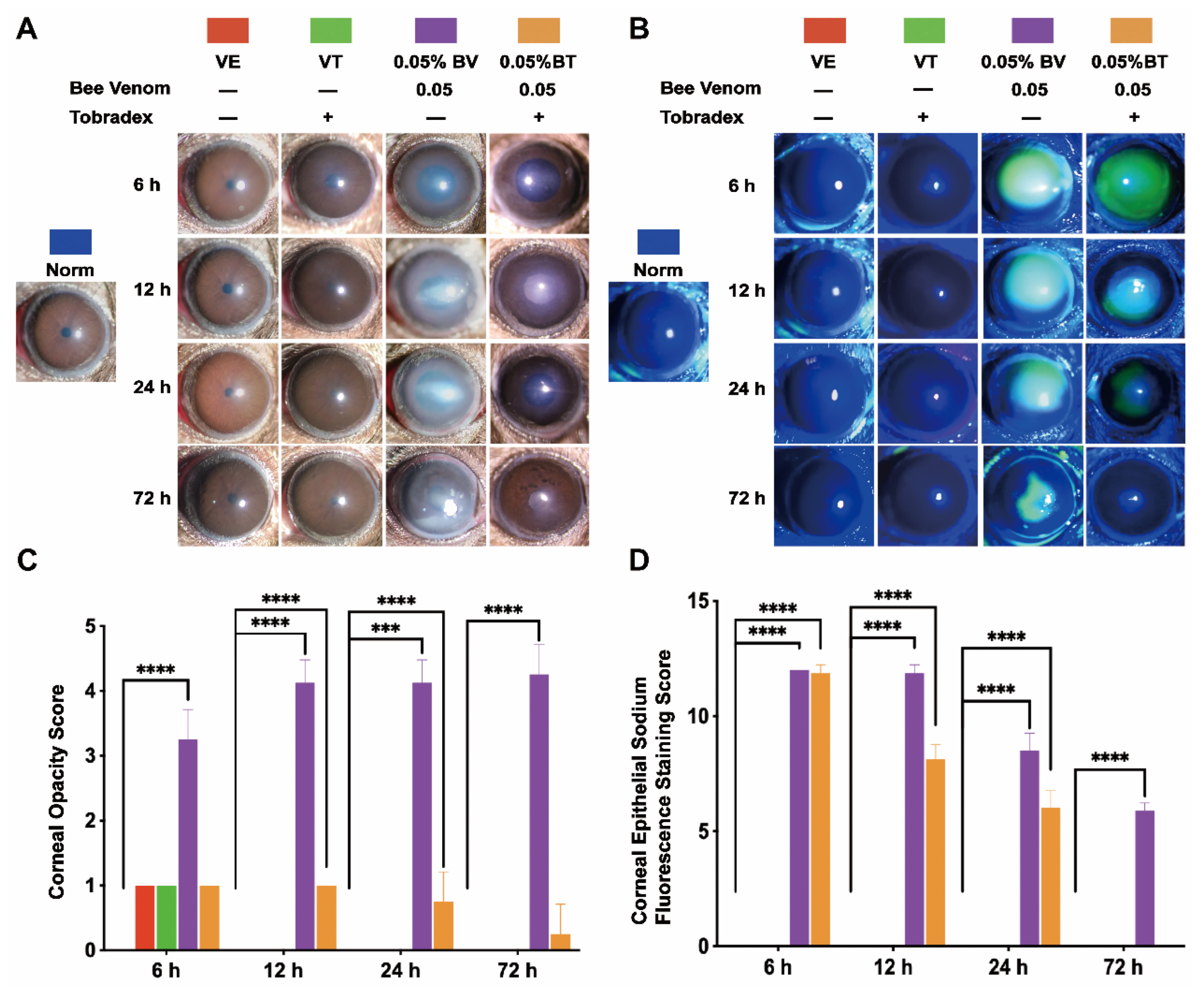
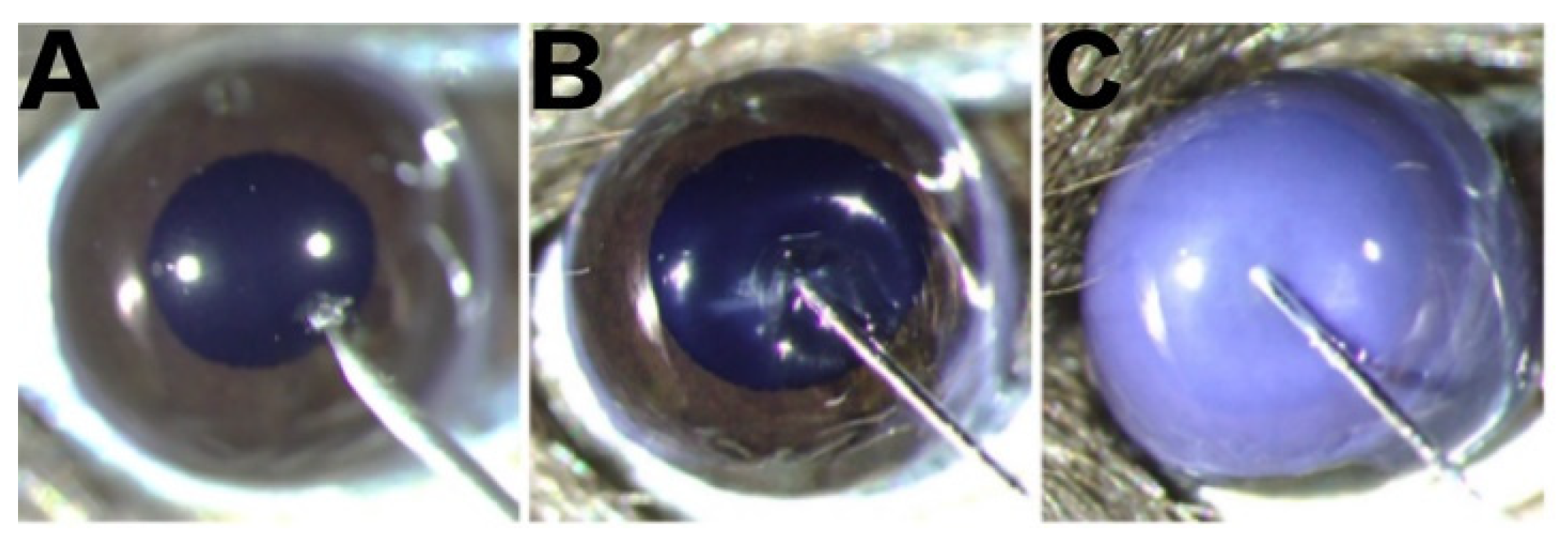
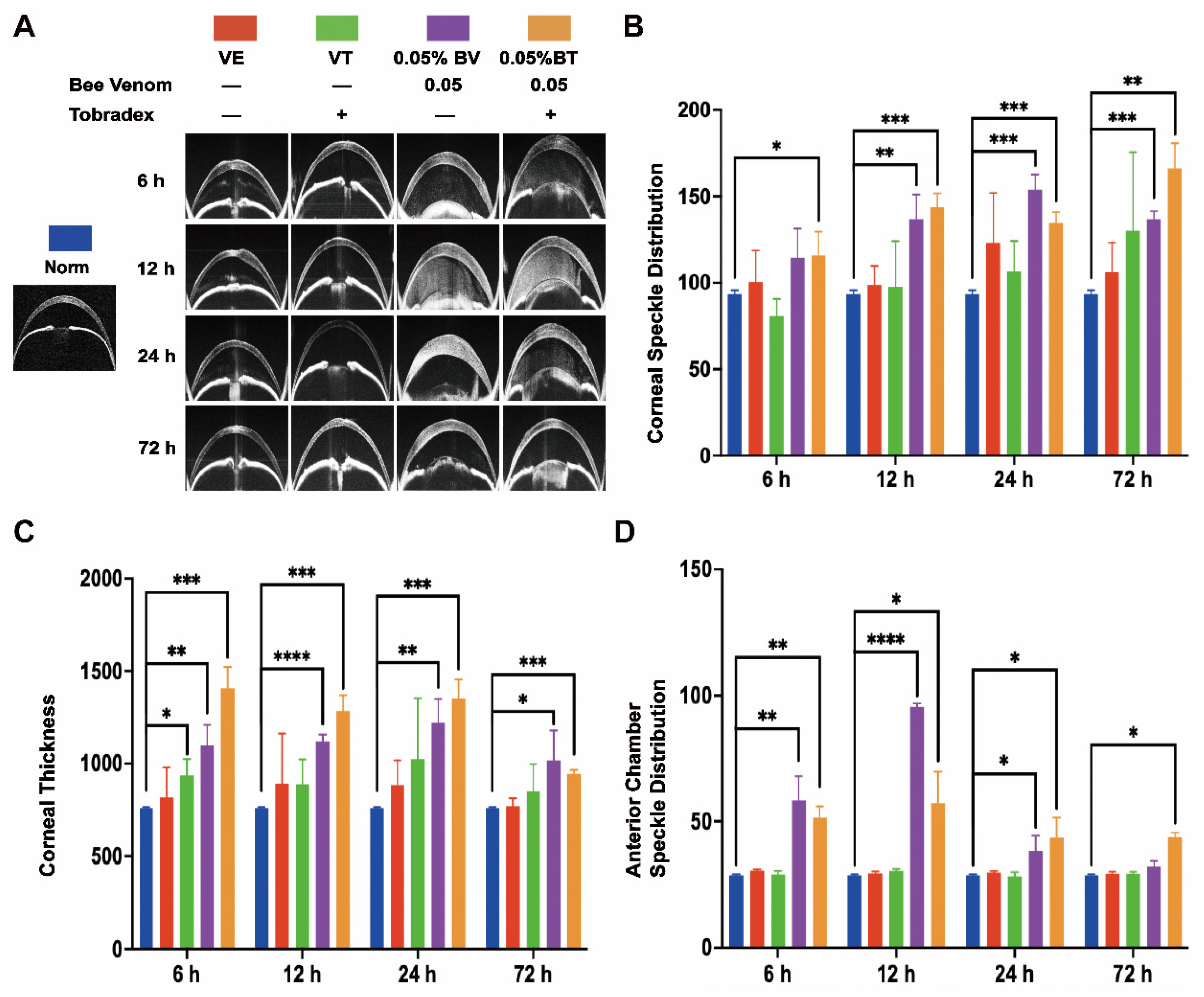
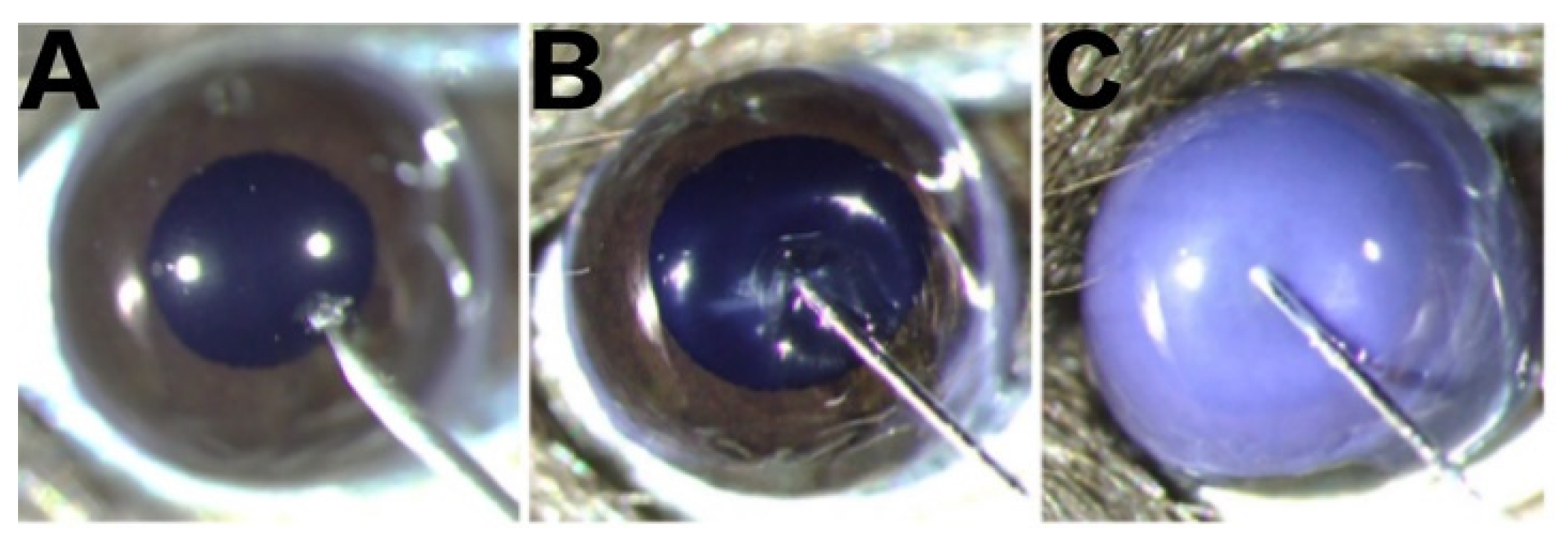
2.1. Morphological Changes in the Cornea
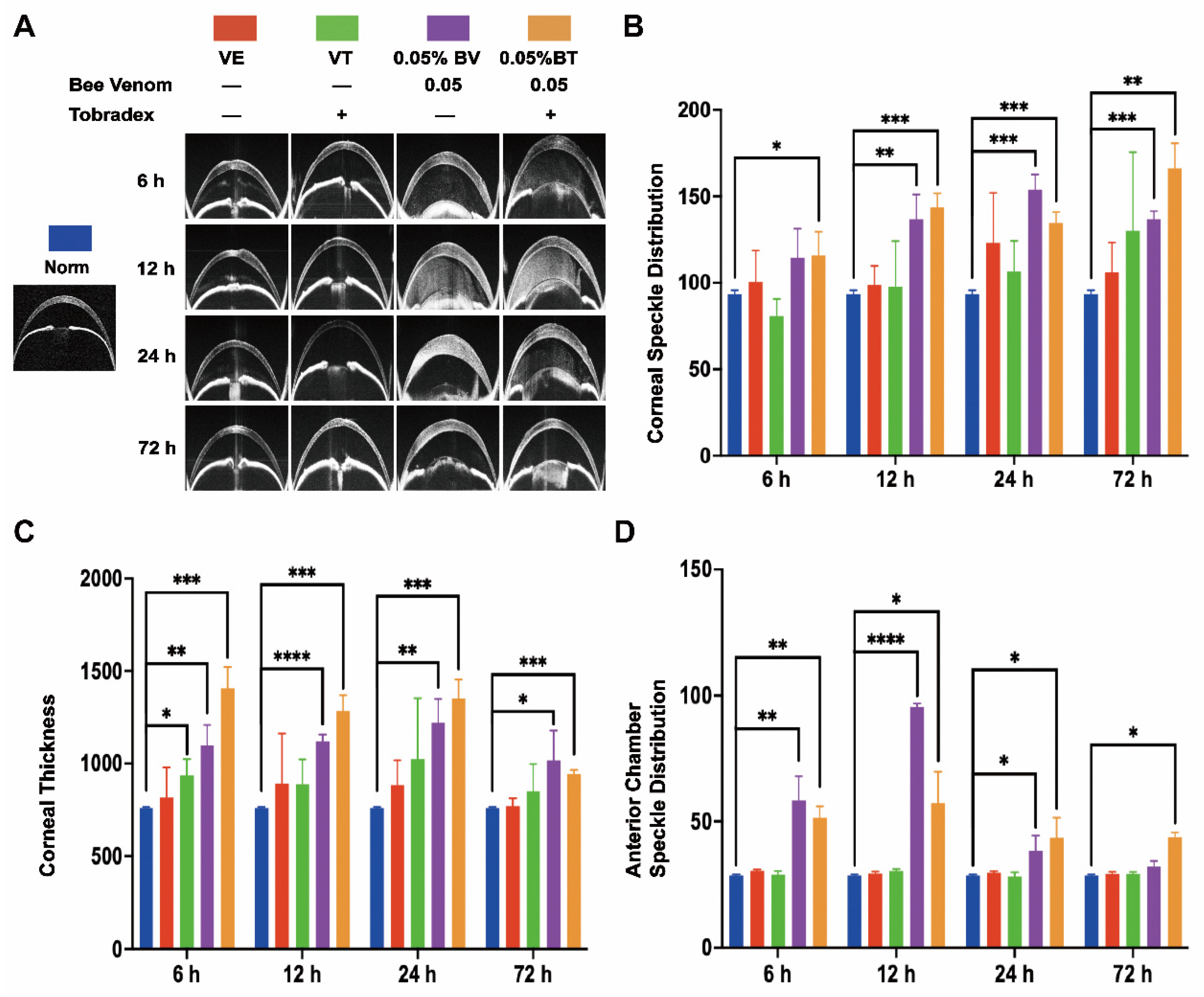
2.2. Morphological Changes in the Anterior Chamber
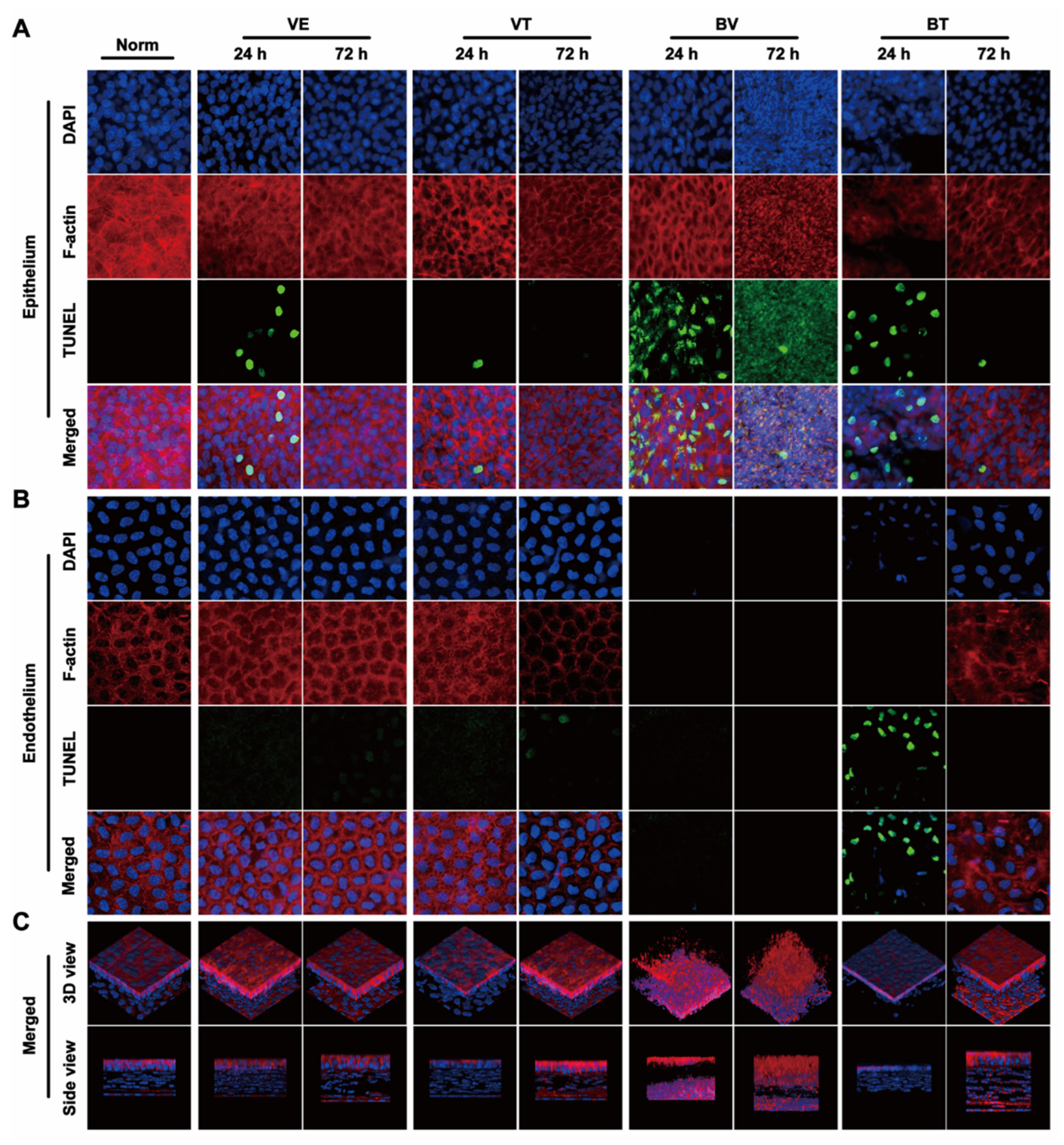
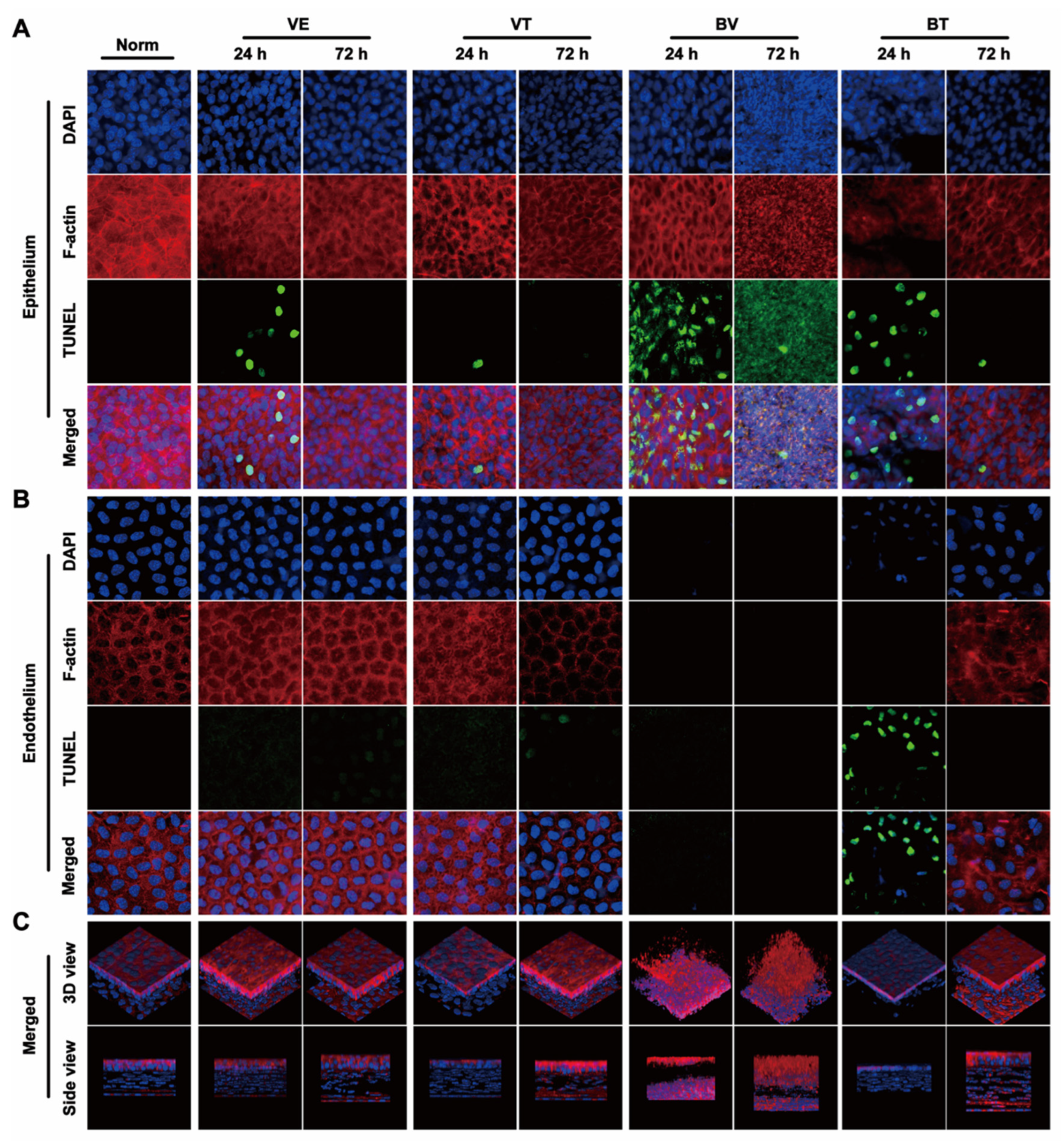
2.3. Apoptosis in the Cornea
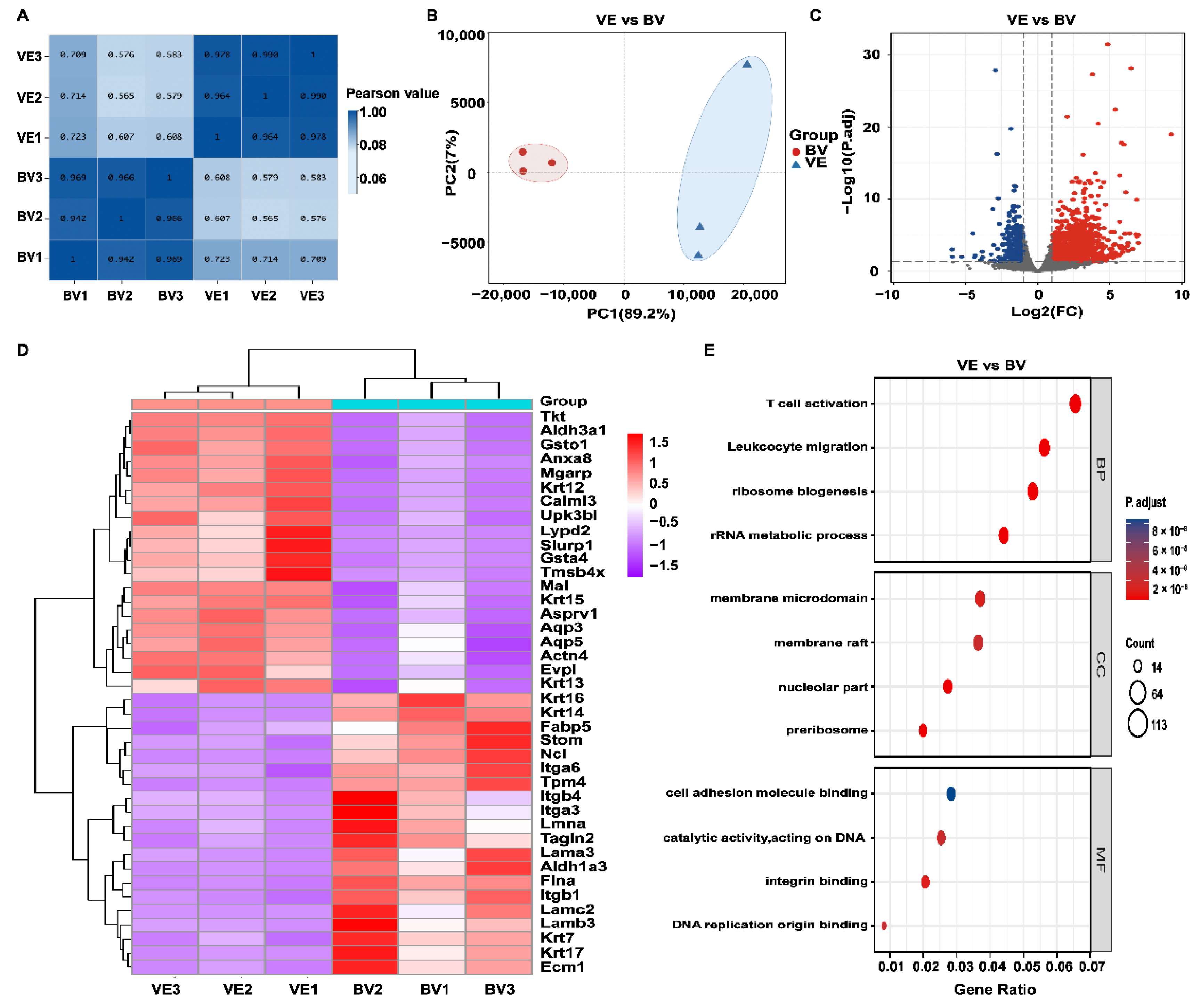
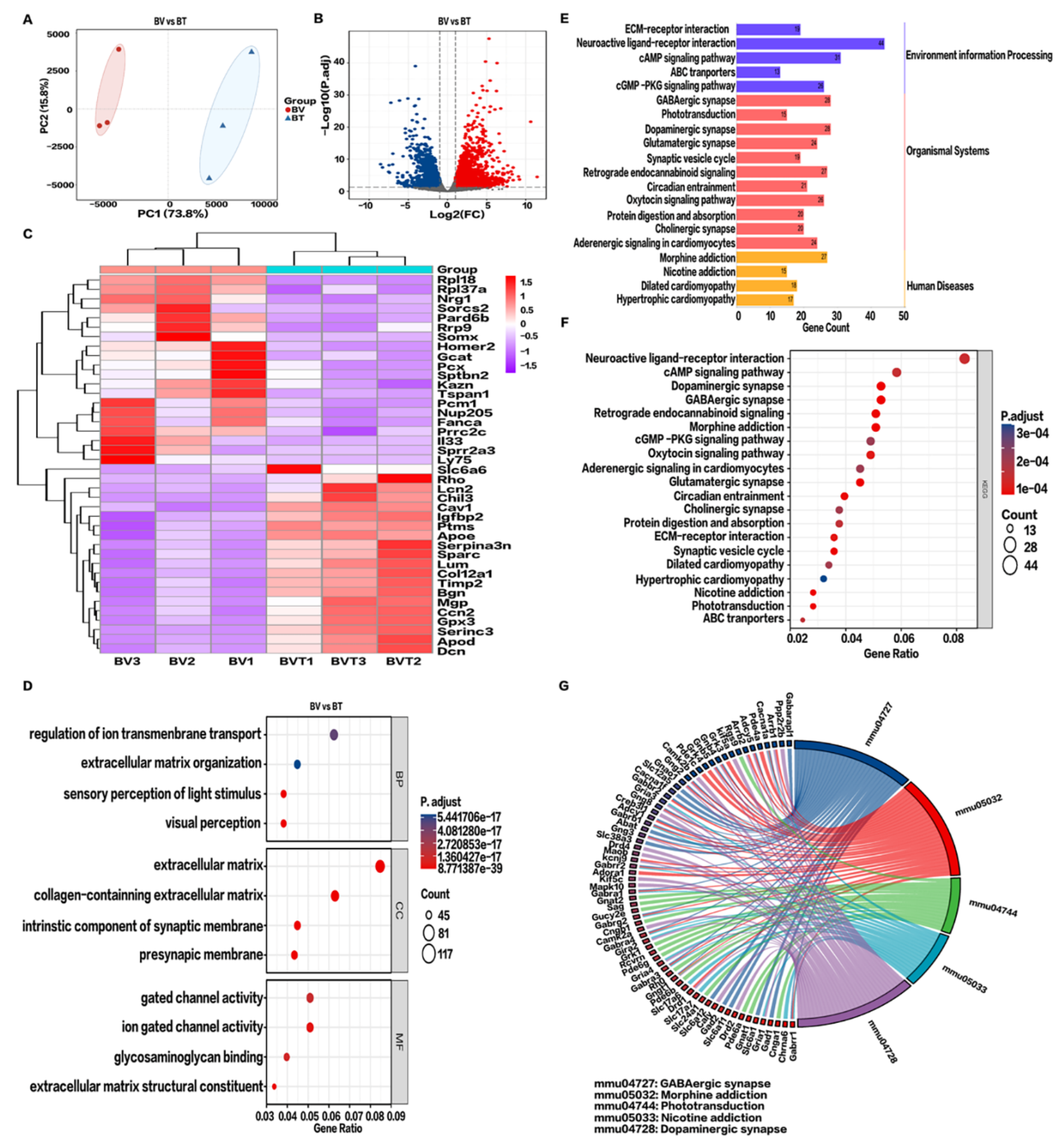
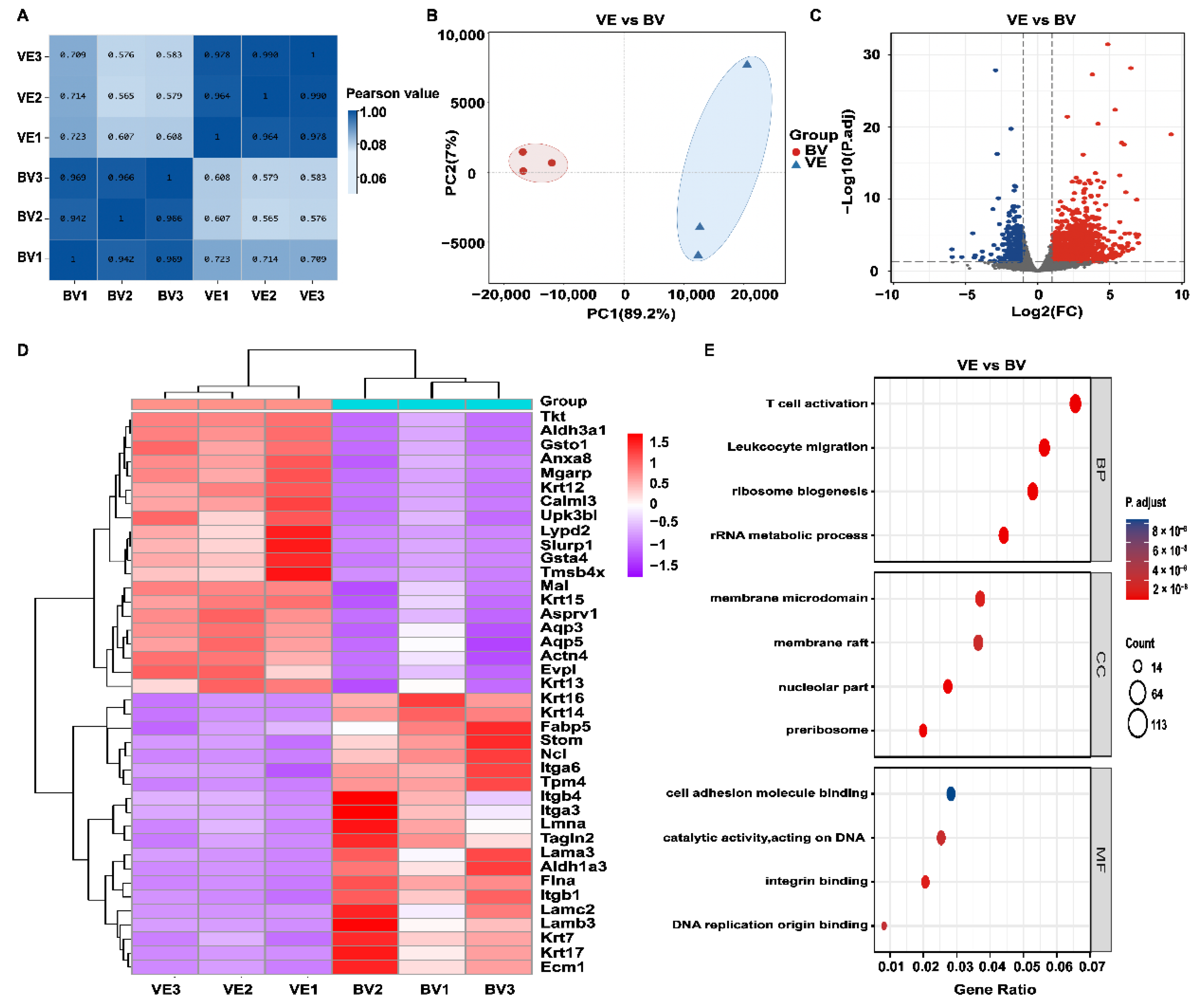
2.4. Transcriptomics Analysis in the BV Group
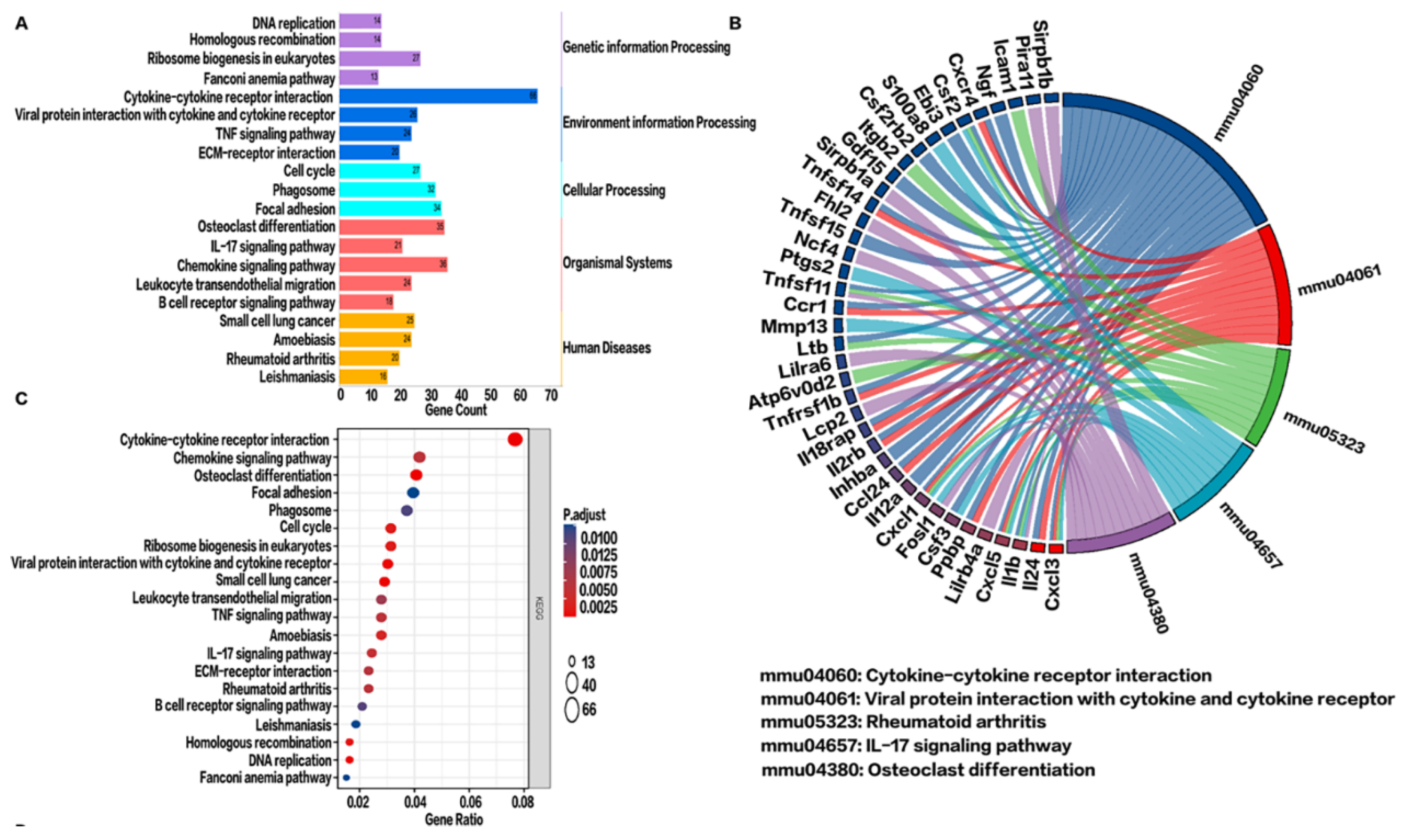
2.5. KEGG Pathway Enrichment Analysis in the BV Group
2.6. RNA-Seq Analysis following Tobradex® Treatment of CBS
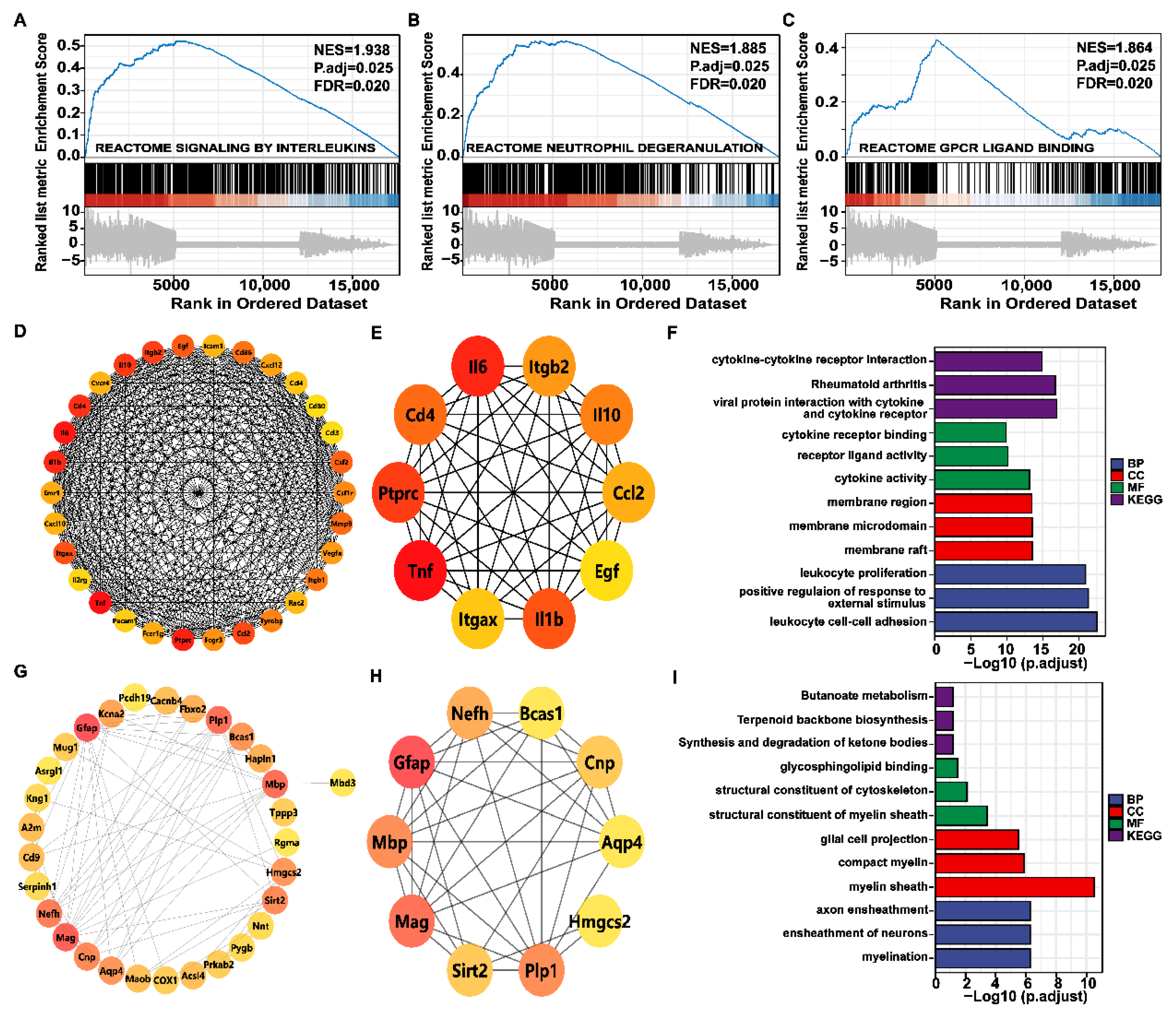
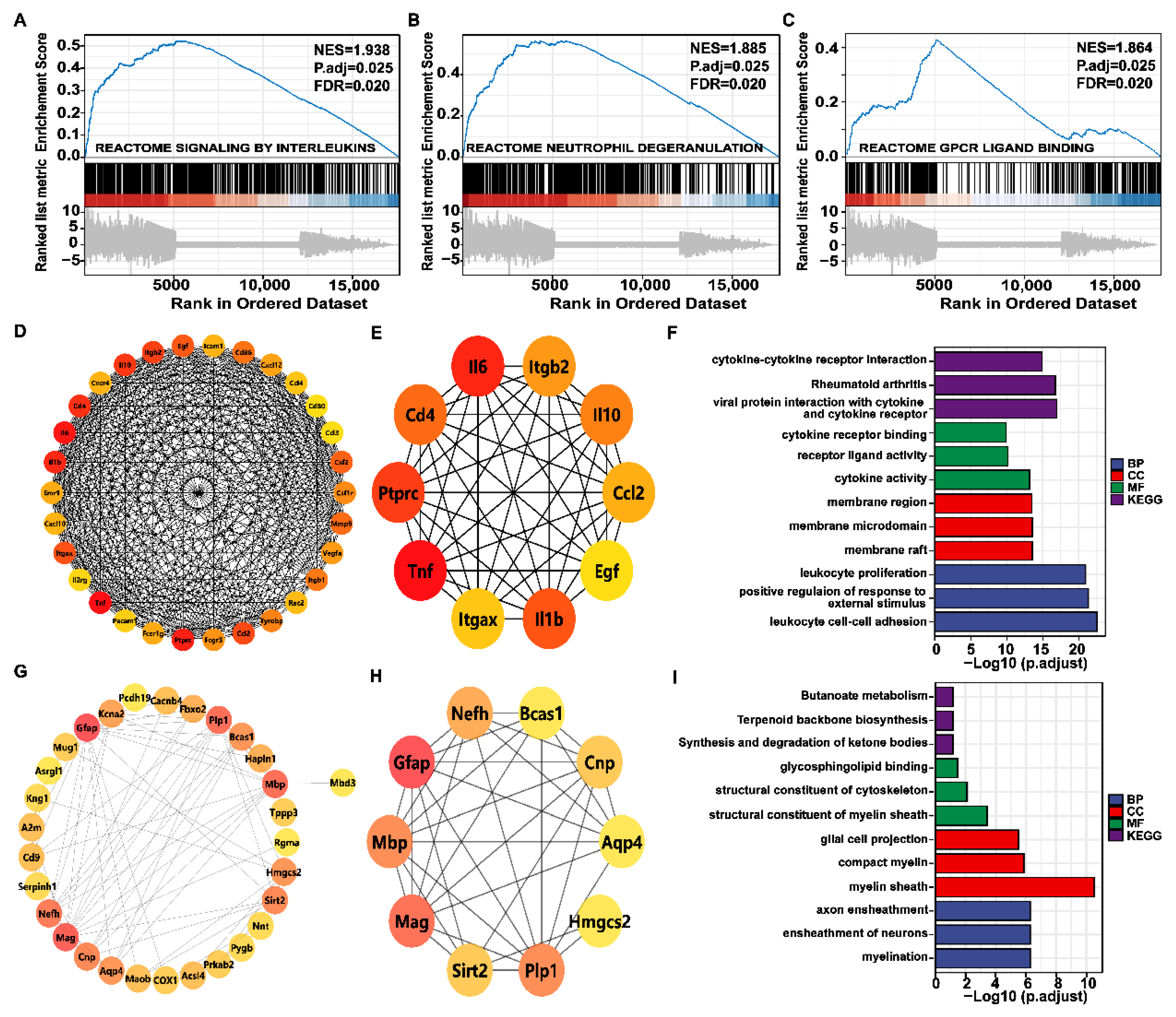
2.7. Genome-Wide Gene Set Enrichment Analysis (GSEA) and PPI Network
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals
5.2. Preparation of the BV Solution
5.3. Establishment of the Corneal Bee Sting Model
5.4. Slit-Lamp Microscopic Examination
5.5. Optical Coherence Tomography
5.6. Preparation of Corneal Whole-Mounts
5.7. F-Actin Staining of Corneal Whole-Mounts
5.8. TUNEL Assay of Corneal Whole-Mounts
5.9. Whole-Genome mRNA Sequencing and Bioinformatics Analysis
5.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kulhari, A.; Rogers, A.; Wang, H.; Kumaraswamy, V.M.; Xiong, W.; DeGeorgia, M. Ischemic Stroke After Wasp Sting. J. Emerg. Med. 2016, 51, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Pucca, M.B.; Cerni, F.A.; Oliveira, I.S.; Jenkins, T.P.; Argemí, L.; Sørensen, C.V.; Ahmadi, S.; Barbosa, J.E.; Laustsen, A.H. Bee Updated: Current Knowledge on Bee Venom and Bee Envenoming Therapy. Front. Immunol. 2019, 10, 2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhuiyan, M.A.A.; Agrawal, P.; Wadhwaniya, S.; Li, Q.; Alonge, O.; Rahman, A.F.; Rahman, A. Animal-related injuries and fatalities: Evidence from a large-scale population-based cross-sectional survey in rural Bangladesh. BMJ Open 2019, 9, e030039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratnayake, G.M.; Weerathunga, P.N.; Dilrukshi, M.S.A.; Amara Witharana, E.W.R.; Jayasinghe, S. Giant honey bee (Apis dorsata) sting and acute limb ischemia: A case report and review of the literature. BMC Res. Notes 2018, 11, 327. [Google Scholar] [CrossRef]
- Silva, G.B.D.; Vasconcelos, A.G.; Rocha, A.M.T.; Vasconcelos, V.R.D.; Barros, J.D.; Fujishima, J.S.; Ferreira, N.B.; Barros, E.J.G.; Daher, E.D.F. Acute kidney injury complicating bee stings—A review. Rev. Inst. Med. Trop. Sao Paulo 2017, 59, e25. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.N.; Kumar, B.K.; Velappan, P.; Sudheer, M.D. Possible complication of bee stings and a review of the cardiac effects of bee stings. BMJ Case Rep. 2016, 2016, bcr2015213974. [Google Scholar] [CrossRef]
- Smallheer, B.A. Bee and Wasp Stings: Reactions and Anaphylaxis. Crit. Care Nurs. Clin. N. Am. 2013, 25, 151–164. [Google Scholar] [CrossRef]
- Steen, C.J.; Janniger, C.K.; Schutzer, S.E.; Schwartz, R.A. Insect sting reactions to bees, wasps, and ants. Int. J. Dermatol. 2005, 44, 91–94. [Google Scholar] [CrossRef]
- Feás, X.; Vidal, C.; Remesar, S. What We Know about Sting-Related Deaths? Human Fatalities Caused by Hornet, Wasp and Bee Stings in Europe (1994–2016). Biology 2022, 11, 282. [Google Scholar] [CrossRef]
- Danforth, B.N.; Sipes, S.; Fang, J.; Brady, S.G. The history of early bee diversification based on five genes plus morphology. Proc. Natl. Acad. Sci. USA 2006, 103, 15118–15123. [Google Scholar] [CrossRef] [Green Version]
- Ilyasov, R.A.; Lee, M.-L.; Takahashi, J.-I.; Kwon, H.W.; Nikolenko, A.G. A revision of subspecies structure of western honey bee Apis mellifera. Saudi J. Biol. Sci. 2020, 27, 3615–3621. [Google Scholar] [CrossRef] [PubMed]
- Kurek-Górecka, A.; Komosinska-Vassev, K.; Rzepecka-Stojko, A.; Olczyk, P. Bee Venom in Wound Healing. Molecules 2020, 26, 248. [Google Scholar] [CrossRef] [PubMed]
- Kim, W. Bee Venom and Its Sub-Components: Characterization, Pharmacology, and Therapeutics. Toxins 2021, 13, 191. [Google Scholar] [CrossRef]
- Carpena, M.; Nuñez-Estevez, B.; Soria-Lopez, A.; Simal-Gandara, J. Bee Venom: An Updating Review of Its Bioactive Molecules and Its Health Applications. Nutrients 2020, 12, 3360. [Google Scholar] [CrossRef]
- Khalil, A.; Elesawy, B.H.; Ali, T.M.; Ahmed, O.M. Bee venom: From venom to drug. Molecules 2021, 26, 4941. [Google Scholar] [CrossRef]
- Chen, J.; Guan, S.M.; Sun, W.; Fu, H. Melittin, the Major Pain-Producing Substance of Bee Venom. Neurosci. Bull. 2016, 32, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Zolfagharian, H.; Mohajeri, M.; Babaie, M. Honey Bee Venom (Apis mellifera) Contains Anticoagulation Factors and Increases the Blood-clotting Time. J. Pharmacopunct. 2015, 18, 7–11. [Google Scholar] [CrossRef]
- Elieh Ali Komi, D.; Shafaghat, F.; Zwiener, R.D. Immunology of Bee Venom. Clin. Rev. Allergy Immunol. 2018, 54, 386–396. [Google Scholar] [CrossRef]
- Kreil, G. Hyaluronidases—A group of neglected enzymes. Protein Sci. 1995, 4, 1666–1669. [Google Scholar] [CrossRef] [Green Version]
- Moreno, M.; Giralt, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin, apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar] [CrossRef] [Green Version]
- Horta, C.C.R.; Magalhaes, B.D.F.; Oliveira-Mendes, B.B.R.; Carmo, A.O.D.; Duarte, C.G.; Felicori, L.F.; Machado-de-Avila, R.A.; Chavez-Olortegui, C.; Kalapothakis, E. Molecular, Immunological, and Biological Characterization of Tityus serrulatus Venom Hyaluronidase: New Insights into Its Role in Envenomation. PLoS Negl. Trop. Dis. 2014, 8, e2693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boldrini-Franca, J.; Cologna, C.T.; Pucca, M.B.; Bordon, K.D.C.F.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cerni, F.A.; Pinheiro-Junior, E.L.; et al. Minor snake venom proteins: Structure, function and potential applications. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2017, 1861, 824–838. [Google Scholar] [CrossRef] [PubMed]
- Cherubini, E.; Ben Ari, Y.; Gho, M.; Bidard, J.N.; Lazdunski, M. Long-term potentiation of synaptic transmission in the hippocampus induced by a bee venom peptide. Nature 1987, 328, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.Z.; Xu, M.; Sun, S. In Vivo Confocal Microscopy Observation of Cell and Nerve Density in Different Corneal Regions with Monocular Pterygium. J. Ophthalmol. 2020, 2020, 6506134. [Google Scholar] [CrossRef]
- Rhee, S.S.; Mah, F.S. Comparison of tobramycin 0.3%/dexamethasone 0.1% and tobramycin 0.3%/loteprednol 0.5% in the management of blepharo-keratoconjunctivitis. Adv. Ther. 2007, 24, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Al-Towerki, A.E. Corneal honeybee sting. Cornea 2003, 22, 672–674. [Google Scholar] [CrossRef]
- Ahmed, M.; Lee, C.S.; McMillan, B.; Jain, P.; Wiley, L.; Odom, J.V.; Leys, M. Predicting visual function after an ocular bee sting. Int. Ophthalmol. 2019, 39, 1621–1626. [Google Scholar] [CrossRef]
- Lin, P.-H.; Wang, N.-K.; Hwang, Y.-S.; Ma, D.H.-K.; Yeh, L.-K. Bee Sting of the Cornea and Conjunctiva: Management and Outcomes. Cornea 2011, 30, 392–394. [Google Scholar] [CrossRef]
- Gürlü, V.P.; Erda, N. Corneal bee sting-induced endothelial changes. Cornea 2006, 25, 981–983. [Google Scholar] [CrossRef]
- Chen, J.; Chen, H.S. Pivotal role of capsaicin-sensitive primary afferents in development of both heat and mechanical hyperalgesia induced by intraplantar bee venom injection. Pain 2001, 91, 367–376. [Google Scholar] [CrossRef]
- Chen, J.; Luo, C.; Li, H.L.; Chen, H.S. Primary hyperalgesia to mechanical and heat stimuli following subcutaneous bee venom injection into the plantar surface of hindpaw in the conscious rat: A comparative study with the formalin test. Pain 1999, 83, 67–76. [Google Scholar] [CrossRef]
- Chen, J.; Larivierec, W.R. The nociceptive and anti-nociceptive effects of bee venom injection and therapy: A double-edged sword. Prog. Neurobiol. 2010, 92, 151–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oršolić, N. Bee venom in cancer therapy. Cancer Metastasis Rev. 2012, 31, 173–194. [Google Scholar] [CrossRef] [PubMed]
- Han, S.M.; Lee, G.G.; Park, K.K. Acute dermal toxicity study of bee venom (Apis mellifera L.) in rats. Toxicol. Res. 2012, 28, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Han, S.M.; Lee, K.G.; Park, K.K.; Pak, S.C. Skin sensitization study of bee venom (Apis mellifera L.) in Guinea pigs and rats. Cutan. Ocul. Toxicol. 2013, 32, 27–30. [Google Scholar] [CrossRef]
- Han, S.M.; Lee, K.G.; Yeo, J.H.; Pak, S.C. Dermal and ocular irritation studies of honeybee (Apis mellifera L.) venom. Am. J. Chin. Med. 2012, 40, 795–800. [Google Scholar] [CrossRef]
- Mendonca, T.M.; Rodrigues, G.R. Corneal Bee Sting. N. Engl. J. Med. 2020, 383, 2274. [Google Scholar] [CrossRef]
- Olivo Payne, A.; Chong, E. Bee sting to the cornea: Toxic effects and management. Med. J. Aust. 2018, 209, 155. [Google Scholar] [CrossRef]
- Tyagi, M.; Reddy, S.; Basu, S.; Pappuru, R.R.; Dave, V.P. Endoscopic visualization-assisted corneal bee sting removal. Indian J. Ophthalmol. 2021, 69, 423–425. [Google Scholar] [CrossRef]
- Belmonte, C.; Acosta, M.C.; Merayo-Lloves, J.; Gallar, J. What Causes Eye Pain? Curr. Ophthalmol. Rep. 2015, 3, 111–121. [Google Scholar] [CrossRef] [Green Version]
- PParsadaniantz, S.M.; Rivat, C.; Rostène, W.; Goazigo, A.R.L. Opioid and chemokine receptor crosstalk: A promising target for pain therapy? Nat. Rev. Neurosci. 2015, 16, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Meves, H. Arachidonic acid and ion channels: An update. Br. J. Pharmacol. 2008, 155, 4–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khasabova, I.A.; Simone, D.A.; Seybold, V.S. Cannabinoids attenuate depolarization-dependent Ca2+ influx in intermediate-size primary afferent neurons of adult rats. Neuroscience 2002, 115, 613–625. [Google Scholar] [CrossRef]
- Goudet, C.; Magnaghi, V.; Landry, M.; Nagy, F.; Gereau IV, R.W.; Pin, J.P. Metabotropic receptors for glutamate and GABA in pain. Brain Res. Rev. 2009, 60, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Hucho, T.; Levine, J.D. Signaling Pathways in Sensitization: Toward a Nociceptor Cell Biology. Neuron 2007, 55, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.H.; Hou, J.F.; Liu, M.G.; Li, M.M.; Cui, X.Y.; Lu, Z.M.; Zhang, F.K.; An, Y.Y.; Shi, L.; Chen, J. Imbalance between excitatory and inhibitory amino acids at spinal level is associated with maintenance of persistent pain-related behaviors. Pharmacol. Res. 2009, 59, 290–299. [Google Scholar] [CrossRef]
- Launay, P.S.; Reboussin, E.; Liang, H.; Kessal, K.; Godefroy, D.; Rostene, W.; Sahel, J.A.; Baudouin, C.; Parsadaniantz, S.M.; Le Goazigo, A.R. Ocular inflammation induces trigeminal pain, peripheral and central neuroinflammatory mechanisms. Neurobiol. Dis. 2016, 88, 16–28. [Google Scholar] [CrossRef] [Green Version]
- Shamash, S.; Reichert, F.; Rotshenker, S. The cytokine network of wallerian degeneration: Tumor necrosis factor-α, interleukin-1α, and interleukin-1β. J. Neurosci. 2002, 22, 3052–3060. [Google Scholar] [CrossRef] [Green Version]
- Oktiansyah, R.; Juliandi, B.; Widayati, K.A.; Juniantito, V. Neuronal Cell Death and Mouse (Mus musculus) Behaviour Induced by Bee Venom. Trop. Life Sci. Res. 2018, 29, 1–11. [Google Scholar] [CrossRef]
- Djalilian, A.R.; Nagineni, C.N.; Mahesh, S.P.; Smith, J.A.; Nussenblatt, R.B.; Hooks, J.J. Inhibition of inflammatory cytokine production in human corneal cells by dexamethasone, but not cyclosporin. Cornea 2006, 25, 709–714. [Google Scholar] [CrossRef]
- Blackwell, G.J.; Carnuccio, R.; Rosa, M.D.; Flower, R.J.; Parente, L.; Persico, P. Macrocortin: A polypeptide causing the anti-phospholipase effect of glucocorticoids. Nature 1980, 287, 147–149. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Zha, J.; Shi, Y.; Li, Y.; Yuan, D.; Chen, Q.; Lin, F.; Fang, Z.; Yu, Y.; Dai, Y.; et al. Co-inhibition of HDAC and MLL-menin interaction targets MLL-rearranged acute myeloid leukemia cells via disruption of DNA damage checkpoint and DNA repair. Clin. Epigenet. 2019, 11, 137. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Kang, H.; Jin, M.; Wang, G.; Ma, W.; Liu, Z.; Xue, Y.; Li, C. Phenotypic and Transcriptomics Analyses Reveal Underlying Mechanisms in a Mouse Model of Corneal Bee Sting. Toxins 2022, 14, 468. https://doi.org/10.3390/toxins14070468
Wang Y, Kang H, Jin M, Wang G, Ma W, Liu Z, Xue Y, Li C. Phenotypic and Transcriptomics Analyses Reveal Underlying Mechanisms in a Mouse Model of Corneal Bee Sting. Toxins. 2022; 14(7):468. https://doi.org/10.3390/toxins14070468
Chicago/Turabian StyleWang, Yanzi, Honghua Kang, Mengyi Jin, Guoliang Wang, Weifang Ma, Zhen Liu, Yuhua Xue, and Cheng Li. 2022. "Phenotypic and Transcriptomics Analyses Reveal Underlying Mechanisms in a Mouse Model of Corneal Bee Sting" Toxins 14, no. 7: 468. https://doi.org/10.3390/toxins14070468
APA StyleWang, Y., Kang, H., Jin, M., Wang, G., Ma, W., Liu, Z., Xue, Y., & Li, C. (2022). Phenotypic and Transcriptomics Analyses Reveal Underlying Mechanisms in a Mouse Model of Corneal Bee Sting. Toxins, 14(7), 468. https://doi.org/10.3390/toxins14070468