Assessment of the Potential of a Native Non-Aflatoxigenic Aspergillus flavus Isolate to Reduce Aflatoxin Contamination in Dairy Feed

 ,
,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
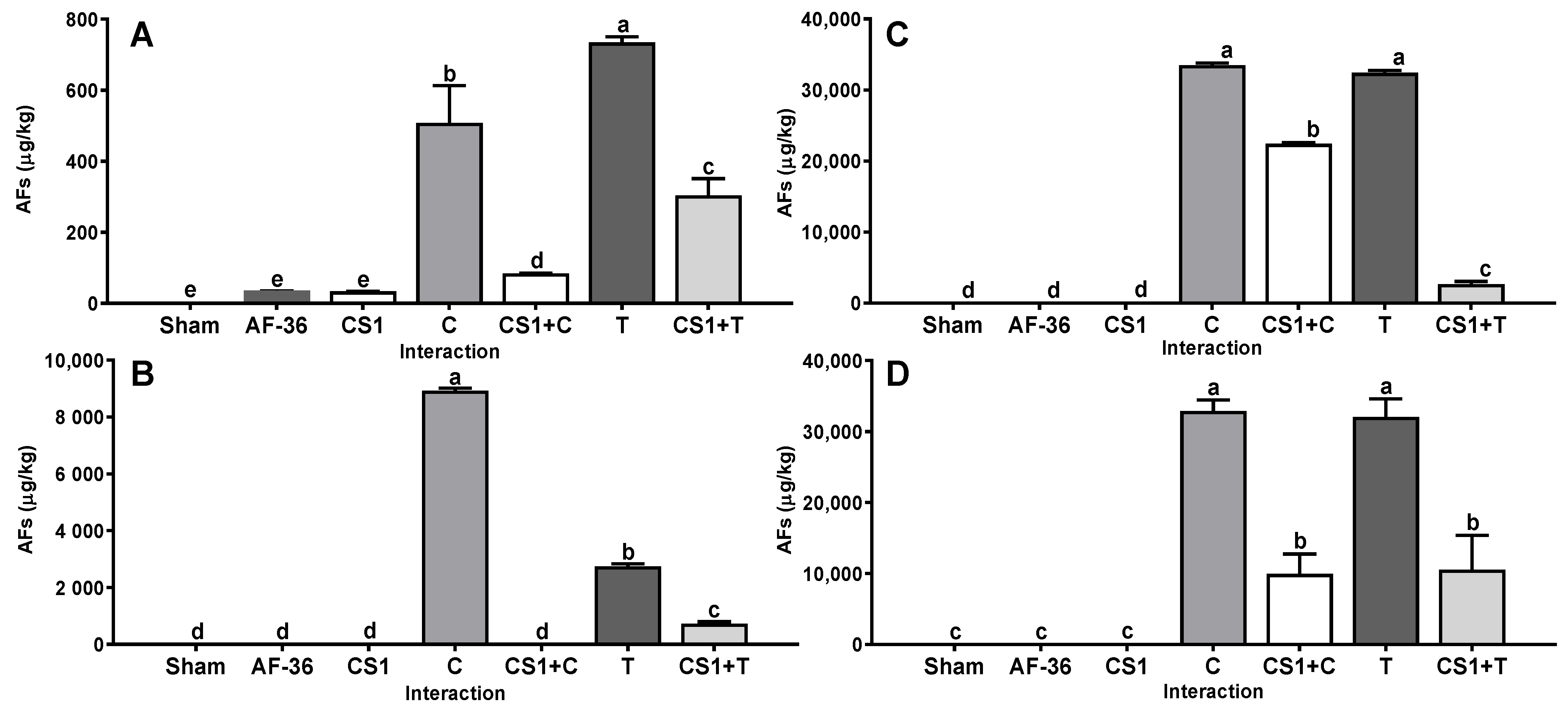
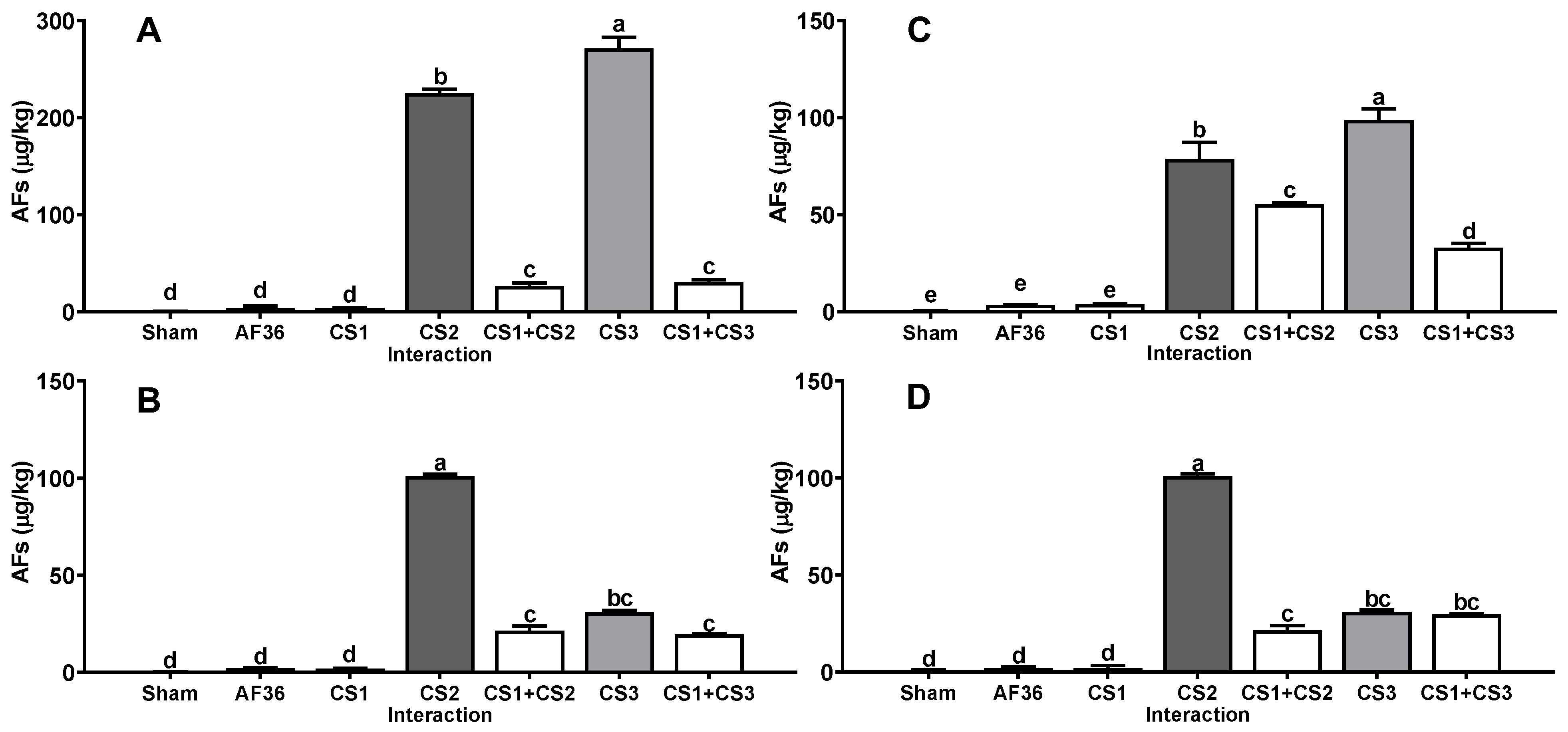
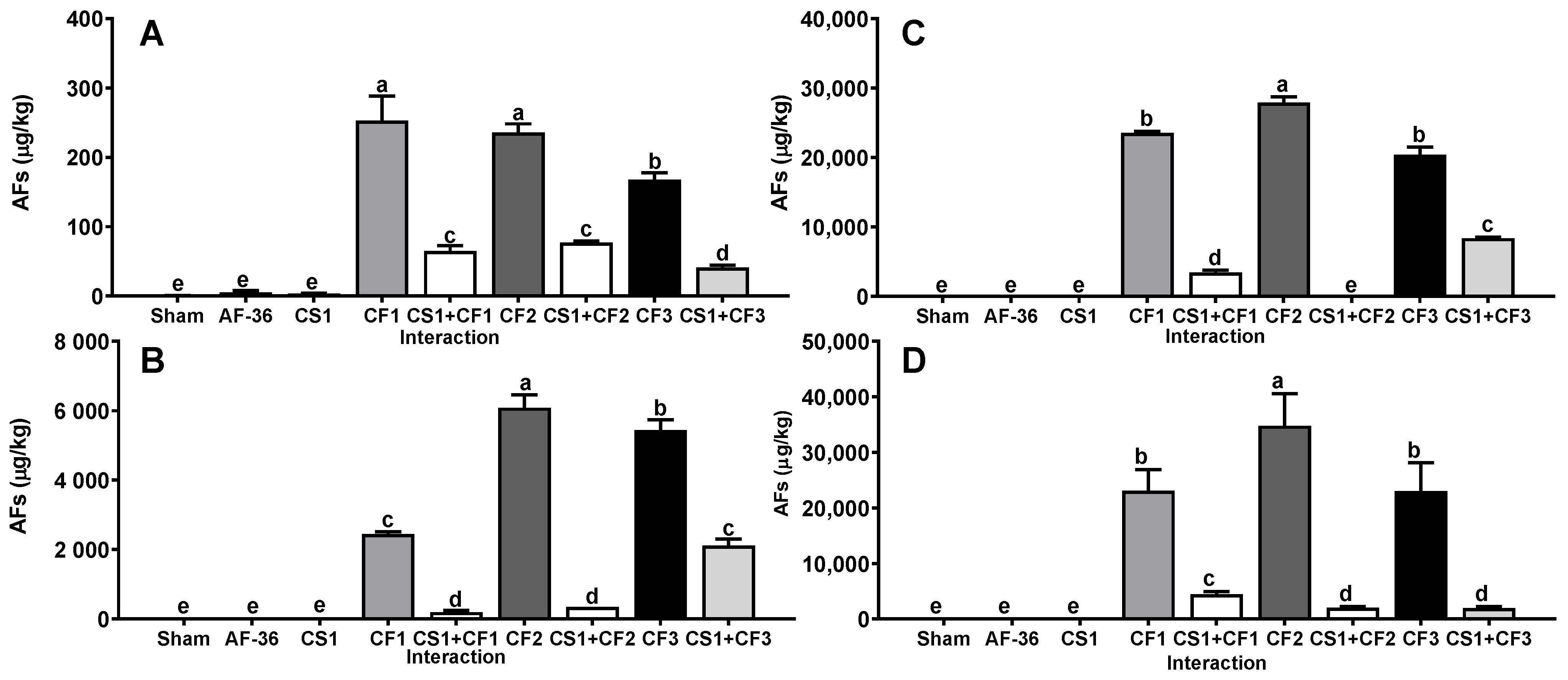
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fungal Strains
5.2. Treatment Design
5.3. In Situ Inoculation
5.4. Immature Kernels
5.5. Corn Silage
5.6. In Vitro Culture
5.7. Quantification of Aflatoxins
5.8. Molecular Analysis
5.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peles, F.; Sipos, P.; Kovács, S.; Györi, Z.; Pócsi, I.; Pusztahelyi, T. Biological Control and Mitigation of Aflatoxin Contamination in Commodities. Toxins 2021, 13, 104. [Google Scholar] [CrossRef] [PubMed]
- Omara, T.; Nassazi, W.; Omute, T.; Awath, A.; Laker, F.; Kalukusu, R.; Musau, B.; Nakabuye, B.V.; Kagoya, S.; Otim, G.; et al. Aflatoxins in Uganda: An Encyclopedic Review of the Etiology, Epidemiology, Detection, Quantification, Exposure Assessment, Reduction, and Control. Int. J. Microbiol. 2020, 2020, 4723612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Valdivia, E.; Valdivia-Flores, A.G.; Cruz-Vázquez, C.; Martínez-Saldaña, M.C.; Quezada-Tristán, T.; Rangel-Muñoz, E.J.; Ortiz-Martinez, R.; Medina-Esparza, L.E.; Jaramillo-Juarez, F. Diagnosis of Subclinical Aflatoxicosis by Biochemical Changes in Dairy Cows Under Field Conditions. Pak. Vet. J. 2020, 41, 2074–7764. [Google Scholar] [CrossRef]
- Walte, H.G.; Schwake-Anduschus, C.; Geisen, R.; Fritsche, J. Aflatoxin: Food Chain Transfer from Feed to Milk. J. Fur. Ver-Braucherschutz Leb. 2016, 11, 295–297. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Tozcano, L.J.; Juárez-Atonal, R.; Nava Galicia, S.B.; Garrido-Bazán, V.; Bibbins-Martínez, M. Aflatoxins, A Silent Danger? Front. Biotecnol. 2021, 20, 16–20. Available online: https://www.revistafronterabiotecnologica.cibatlaxcala.ipn.mx/volumen/vol20/pdf/Revista-diciembre-2021.pdf (accessed on 12 May 2022).
- Ogunade, I.M.; Martinez-Tuppia, C.; Queiroz, O.C.M.; Jiang, Y.; Drouin, P.; Wu, F.; Vyas, D.; Adesogan, A.T. Silage review: Mycotoxins in silage: Occurrence, effects, prevention, and mitigation. J. Dairy Sci. 2018, 101, 4034–4059. [Google Scholar] [CrossRef]
- De Luna-López, M.C.; Valdivia-Flores, A.G.; Jaramillo-Juárez, F.; Reyes, J.L.; Ortiz-Martínez, R.; Quezada-Tristán, T. Association between Aspergillus flavus Colonization and Aflatoxins Production in Immature Grains of Maize Genotypes. J. Food Sci. Eng. 2013, 3, 688–698. [Google Scholar] [CrossRef]
- Di Gregorio, M.C.; de Neeff, D.V.; Jager, A.V.; Corassin, C.H.; de Pinho, A.C.; de Albuquerque, R.; de Azevedo, A.C.; Fernandes, C.A. Mineral adsorbents for prevention of mycotoxins in animal feeds. Toxin Rev. 2014, 33, 125–135. [Google Scholar] [CrossRef]
- Martínez-Martínez, L.; Valdivia-Flores, A.G.; Guerrero-Barrera, A.L.; Quezada-Tristán, T.; Rangel-Muñoz, E.J.; Ortiz-Martínez, R. Toxic Effect of Aflatoxins in Dogs Fed Contaminated Commercial Dry Feed: A Review. Toxins 2021, 13, 65. [Google Scholar] [CrossRef]
- Valdivia, A.G.; Martinez, A.; Damian, F.J.; Quezada, T.; Ortiz, R.; Martinez, C.; Llamas, J.; Rodriguez, M.L.; Yamamoto, L.; Jaramillo, F.; et al. Efficacy of N-acetylcysteine to reduce the effects of aflatoxin B1 intoxication in broiler chickens. Poult. Sci. 2001, 80, 727–734. [Google Scholar] [CrossRef]
- Damiano, S.; Jarriyawattanachaikul, W.; Girolami, F.; Longobardi, C.; Nebbia, C.; Andretta, E.; Lauritano, C.; Dabbou, S.; Avantaggiato, G.; Schiavone, A.; et al. Curcumin Supplementation Protects Broiler Chickens Against the Renal Oxidative Stress Induced by the Dietary Exposure to Low Levels of Aflatoxin B1. Front. Vet. Sci. 2022, 8, 822227. [Google Scholar] [CrossRef]
- Krishnamurthy, R.; Padma, P.R.; Dhandapani, K. Antagonistic efficiency of Aspergillus giganteus as a Biocontrol Agent Against Aflatoxigenic Aspergillus flavus Infecting Maize. J. Pure Appl. Microbiol. 2020, 14, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Moore, G.G. Practical Considerations will Ensure the Continued Success of Pre-harvest Biocontrol Using Non-aflatoxigenic Aspergillus flavus Strains. Crit. Rev. Food Sci. Nutr. 2021, 62, 4208–4225. [Google Scholar] [CrossRef] [PubMed]
- Chanda, A.; Roze, L.V.; Linz, J.E. A Possible Role for Exocytosis in Aflatoxin Export in Aspergillus parasiticus. Eukaryot. Cell 2010, 9, 1724–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damann, K.E. Atoxigenic Aspergillus flavus Biological Control of Aflatoxin Contamination: What is the Mechanism? World Mycotoxin J. 2015, 8, 235–244. [Google Scholar] [CrossRef]
- Khan, R.; Ghazali, F.M.; Mahyudin, N.A.; Samsudin, N.I.P. Biocontrol of Aflatoxins Using Non-aflatoxigenic Aspergillus flavus: A Literature Review. J. Fungi 2021, 7, 381. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.K.; Chang, T.D.; Katoh, K. Deciphering the Origin of Aspergillus flavus NRRL21882, the Active Biocontrol Agent of Afla-Guard®. Lett. Appl. Microbiol. 2020, 72, 509–516. [Google Scholar] [CrossRef]
- Khan, R.; Ghazali, F.M.; Mahyudin, N.A.; Samsudin, N.I.P. Co-Inoculation of Aflatoxigenic and Non-aflatoxigenic Strains of Aspergillus flavus to Assess the Efficacy of Non-aflatoxigenic Strains in Growth Inhibition and Aflatoxin B1 Reduction. Agriculture 2021, 11, 198. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, M.; Xie, R.; Zhang, F.; Wang, S.; Pan, X.; Wang, S.; Zhuang, Z. The Methyltransferase AflSet1 is Involved in Fungal Morphogenesis, AFB1 Biosynthesis, and Virulence of Aspergillus flavus. Front. Microbiol. 2020, 11, 234. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Aziz, A.R.; Shehata, S.M.; Hisham, S.M.; Alobathani, A.A. Molecular Profile of Aflatoxigenic and Non-aflatoxigenic Isolates of Aspergillus flavus Isolated from Stored Maize. Saudi J. Biol. Sci. 2021, 28, 1383–1391. [Google Scholar] [CrossRef]
- Wang, N.; Tytell, J.D.; Ingber, D.E. Mechanotransduction at a Distance: Mechanically Coupling the Extracellular Matrix with the Nucleus. Nat. Rev. Mol. Cell Biol. 2009, 10, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Kumamoto, C.A. Molecular Mechanisms of Mechanosensing and their Roles in Fungal Contact Sensing. Nat. Rev. Microbiol. 2008, 6, 667–673. [Google Scholar] [CrossRef] [Green Version]
- Weaver, M.A.; Abbas, H.K. Field Displacement of Aflatoxigenic Aspergillus flavus Strains through Repeated Biological Control Applications. Front. Microbiol. 2019, 10, 1788. [Google Scholar] [CrossRef] [Green Version]
- Moore, G.G.; Lebar, M.D.; Carter-Wientjes, C.H. The Role of Extrolites Secreted by Non aflatoxigenic Aspergillus flavus in Biocontrol Efficacy. J. Appl. Microbiol. 2019, 126, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Muñoz, E.J.; Valdivia-Flores, A.G.; Moreno-Rico, O.; Hernández-Delgado, S.; Cruz-Vázquez, C.; De Luna-López, M.C.; Quezada-Tristán, T.; Ortiz-Martinez, R.; Máyek-Pérez, N. Characterization of Aspergillus flavus and Quantification of Aflatoxins in Feed and Raw Milk of Cows in Aguascalientes, Mexico. Rev. Mex. Cienc. Pecu. 2020, 11, 435–454. [Google Scholar] [CrossRef]
- Álvarez-Días, F.; Torres-Parga, B.; Valdivia-Flores, A.G.; Quezada-Tristán, T.; Alejos-De La Fuente, J.I.; Sosa-Ramírez, J.; Rangel-Muñoz, E.J. Aspergillus flavus and Total Aflatoxins Occurrence in Dairy Feed and Aflatoxin M1 in Bovine Milk in Aguascalientes, Mexico. Toxins 2022, 14, 292. [Google Scholar] [CrossRef]
- Ehrlich, K. Non-aflatoxigenic Aspergillus flavus to Prevent Aflatoxin Contamination in Crops: Advantages and Limitations. Front. Microbiol. 2014, 5, 50. [Google Scholar] [CrossRef]
- Bandyopadhyay, R.; Ortega-Beltran, A.; Akande, A.; Mutegi, C.; Atehnkeng, J.; Kaptoge, L.; Senghor, A.L.; Adhikari, B.N.; Cotty, P.J. Biological Control of Aflatoxins in Africa: Current Status and Potential Challenges in the Face of Climate Change. World Mycotoxin J. 2016, 9, 771–789. [Google Scholar] [CrossRef] [Green Version]
- Dorner, J.W. Efficacy of a Biopesticide for Control of Aflatoxins in Corn. J. Food Prot. 2010, 73, 495–499. [Google Scholar] [CrossRef]
- Alaniz-Zanon, M.S.; Clemente, M.P.; Chulze, S.N. Characterization and Competitive Ability of Non-aflatoxigenic Aspergillus flavus Isolated from the Maize Agro-ecosystem in Argentina as Potential Aflatoxin Biocontrol Agents. Int. J. Food Microbiol. 2018, 277, 58–63. [Google Scholar] [CrossRef]
- Yan, L.; Song, W.; Chen, Y.; Kang, Y.; Lei, Y.; Huai, D.; Wang, Z.; Wang, X.; Liao, B. Effect of Non-aflatoxigenic Strains of Aspergillus flavus on Aflatoxin Contamination of Pre-harvest Peanuts in Fields in China. Oil Crop Sci. 2021, 6, 81–86. [Google Scholar] [CrossRef]
- Agbetiameh, D.; Ortega-Beltran, A.; Awuah, R.T.; Atehnkeng, J.; Islam, M.S.; Callicott, K.A.; Cotty, P.J.; Bandyopadhyay, R. Potential of Atoxigenic Aspergillus flavus Vegetative Compatibility Groups Associated with Maize and Groundnut in Ghana as Biocontrol Agents for Aflatoxin Management. Front. Microbiol. 2019, 10, 2069. [Google Scholar] [CrossRef] [PubMed]
- Mallikarjunaiah, N.H.; Jayapala, N.; Puttaswamy, H.; Ramachandrappa, N.S. Characterization of Non-aflatoxigenic Strains of Aspergillus flavus as Potential Biocontrol Agent for the Management of Aflatoxin Contamination in Groundnut. Microb. Pathog. 2017, 102, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Abbas, H.; Zablotowicz, R.; Horn, B.; Phillips, N.; Johnson, B.; Jin, X.; Abel, C.A. Comparison of Major Biocontrol Strains of Non-aflatoxigenic Aspergillus flavus for the Reduction of Aflatoxins and Cyclopiazonic Acid in Maize. Food Addit. Contam. Part A 2011, 28, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Uka, V.; Moore, G.G.; Arroyo-Manzanares, N.; Nebija, D.; De Saeger, S.; Diana Di Mavungu, J. Unravelling the Diversity of the Cyclopiazonic acid Family of Mycotoxins in Aspergillus flavus by UHPLC triple-TOF HRMS. Toxins 2017, 9, 35. [Google Scholar] [CrossRef] [Green Version]
- Okoth, S.; De Boevre, M.; Vidal, A.; Diana Di Mavungu, J.; Landschoot, S.; Kyallo, M.; Njuguna, J.; Harvey, J.; De Saeger, S. Genetic and Toxigenic Variability within Aspergillus flavus Population Isolated from Maize in Two Diverse Environments in Kenya. Front. Microbiol. 2018, 9, 57. [Google Scholar] [CrossRef]
- Moore, G.G.; Elliott, J.L.; Singh, R.; Horn, B.W.; Dorner, J.W.; Stone, E.A.; Stone, E.A.; Chulze, S.N.; Barros, G.G.; Naik, M.K.; et al. Sexuality Generates Diversity in the Aflatoxin Gene Cluster: Evidence on a Global Scale. PLoS Pathog. 2013, 9, e1003574. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, L.A.; Callicott, K.A.; Bandyopadhyay, R.; Mehl, H.L.; Orbach, M.J.; Cotty, P.J. Degradation of Aflatoxins B1 by Atoxigenic Aspergillus flavus Biocontrol Agents. Plant Dis. 2021, 105, 2343–2350. [Google Scholar] [CrossRef]
- Sserumaga, J.P.; Ortega-Beltran, A.; Wagacha, J.M.; Mutegi, C.K.; Bandyopadhyay, R. Aflatoxin-Producing Fungi Associated with Pre-harvest Maize Contamination in Uganda. Int. J. Food Microbiol. 2020, 313, 108376. [Google Scholar] [CrossRef]
- King, E.D.; Bassi, A.B.; Ross, D.C.; Druebbisch, B. An Industry Perspective on the Use of “Atoxigenic” Strains of Aspergillus flavus as Biological Control Agents and the Significance of Cyclopiazonic Acid. Toxin Rev. 2011, 30, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Quezada-Viay, M.Y.; Flores-Olivas, A.; Arrúa-Alvarenga, A.A.; Vázquez-Badillo, M.E.; Moreno-Martínez, E. Corn Plant Resistance to Infection by Aspergillus flavus Link in Greenhouse conditions. Rev. Agrar.-Nueva Época 2011, 8, 15–20. Available online: https://revista.uaaan.edu.mx/wp-content/uploads/2021/09/2011-2.pdf#page=17 (accessed on 12 May 2022).
- Gallo, A.; Fancello, F.; Ghilardelli, F.; Zara, S.; Froldi, F.; Spanghero, M. Effects of Several Lactic Acid Bacteria Inoculants on Fermentation and Mycotoxins in Corn Silage. Anim. Feed Sci. Technol. 2021, 277, 114962. [Google Scholar] [CrossRef]
- Hernández-Martínez, J.L.; Carranza-Álvarez, C.; Maldonado-Miranda, J.J.; Martínez-Soto, D. Isolation of Fusarium from Vanilla Plants Grown in the Huasteca Potosina Mexico. Mex. J. Phytopathol. 2020, 38, 475–484. [Google Scholar] [CrossRef]
- Santibáñez-Escobar, R.; Martínez-Ibarra, J.A.; Tapia-González, J.M.; Avellaneda-Cevallos, J.H.; Hernández-Gallardo, M.; Montañez-Valdez, O.D. Identification and Quantification of Mycotoxigenic Fungi in Bovine Feed. Rev. Cienc. Tecnol. 2011, 4, 19–23. [Google Scholar] [CrossRef]
- Scott, P.M. Mycotoxin Methodology. Food Addit. Contam. 1995, 12, 395–403. [Google Scholar] [CrossRef]
- Hoffman, C.S.; Winston, F. A ten-minute DNA preparation from yeast efficiently releases autonomous plasmids for transformation of Escherichia coli. Gene 1987, 57, 267–272. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal Ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Application; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Shapira, R.; Paster, N.; Eyal, O.; Menasherov, M.; Mett, A.; Salomon, R. Detection of aflatoxinogenic molds in grains by PCR. Appl. Environ. Microbiol. 1996, 62, 3270–3273. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC168121/pdf/623270.pdf (accessed on 12 May 2022). [CrossRef] [Green Version]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
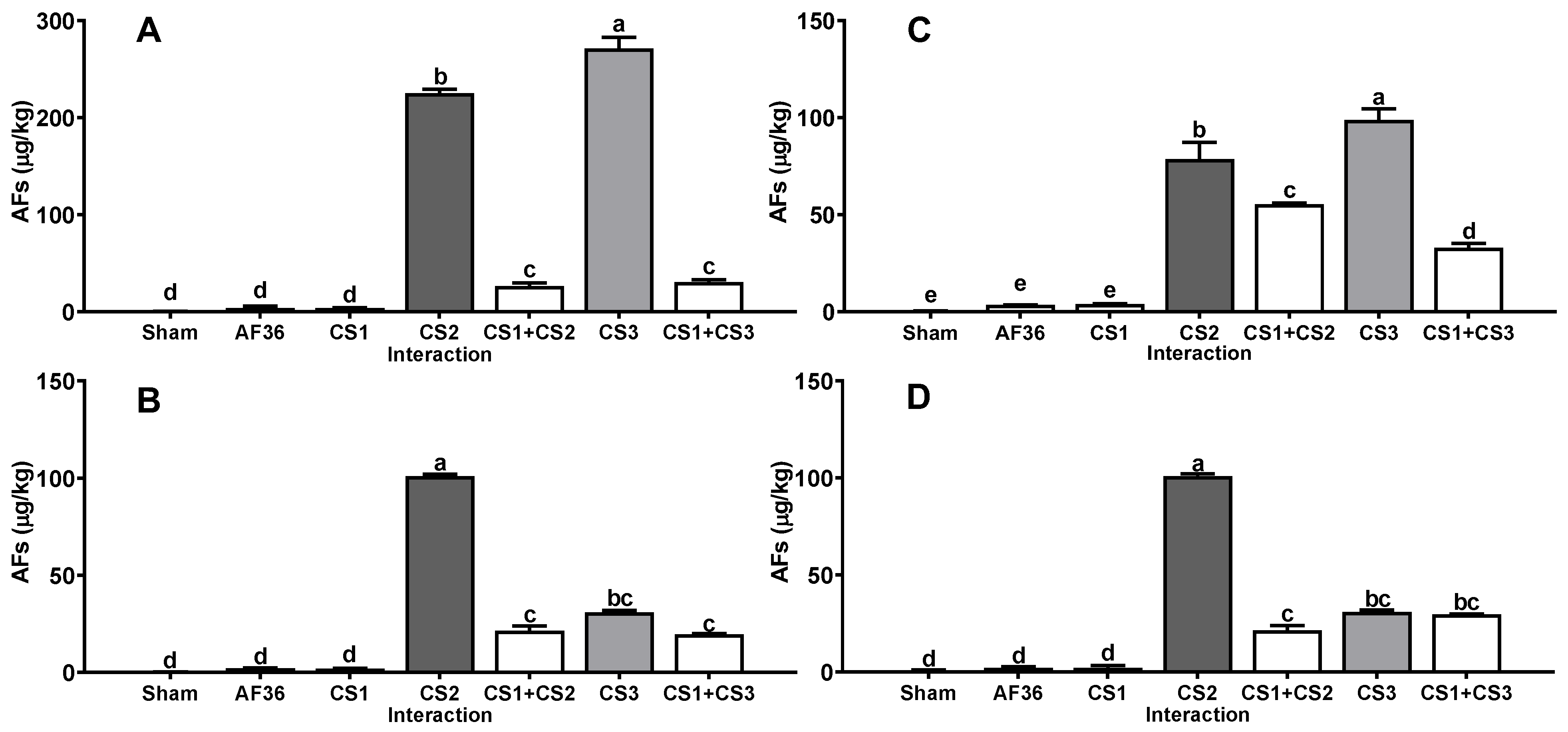{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CS 1 VS: | A. Whole Plant | B. Immature Grain | C. Corn Silage | D. Culture Media | General | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | CI 95% | Mean | CI 95% | Mean | CI 95% | Mean | CI 95% | Mean | CI 95% | |
| All strains | 76.0 A | 73.9–78.2 | 66.1 AB | 63.8–68.4 | 69.4 B | 65.1–73.7 | 62.5 AB | 57.4–67.6 | 68.5 | 66.9–70.1 |
| C | 79.1 ab | 73.5–84.8 | 33.0 d | 26.9–39.1 | 69.5 b | 68.4–70.5 | 70.7 a | 57.2–84.2 | 63.1 ab | 58.8–67.4 |
| T | 59.2 c | 53.5–64.9 | 91.8 ab | 85.7–97.9 | 65.6 b | 61.0–70.1 | 69.4 a | 55.9–82.9 | 71.5 ab | 67.2–75.8 |
| CF1 | 73.8 b | 68.1–79.5 | 85.3 b | 79.3–91.4 | 91.8 a | 89.4–94.2 | 79.8 a | 66.2–93.3 | 82.7 a | 78.4–87.0 |
| CF2 | 67.1 bc | 61.4–72.8 | 99.3 a | 93.2–100 | 94.3 a | 93.9–94.6 | 94.0 a | 80.5–100 | 88.7 a | 84.4–92.9 |
| CF3 | 75.7 ab | 70.1–81.4 | 58.7 c | 52.6–64.8 | 61.2 b | 68.5–70.3 | 90.8 a | 77.3–100 | 71.6 ab | 67.3–75.9 |
| CS2 | 88.4 a | 82.7–94.1 | 27.8 d | 21.7–33.9 | 70.4 b | 68.5–72.3 | 17.8 b | 4.3–31.3 | 51.1 b | 46.8–55.4 |
| CS3 | 88.9 a | 83.2–94.6 | 66.8 c | 60.7–72.9 | 33.1 c | 28.2–37.9 | 15.3 b | 1.8–28.8 | 51.0 b | 46.7–55.3 |
| R2 (%) * | 71.7 | 97.7 | 97.3 | 87.9 | 90.6 | |||||
| p-value | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |||||
| No. | Isolate | ID | Morphotype | Aflatoxins Production | Interaction |
|---|---|---|---|---|---|
| 1 | Control 1 | Sham | -- | -- | |
| 2 | AF-36 (Negative control) | AF36 | L | Negative | -- |
| 3 | Cuautitlán (positive control) | C | S | Positive | CS1 + C |
| 4 | Tamaulipas (positive control) | T | S | Positive | CS1 + T |
| 5 | Concentrate Feed- 1 | CF1 | S | Positive | CS1 + CF1 |
| 6 | Concentrate Feed-2 | CF2 | S | Positive | CS1 + CF2 |
| 7 | Concentrate Feed-3 | CF3 | S | Positive | CS1 + CF3 |
| 8 | Corn Silage-1 | CS1 | L | Negative | -- |
| 9 | Corn Silage-2 | CS2 | S | Positive | CS1 + CS2 |
| 10 | Corn Silage-3 | CS3 | S | Positive | CS1 + CS3 |
| Isolate | ITS-1 | ITS-4 | aflR-F | aflR-R | cmdA7-F | cmdA8-R |
|---|---|---|---|---|---|---|
| C | ON351284 | ON351498 | MN987040.1 *** | AF441434.1 *** | (NS) | (NS) |
| T | ON351288 | ON351503 | CP051029.1 *** | KY769956.1 *** | (NS) | (NS) |
| CF1 | HQ844707.1 *** | ON351496 | XM_041285628.1 *** | L32577.1 *** | CP051084.1 *** | MK119700.1 *** |
| CF2 | ON351282 | ON351497 | HQ844707.1 *** | MH511139.1 *** | CP051020.1 * | (NS) |
| CF3 | ON351283 | ON351501 | MH752564.1 ** | EF565462.1 *** | MN987032.1 *** | MK119699.1 *** |
| CS1 | ON351285 | ON351499 | --- | --- | CP044620.1 ** | CP051060.1 *** |
| CS2 | ON351286 | ON351500 | KY769955.1 *** | AF441432.1 *** | CP044622.1 ** | (NS) |
| CS3 | ON351287 | ON351502 | MG720232.1 ** | MH280087.1 ** | CP051036.1 *** | MK119701.1 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rangel-Muñoz, E.J.; Valdivia-Flores, A.G.; Hernández-Delgado, S.; Cruz-Vázquez, C.; de-Luna-López, M.C.; Quezada-Tristán, T.; Ortiz-Martínez, R.; Mayek-Pérez, N. Assessment of the Potential of a Native Non-Aflatoxigenic Aspergillus flavus Isolate to Reduce Aflatoxin Contamination in Dairy Feed. Toxins 2022, 14, 437. https://doi.org/10.3390/toxins14070437
Rangel-Muñoz EJ, Valdivia-Flores AG, Hernández-Delgado S, Cruz-Vázquez C, de-Luna-López MC, Quezada-Tristán T, Ortiz-Martínez R, Mayek-Pérez N. Assessment of the Potential of a Native Non-Aflatoxigenic Aspergillus flavus Isolate to Reduce Aflatoxin Contamination in Dairy Feed. Toxins. 2022; 14(7):437. https://doi.org/10.3390/toxins14070437
Chicago/Turabian StyleRangel-Muñoz, Erika Janet, Arturo Gerardo Valdivia-Flores, Sanjuana Hernández-Delgado, Carlos Cruz-Vázquez, María Carolina de-Luna-López, Teódulo Quezada-Tristán, Raúl Ortiz-Martínez, and Netzahualcóyotl Mayek-Pérez. 2022. "Assessment of the Potential of a Native Non-Aflatoxigenic Aspergillus flavus Isolate to Reduce Aflatoxin Contamination in Dairy Feed" Toxins 14, no. 7: 437. https://doi.org/10.3390/toxins14070437
APA StyleRangel-Muñoz, E. J., Valdivia-Flores, A. G., Hernández-Delgado, S., Cruz-Vázquez, C., de-Luna-López, M. C., Quezada-Tristán, T., Ortiz-Martínez, R., & Mayek-Pérez, N. (2022). Assessment of the Potential of a Native Non-Aflatoxigenic Aspergillus flavus Isolate to Reduce Aflatoxin Contamination in Dairy Feed. Toxins, 14(7), 437. https://doi.org/10.3390/toxins14070437