The Unusual Metalloprotease-Rich Venom Proteome of the Australian Elapid Snake Hoplocephalus stephensii

 , and
, and 
Abstract
:1. Introduction
2. Results
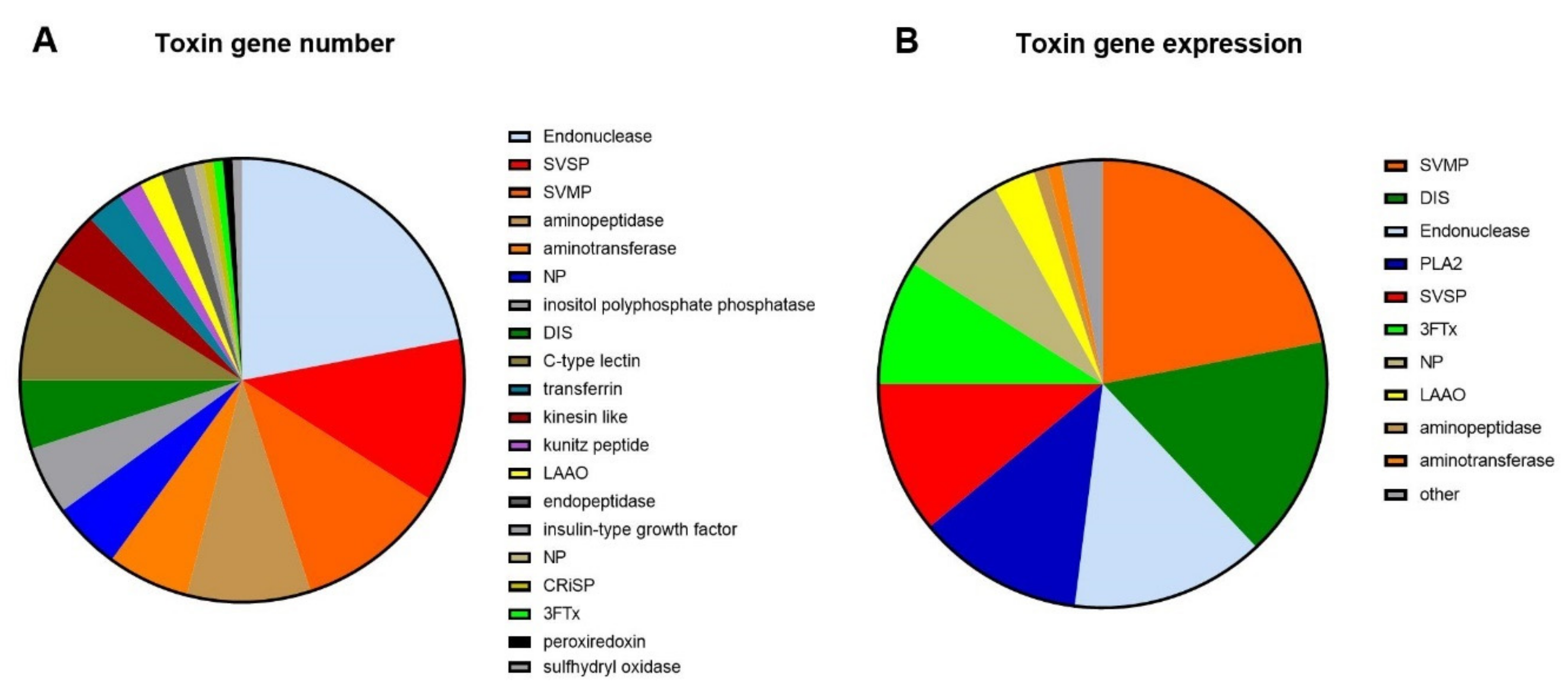
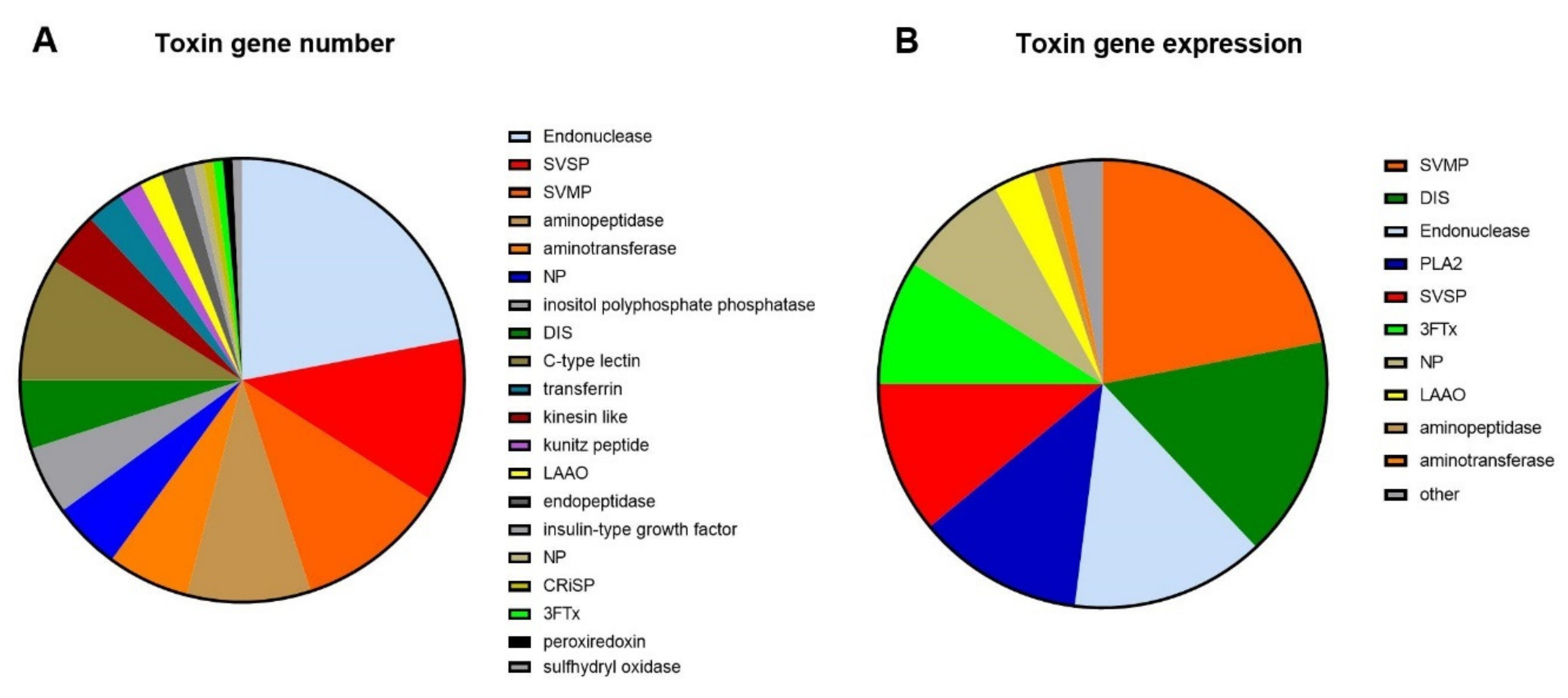
2.1. Transcriptomics
2.2. Proteomics
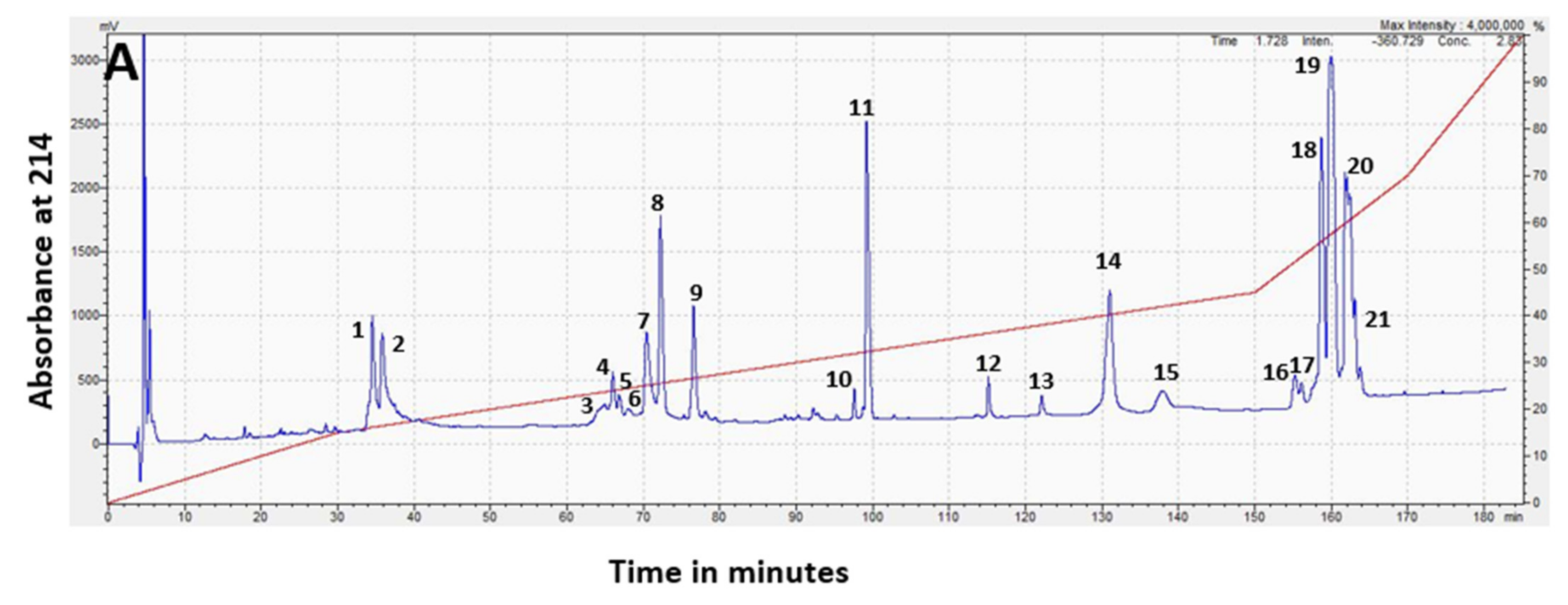
2.2.1. Separation of the whole venom by Reverse-Phase High-Performance Liquid Chromatography (RP-HPLC)
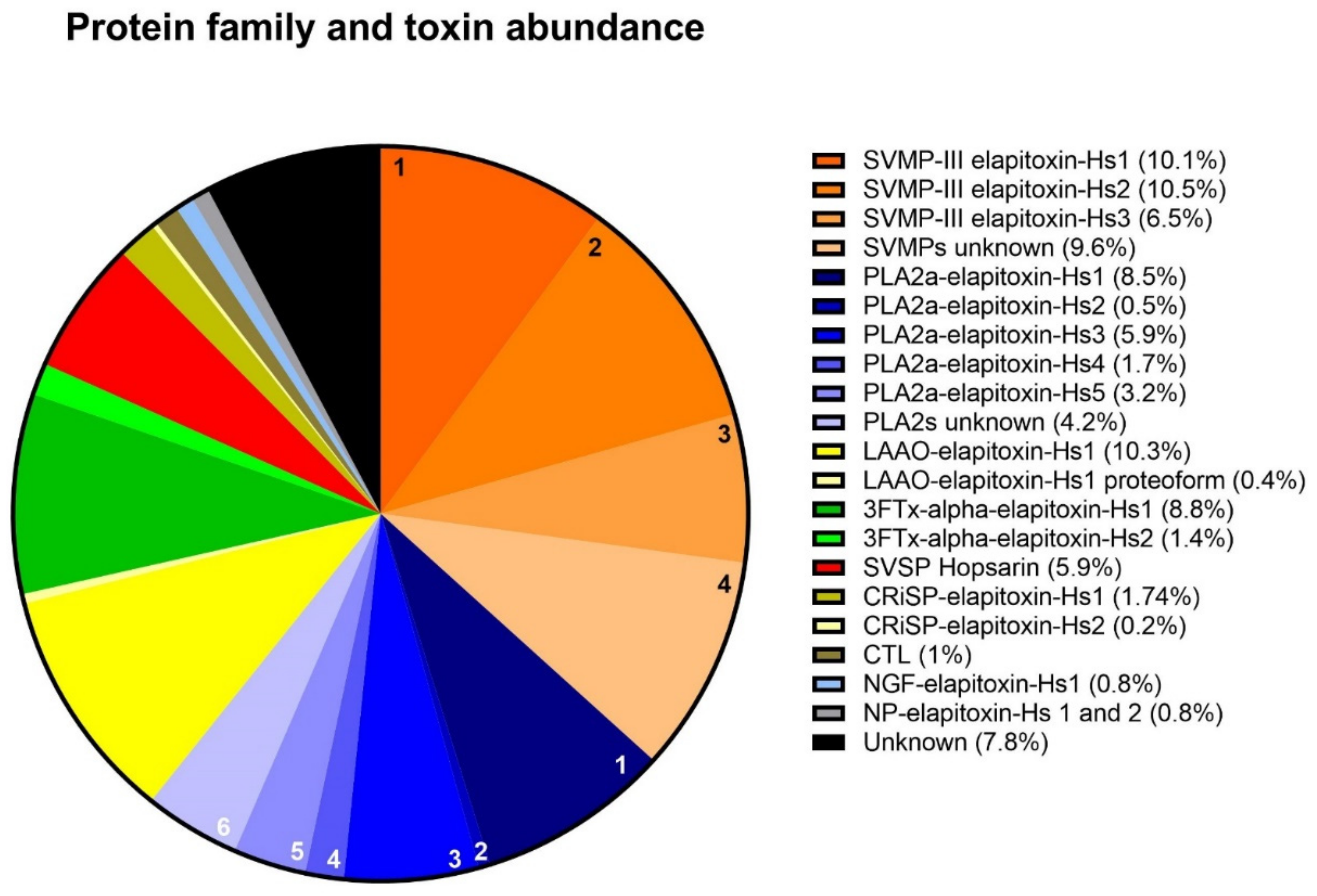
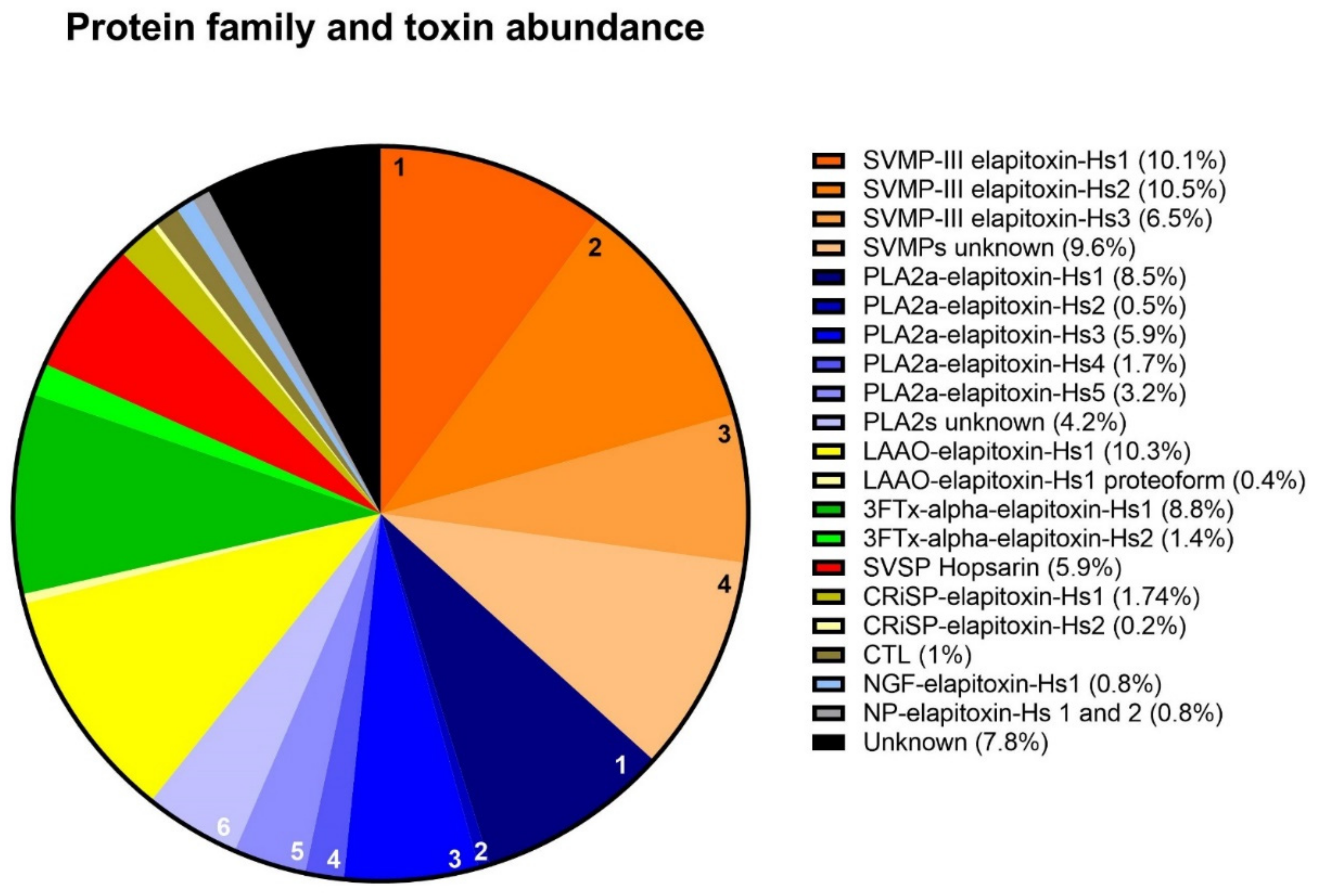
2.2.2. Protein Family Identification and Quantification
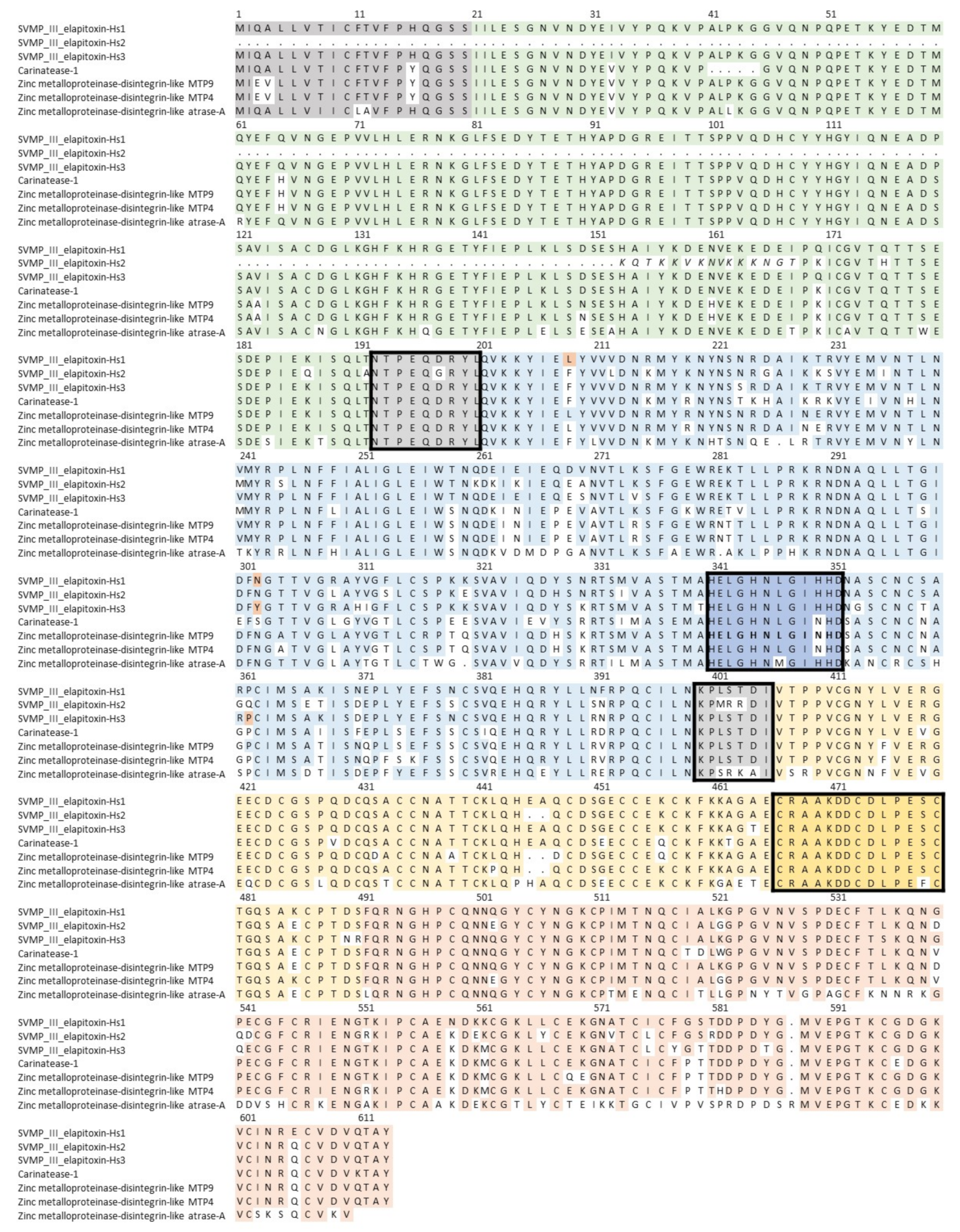
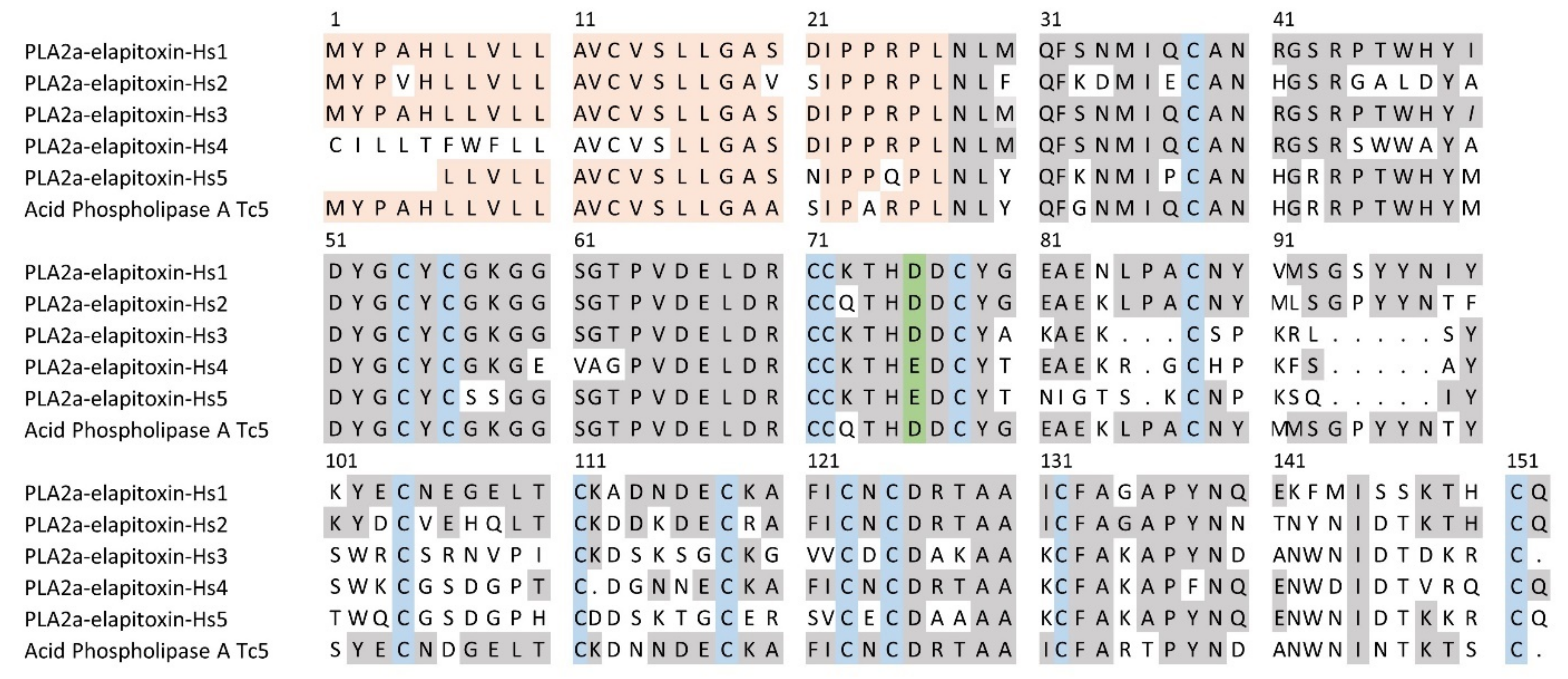
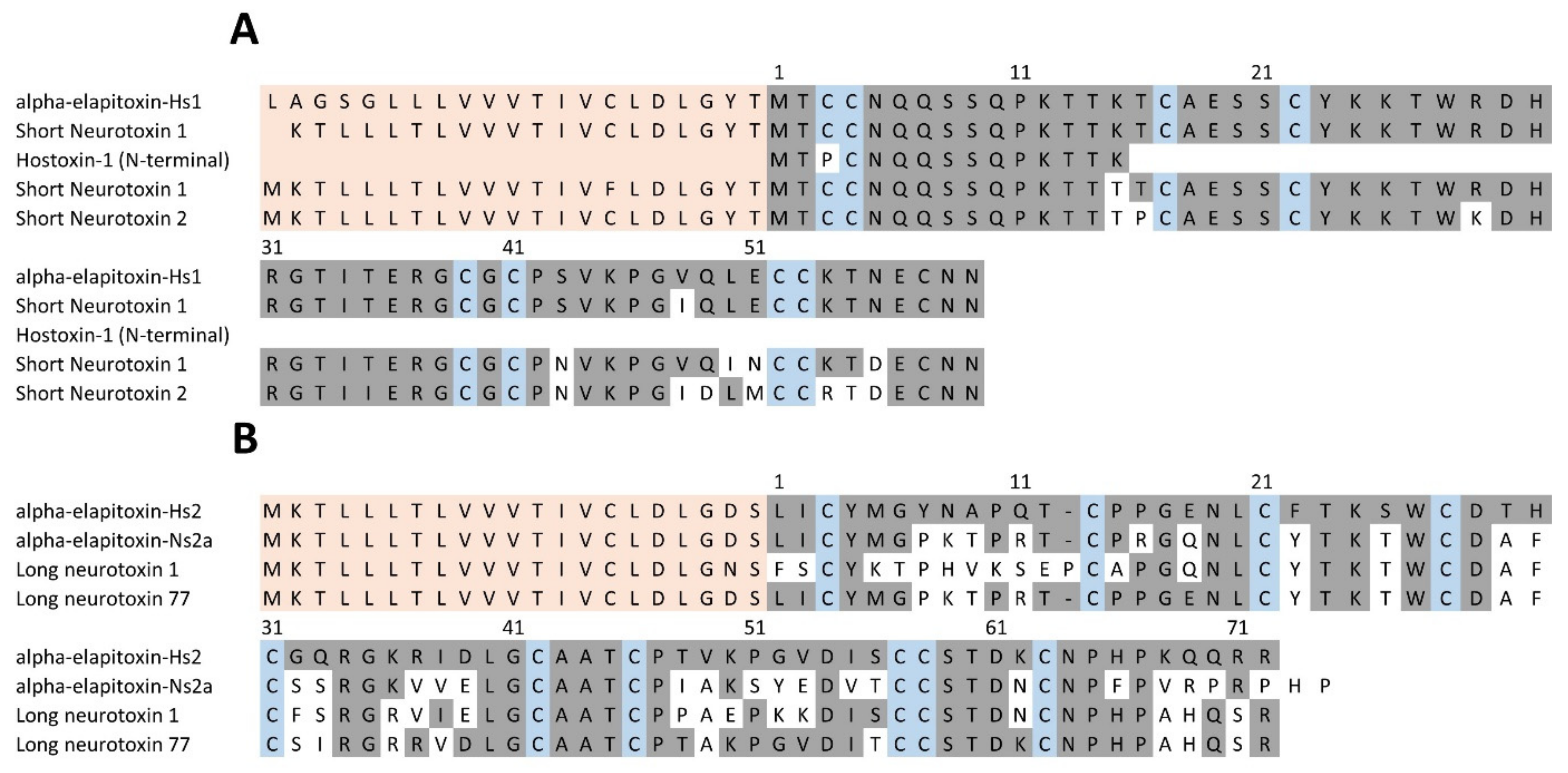
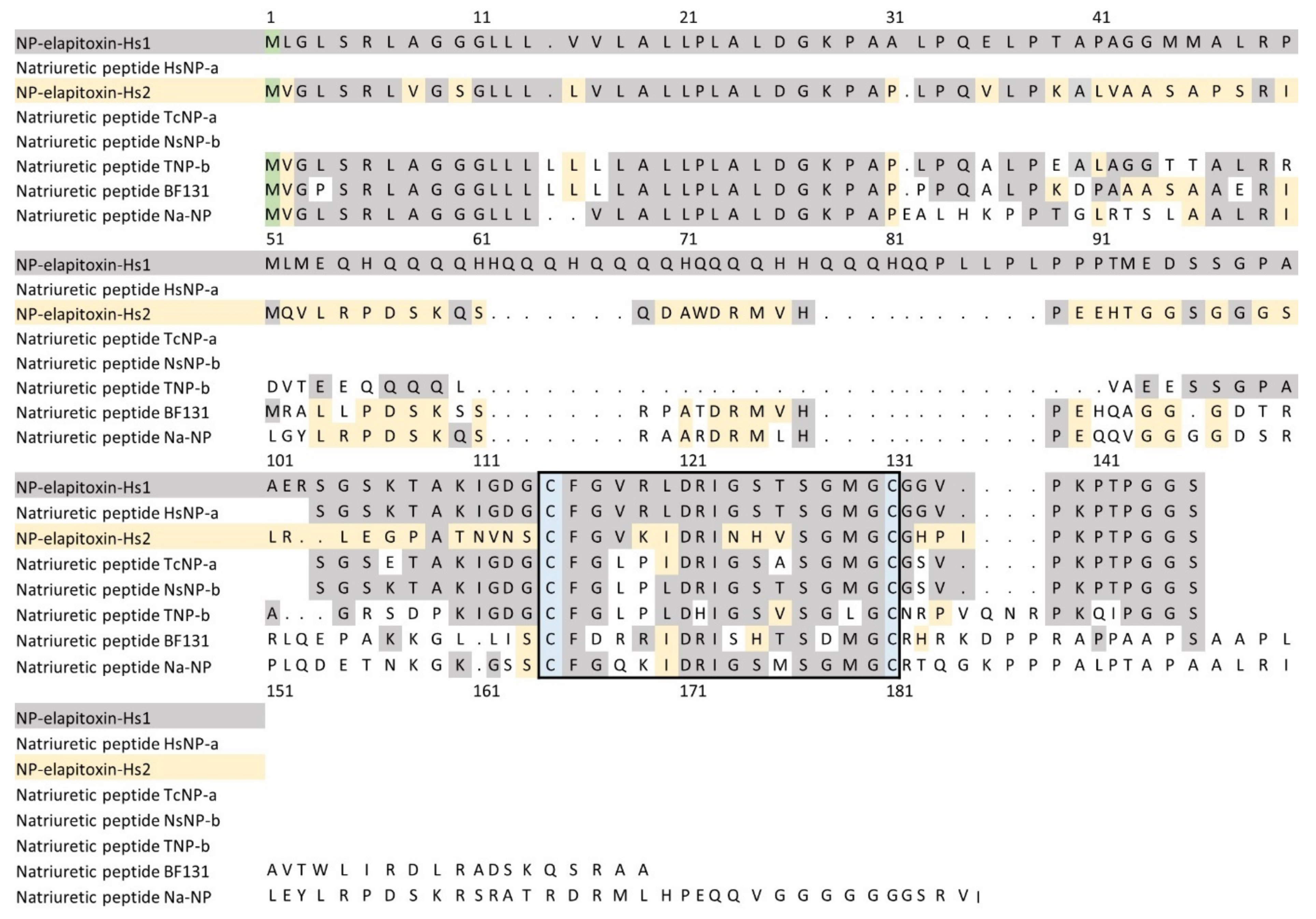
2.2.3. Toxin Identification and Amino Acid Sequences Deduced from the Transcriptome
3. Discussion
4. Methods
4.1. RNA Extraction
4.2. Quality Assessment of Raw Reads
4.3. Transcriptome Assembly
4.4. Assembly Validation
4.5. Differential Gene Expression
4.6. Functional Annotation
4.7. Toxin Identification and Classification
4.8. Comparison of Expression Levels and Gene Counts
4.9. Orthologous Identification
4.10. Mapping Genes to the Cobra Genome Assembly
4.11. Venom
4.12. Reverse-Phase High-Performance Liquid Chromatography
4.13. One Dimensional (1D) SDS-PAGE
4.14. Bottom-up Proteomics
4.14.1. In-Gel Trypsin Digestion
4.14.2. In-Solution Trypsin Digestion
4.14.3. LC–MS/MS Analysis of Venom Fractions
4.14.4. Identification of Toxins Using PEAKS Software and Uniprot
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kelly, C.M.R.; Barker, N.P.; Villet, M.H.; Broadley, N.G. Phylogeny, biogeography and classification of the snake superfamily Elapoidea: A rapid radiation in the late Eocene. Cladistics 2009, 25, 38–63. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, A.; McKelvy, A.D.; Grismer, L.L.; Bell, C.D.; Lailvaux, S.P. A Species-Level Phylogeny of Extant Snakes with Description of a New Colubrid Subfamily and Genus. PLoS ONE 2016, 11, e0161070. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.L.; Lee, M.S.Y.; Leys, R.; Foster, R.; Keogh, J.S. Molecular phylogeny and divergence dates for Australasian elapids and sea snakes (hydrophiinae): Evidence from seven genes for rapid evolutionary radiations. J. Evol. Biol. 2008, 21, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, M.; Shine, R.; Lemckert, F. Radiotelemetric Study of Habitat Use by the Arboreal Snake Hoplocephalus stephensii (Elapidae) in Eastern Australia. Copeia 2002, 2002, 321–332. [Google Scholar] [CrossRef]
- Fitzgerald, M.; Shine, R.; Lemckert, F. Spatial ecology of arboreal snakes (Hoplocephalus stephensii, Elapidae) in an eastern Australian forest. Austral Ecol. 2002, 27, 537–545. [Google Scholar] [CrossRef]
- Fitzgerald, M.; Shine, R.; Lemckert, F. Life history attributes of the threatened Australian snake (Stephen’s banded snake Hoplocephalus stephensii, Elapidae). Biol. Conserv. 2004, 119, 121–128. [Google Scholar] [CrossRef]
- Shine, R. Arboreality in Snakes: Ecology of the Australian Elapid Genus Hoplocephalus. Copeia 1983, 1983, 198. [Google Scholar] [CrossRef]
- Keogh, J.S.; Scott, I.A.W.; Scanlon, J.D. Molecular phylogeny of viviparous Australian elapid snakes: Affinities of Echiopsis atriceps (Storr, 1980) and Drysdalia coronata (Schlegel, 1837), with description of a new genus. J. Zool. 2000, 252, 317–326. [Google Scholar] [CrossRef]
- Fitzgerald, M.; Lazell, B.; Shine, R. Ecology and conservation of the Pale-headed Snake (Hoploceph alus bitorquatus, Elapidae). Aust. Zool. 2010, 35, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Webb, J.; Shine, R. A field study of spatial ecology and movements of a threatened snake species, Hoplocephalus bungaroides. Biol. Conserv. 1997, 82, 203–217. [Google Scholar] [CrossRef]
- Webb, J.K.; Shine, R. Out on a limb: Conservation implications of tree-hollow use by a threatened snake species (Hoplocephalus bungaroides: Serpentes, Elapidae). Biol. Conserv. 1997, 81, 21–33. [Google Scholar] [CrossRef]
- Lister, C.; Arbuckle, K.; Jackson, T.; Debono, J.; Zdenek, C.N.; Dashevsky, D.; Dunstan, N.; Allen, L.; Hay, C.; Bush, B.; et al. Catch a tiger snake by its tail: Differential toxicity, co-factor dependence and antivenom efficacy in a procoagulant clade of Australian venomous snakes. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2017, 202, 39–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodgson, W.C.; Eriksson, C.O.; Alewood, P.F.; Fry, B.G. Comparison of the in vitro neuromuscular activity of venom from three Australian snakes (Hoplocephalus stephensi, Austrelaps superbus and Notechis scutatus): Efficacy of tiger snake antivenom. Clin. Exp. Pharmacol. Physiol. 2003, 30, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Rao, V.S.; Joseph, J.S.; Kini, M. Group D prothrombin activators from snake venom are structural homologues of mammalian blood coagulation factor Xa. Biochem. J. 2003, 369, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Pierre, L.S.; Masci, P.P.; Filippovich, I.; Sorokina, N.; Marsh, N.; Miller, D.J.; Lavin, M.F. Comparative Analysis of Prothrombin Activators from the Venom of Australian Elapids. Mol. Biol. Evol. 2005, 22, 1853–1864. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.C.; Kuruppu, S.; Smith, A.I.; Reeve, S.; Hodgson, W. C Isolation and pharmacological characterisation of hostoxin-1, a postsynaptic neurotoxin from the venom of the Stephen’s banded snake (Hoplocephalus stephensi). Neuropharmacology 2006, 51, 782–788. [Google Scholar] [CrossRef]
- Isbister, G.; White, J.; Currie, B.; O’Leary, M.; Brown, S. Clinical effects and treatment of envenoming by Hoplocephalus spp. snakes in Australia: Australian Snakebite Project (ASP-12). Toxicon 2011, 58, 634–640. [Google Scholar] [CrossRef]
- Noutsos, T.; Currie, B.J.; Isoardi, K.Z.; Brown, S.G.; Isbister, G.K. Snakebite-associated thrombotic microangiopathy: An Australian prospective cohort study [ASP30]. Clin. Toxicol. 2022, 60, 205–213. [Google Scholar] [CrossRef]
- Tasoulis, T.; Isbister, G.K. A Review and Database of Snake Venom Proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef] [Green Version]
- Tasoulis, T.; Pukala, T.L.; Isbister, G.K. Investigating Toxin Diversity and Abundance in Snake Venom Proteomes. Front. Pharmacol. 2022, 12:768015-1-768015-13. [Google Scholar] [CrossRef]
- Tan, C.H.; Tan, K.Y.; Tan, N.H. Revisiting Notechis scutatus venom: On shotgun proteomics and neutralization by the “bivalent” Sea Snake Antivenom. J. Proteom. 2016, 144, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Pierre, L.S.; Fischer, H.; Adams, D.J.; Schenning, M.; Lavidis, N.; De Jersey, J.; Masci, P.P.; Lavin, M.F. Distinct activities of novel neurotoxins from Australian venomous snakes for nicotinic acetylcholine receptors. Cell. Mol. Life Sci. 2007, 64, 2829–2840. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Fung, S.Y.; Yap, M.K.K.; Leong, P.K.; Liew, J.L.; Tan, N.H. Unveiling the elusive and exotic: Venomics of the Malayan blue coral snake (Calliophis bivirgata flaviceps). J. Proteom. 2016, 132, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ziganshin, R.H.; Kovalchuk, S.I.; Arapidi, G.P.; Starkov, V.G.; Hoang, A.N.; Nguyen, T.T.T.; Nguyen, K.C.; Shoibonov, B.B.; Tsetlin, V.I.; Utkin, Y.N. Quantitative proteomic analysis of Vietnamese krait venoms: Neurotoxins are the major components in Bungarus multicinctus and phospholipases A2 in Bungarus fasciatus. Toxicon 2015, 107, 197–209. [Google Scholar] [CrossRef]
- Pla, D.; Bande, B.W.; Welton, R.E.; Paiva, O.K.; Sanz, L.; Segura, A.; Wright, C.E.; Calvete, J.J.; Gutiérrez, J.M.; Williams, D.J. Proteomics and antivenomics of Papuan black snake (Pseudechis papuanus) venom with analysis of its toxicological profile and the preclinical efficacy of Australian antivenoms. J. Proteom. 2017, 150, 201–215. [Google Scholar] [CrossRef]
- Herrera, M.; Fernández, J.; Vargas, M.; Villalta, M.; Segura, Á.; León, G.; Angulo, Y.; Paiva, O.; Matainaho, T.; Jensen, S.D.; et al. Comparative proteomic analysis of the venom of the taipan snake, Oxyuranus scutellatus, from Papua New Guinea and Australia: Role of neurotoxic and procoagulant effects in venom toxicity. J. Proteom. 2012, 75, 2128–2140. [Google Scholar] [CrossRef]
- Wang, C.R.; Bubner, E.R.; Jovcevski, B.; Mittal, P.; Pukala, T.L. Interrogating the higher order structures of snake venom proteins using an integrated mass spectrometric approach. J. Proteom. 2020, 216, 103680. [Google Scholar] [CrossRef]
- Weinstein, S.A.; Williams, V.; White, J. Preliminary characteristics of the prothrombin converting enzyme from venom of Stephen’s banded snake (Hoplocephalus stephensii). Toxicon 2001, 39, 1937–1939. [Google Scholar] [CrossRef]
- Silva, A.; Cristofori-Armstrong, B.; Rash, L.D.; Hodgson, W.C.; Isbister, G.K. Defining the role of post-synaptic α-neurotoxins in paralysis due to snake envenoming in humans. Cell. Mol. Life Sci. 2018, 75, 4465–4478. [Google Scholar] [CrossRef]
- Tan, C.H.; Wong, K.Y.; Chong, H.P.; Tan, N.H.; Tan, K.Y. Proteomic insights into short neurotoxin-driven, highly neurotoxic venom of Philippine cobra (Naja philippinensis) and toxicity correlation of cobra envenomation in Asia. J. Proteom. 2019, 206, 103418. [Google Scholar] [CrossRef]
- Tasoulis, T.; Lee, M.S.Y.; Ziajko, M.; Dunstan, N.; Sumner, J.; Isbister, G.K. Activity of two key toxin groups in Australian elapid venoms show a strong correlation to phylogeny but not to diet. BMC Evol. Biol. 2020, 20, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, J.M.; Lomonte, B. Phospholipases A2: Unveiling the secrets of a functionally versatile group of snake venom toxins. Toxicon 2013, 62, 27–39. [Google Scholar] [CrossRef]
- Cendron, L.; Mičetić, I.; de Laureto, P.P.; Paoli, M. Structural analysis of trimeric phospholipase A2neurotoxin from the Australian taipan snake venom. FEBS J. 2012, 279, 3121–3135. [Google Scholar] [CrossRef] [PubMed]
- Blacklow, B.; Escoubas, P.; Nicholson, G.M. Characterisation of the heterotrimeric presynaptic phospholipase A2 neurotoxin complex from the venom of the common death adder (Acanthophis antarcticus). Biochem. Pharmacol. 2010, 80, 277–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. (Ed.) A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bibsonomy.org/bibtex/f230a919c34360709aa298734d63dca3 (accessed on 6 April 2022).
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerveau, N.; Jackson, D.J. Combining independent de novo assemblies optimizes the coding transcriptome for nonconventional model eukaryotic organisms. BMC Bioinform. 2016, 17, 525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, M.H.; Zerbino, D.R.; Vingron, M.; Birney, E. Oases: Robust de novo RNA-seq assembly across the dynamic range of expression levels. Bioinformatics 2012, 28, 1086–1092. [Google Scholar] [CrossRef]
- Kannan, S.; Hui, J.; Mazooji, K.; Pachter, L.; Tse, D. Shannon: An Information-Optimal de Novo RNA-Seq Assembler. bioRxiv 2016. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Stoeckert, C.J., Jr.; Roos, D.S. OrthoMCL: Identification of Ortholog Groups for Eukaryotic Genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
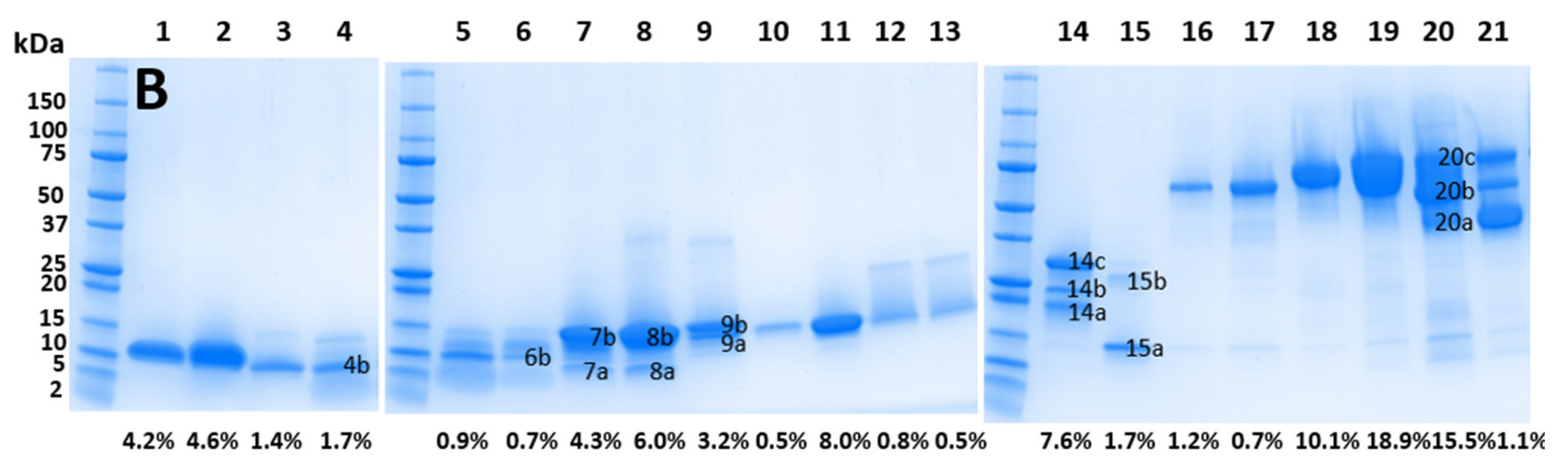
| Peak No # | Band Designation | Protein Family | Peak % of Whole Venom (WV) (Integration) | Band % of Peak (Densitometry) | Toxin % of Band (MS1) | Protein Family % of WV |
|---|---|---|---|---|---|---|
| 1 | 3FTx | 4.2 | 100 | 4.2 | ||
| 2 | 3FTx | 4.6 | 100 | 4.6 | ||
| 3 | 3FTx | 1.4 | 100 | 1.4 | ||
| 4 | 1.7 | |||||
| 4b | PLA2 | 55 | 0.93 | |||
| 5 | 0.9 | |||||
| 5b * | PLA2 | 39 | 0.35 | |||
| 6 | 0.7 | |||||
| 6b | PLA2 | 66.5 | 0.46 | |||
| 7 | 4.3 | |||||
| 7a | 16.5 | 0.7 | ||||
| 7b | PLA2 | 83.5 | 3.59 | |||
| 8 | 6.0 | 6.0 | ||||
| 8a | PLA2 | 15.33 | ||||
| 8b | PLA2 | 84.67 | ||||
| 9 | 3.2 | 3.2 | ||||
| 9a | PLA2 | 46 | ||||
| 9b | PLA2 | 54 | ||||
| 10 | PLA2 | 0.5 | 100 | 0.5 | ||
| 11 | PLA2 | 8.0 | 100 | 8.0 | ||
| 12 | NGF | 0.8 | 74.5 | 69.05 | 0.41 | |
| PLA2 | 30.94 | 0.18 | ||||
| 13 | NGF | 0.5 | 82.9 | 94.74 | 0.39 | |
| 14 | 7.6 | |||||
| 14a | SVSP | 27.7 | 84 | 1.76 | ||
| PLA2 | 5.86 | 0.12 | ||||
| CRiSP | 8.68 | 0.18 | ||||
| 14b | CRiSP | 18 | 1.34 | |||
| 14c | SVSP | 54.3 | 4.13 | |||
| 15 | 1.7 | |||||
| 15a | CTL | 74 | 1.08 | |||
| 15b | CRiSP | 26 | 50.6 | 0.22 | ||
| 16 | SVMP | 1.2 | 100 | 1.2 | ||
| 17 | SVMP | 0.7 | 100 | 0.7 | ||
| 18 | SVMP | 10.1 | 100 | 10.1 | ||
| 19 | LAAO | 18.9 | 100 | 44.14 | 8.34 | |
| SVMP | 55.86 | 10.53 | ||||
| 20 | 15.5 | |||||
| 20a | LAAO | 19.72 | 13.8 | 0.42 | ||
| SVMP | 86.2 | 2.63 | ||||
| 20b | SVMP | 42 | 6.51 | |||
| 20c | LAAO | 38.28 | 33.67 | 1.99 | ||
| SVMP | 66.33 | 3.93 | ||||
| 21 | SVMP | 1.1 | 100 | 1.1 |
| Protein Family/Toxin | % of Whole-Venom Composition | Peaks/Bands |
|---|---|---|
| 3FTx | ||
| alpha-elapitoxin-Hs1 (short-chain neurotoxin) | 8.8 | 1,2 |
| alpha-elapitoxin-Hs2 (long-chain neurotoxin) | 1.4 | 3 |
| PLA2 | ||
| PLA2a-elapitoxin-Hs1 | 8.5 | 10, 11, 12 |
| PLA2a-elapitoxin-Hs2 | 0.5 | 9a |
| PLA2a-elapitoxin-Hs3 | 5.9 | 8b, 9a |
| PLA2a-elapitoxin-Hs4 | 1.7 | 9b |
| PLA2a-elapitoxin-Hs5 | 3.2 | 7 |
| SVMP | ||
| SVMP-III-elapitoxin-Hs1 | 10.1 | 18 |
| SVMP-III-elapitoxin-Hs2 | 10.53 | 19 |
| SVMP-III-elapitoxin-Hs3 | 6.51 | 20b |
| SVSP | ||
| Hopsarin D | 5.9 | 14a, 14c |
| LAAO | ||
| LAAO-elapitoxin-Hs-1 | 10.33 | 19 20c |
| LAAO-elapitoxin-Hs-1 proteoform | 0.42 | 20a |
| CRiSP | ||
| CRiSP-elapitoxin-Hs-1 | 1.52 | 14a,14b |
| CRiSP-elapitoxin-Hs-2 | 0.22 | 15b |
| CTL | ||
| unsequenced | 1.08 | 15a |
| NGF | ||
| NGF-elapitoxin-Hs1 | 0.8 | 12,13 |
| NP | 0.79 | 4, 5, 6 |
| NP-elapitoxin-Hs1 | ||
| NP-elapitoxin-Hs2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tasoulis, T.; Wang, C.R.; Sumner, J.; Dunstan, N.; Pukala, T.L.; Isbister, G.K. The Unusual Metalloprotease-Rich Venom Proteome of the Australian Elapid Snake Hoplocephalus stephensii. Toxins 2022, 14, 314. https://doi.org/10.3390/toxins14050314
Tasoulis T, Wang CR, Sumner J, Dunstan N, Pukala TL, Isbister GK. The Unusual Metalloprotease-Rich Venom Proteome of the Australian Elapid Snake Hoplocephalus stephensii. Toxins. 2022; 14(5):314. https://doi.org/10.3390/toxins14050314
Chicago/Turabian StyleTasoulis, Theo, C. Ruth Wang, Joanna Sumner, Nathan Dunstan, Tara L. Pukala, and Geoffrey K. Isbister. 2022. "The Unusual Metalloprotease-Rich Venom Proteome of the Australian Elapid Snake Hoplocephalus stephensii" Toxins 14, no. 5: 314. https://doi.org/10.3390/toxins14050314
APA StyleTasoulis, T., Wang, C. R., Sumner, J., Dunstan, N., Pukala, T. L., & Isbister, G. K. (2022). The Unusual Metalloprotease-Rich Venom Proteome of the Australian Elapid Snake Hoplocephalus stephensii. Toxins, 14(5), 314. https://doi.org/10.3390/toxins14050314