Detection of Enterotoxigenic Psychrotrophic Presumptive Bacillus cereus and Cereulide Producers in Food Products and Ingredients



Abstract
:1. Introduction
2. Results and Discussion
2.1. Distribution of Toxin Genes among Presumptive B. cereus Organisms Obtained from Food Products
2.2. Growth of Toxigenic Presumptive B. cereus at 4, 7, and 10 °C
2.3. Toxigenic Profiles of Psychrotrophic Presumptive B. cereus Grown at 4 °C
2.4. Cereulide Production by Psychrotropic Presumptive B. cereus at 37 °C and 4 °C
3. Conclusions
4. Materials and Methods
4.1. Collection of Food Products
4.2. Detection of Presumptive B. cereus
4.3. Toxin Gene Profiling
4.4. Refrigeration and Mild Abuse Temperature Exposure
4.5. Emetic Toxin Detection via Boar Semen Motility Assay
4.6. Quantification of Cereulide Production at 4 °C by LC–MS² Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). Risks for Public Health Related to the Presence of Bacillus cereus and other Bacillus spp. including Bacillus thuringiensis in Foodstuffs. EFSA J. 2016, 14, e04524. [Google Scholar] [CrossRef]
- Jung, M.Y.; Paek, W.K.; Park, I.S.; Han, J.R.; Sin, Y.; Paek, J.; Rhee, M.S.; Kim, H.; Song, H.S.; Chang, Y.H. Bacillus gaemokensis sp. nov., Isolated from Foreshore Tidal Flat Sediment from the Yellow Sea. J. Microbiol. 2010, 48, 867–871. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.Y.; Kim, J.S.; Paek, W.K.; Lim, J.; Lee, H.; Kim, P., II; Ma, J.Y.; Kim, W.; Chang, Y.H. Bacillus manliponensis sp. nov., a New Member of the Bacillus cereus Group Isolated from Foreshore Tidal Flat Sediment. J. Microbiol. 2011, 49, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, G.H.; Hu, G.P.; Cetin, S.; Lin, N.Q.; Tang, J.Y.; Tang, W.Q.; Lin, Y.Z. Bacillus bingmayongensis sp. nov., Isolated from the Pit soil of Emperor Qin’s Terra-cotta Warriors in China. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2014, 105, 501–510. [Google Scholar] [CrossRef]
- Miller, R.A.; Beno, S.M.; Kent, D.J.; Carroll, L.M.; Martin, N.H.; Boor, K.J.; Kovac, J. Bacillus wiedmannii sp. nov., a Psychrotolerant and Cytotoxic Bacillus cereus group species Isolated from Dairy Foods and Dairy Environments. Int. J. Syst. Evol. Microbiol. 2016, 66, 4744–4753. [Google Scholar] [CrossRef]
- Liu, Y.; Du, J.; Lai, Q.; Zeng, R.; Ye, D.; Xu, J.; Shao, Z. Proposal of Nine Novel Species of the Bacillus cereus Group. Int. J. Syst. Evol. Microbiol. 2017, 67, 2499–2508. [Google Scholar] [CrossRef]
- Guo, J.; Wang, Y.; Yang, G.; Chen, Y.; Zhou, S.; Zhao, Y.; Zhuang, L. Bacillus Nitroreducens sp. nov., a Humus-Reducing Bacterium Isolated from a Compost. Arch. Microbiol. 2016, 198, 347–352. [Google Scholar] [CrossRef]
- Helgason, E.; Økstad, O.L.E.A.; Caugant, D.A.; Mock, L.E.; Hegna, I.D.; Johansen, H.A.; Fouet, A.; Mock, M.; Hegna, I.; Kolstø, A.B. One Species on the Basis of Genetic Evidence. Appl. Environ. Microbiol. 2000, 66, 2627–2630. [Google Scholar] [CrossRef] [Green Version]
- Guinebretière, M.H.; Velge, P.; Couvert, O.; Carlin, F.; Debuyser, M.L.; Nguyen-The, C. Ability of Bacillus cereus group strains to cause Food Poisoning Varies According to Phylogenetic Affiliation (groups I to VII) Rather Than Species Affiliation. J. Clin. Microbiol. 2010, 48, 3388–3391. [Google Scholar] [CrossRef] [Green Version]
- Jovanovic, J.; Ornelis, V.F.M.; Madder, A.; Rajkovic, A. Bacillus cereus Food Intoxication and Toxicoinfection. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3719–3761. [Google Scholar] [CrossRef]
- Berthold-Pluta, A.; Pluta, A.; Garbowska, M. The Effect of Selected Factors on the Survival of Bacillus cereus in the Human Gastrointestinal Tract. Microb. Pathog. 2015, 82, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Lund, T.; de Buyser, M.; Einar, P.; Aliments, Â.S. A New Cytotoxin from Bacillus cereus That May Cause Necrotic Enteritis. Mol. Microbiol. 2000, 38, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Senesi, S.; Ghelardi, E. Production, Secretion and Biological activity of Bacillus cereus enterotoxins. Toxins 2010, 2, 1690–1703. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Frenzel, E.; Gohar, M. Food-bacteria interplay: Pathometabolism of Emetic Bacillus cereus. Front. Microbiol. 2015, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkovic, A.; Uyttendaele, M.; Dierick, K.; Samapundo, S.; Botteldoorn, N.; Mahillon, J.; Heyndrickx, M. Risk Profile of the Bacillus cereus Group Implicated in Food Poisoning. Rep. Super. Health Counc. Belg. 2008, 1, 1–80. [Google Scholar]
- Mahler, H.; Pasi, A.; Kramer, J.M.; Schulte, P.; Scoging, A.C.; Bär, W.; Krähenbühl, S. Fulminant Liver Failure in Association with the Emetic Toxin of Bacillus cereus. N. Engl. J. Med. 1997, 336, 1142–1148. [Google Scholar] [CrossRef]
- Vangoitsenhoven, R.; Rondas, D.; Crèvecoeur, I.; D’Hertog, W.; Baatsen, P.; Masini, M.; Van der Schueren, B. Foodborne Cereulide Causes beta-cell Dysfunction and Apoptosis. PLoS ONE 2014, 9, e104866. [Google Scholar] [CrossRef]
- Dierick, K.; van Coillie, E.; Swiecicka, I.; Meyfroidt, G.; Devlieger, H.; Meulemans, A.; Hoedemaekers, G.; Fourie, L.; Heyndrickx, M.; Mahillon, J. Fatal Family Outbreak of Bacillus cereus. Assoc. Food Poisoning 2005, 43, 4277–4279. [Google Scholar]
- Bauer, T.; Sipos, W.; Stark, T.D.; Käser, T.; Knecht, C.; Brunthaler, R.; Saalmüller, A.; Hofmann, T.; Ehling-Schulz, M. First Insights Into within Host Translocation of the Bacillus cereus Toxin Cereulide Using a Porcine Model. Front. Microbiol. 2018, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Naranjo, M.; Denayer, S.; Botteldoorn, N.; Delbrassinne, L.; Veys, J.; Waegenaere, J.; Sirtaine, N.; Driesen, R.B.; Sipido, K.R.; Mahillon, J.; et al. Sudden Death of a Young Adult Associated with Bacillus cereus Food Poisoning. J. Clin. Microbiol. 2011, 49, 4379–4381. [Google Scholar] [CrossRef] [Green Version]
- Shiota, M.; Saitou, K.; Mizumoto, H.; Matsusaka, M.; Agata, N.; Nakayama, M.; Kage, M.; Tatsumi, S.; Okamoto, A.; Yamaguchi, S.; et al. Rapid Detoxification of Cereulide in Bacillus cereus Food Poisoning. Pediatrics 2010, 125, e951–e955. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Agency (EFSA). Opinion of the Scientific Panel on Biological Hazards on Bacillus cereus and Other Bacillus spp. in Foodstuffs. EFSA J. 2005, 175, 1–48. [Google Scholar]
- Schreiber, N.; Hackl, G.; Reisinger, A.C.; Zollner-schwetz, I.; Eller, K.; Schlagenhaufen, C.; Pietzka, A.; Czerwenka, C.; Stark, T.D.; Kranzler, M.; et al. Acute Liver Failure after Ingestion of Fried Rice Balls: A Case Series of Bacillus cereus Food Poisonings. Toxins 2022, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne outbreaks in 2015. EFSA J. 2016, 14, e20449. [Google Scholar]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in 2014. EFSA J. 2015, 13, 4329. [Google Scholar]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in in 2016. EFSA J. 2017, 15, 5077. [Google Scholar]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union One Health 2018 Zoonoses report. EFSA J. 2019, 17, e05926. [Google Scholar]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union One Health 2019 Zoonoses Report 2020. EFSA J. 2021, 19, 6406. [Google Scholar]
- Webb, M.D.; Barker, G.C.; Goodburn, K.E.; Peck, M.W. Risk Presented to Minimally Processed Chilled Foods by Psychrotrophic Bacillus cereus. Trends Food Sci. Technol. 2019, 93, 94–105. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in 2010. EFSA J. 2012, 17, 2597. [Google Scholar]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in 2017. EFSA J. 2018, 16, e05500. [Google Scholar]
- James, C.; Onarinde, B.A.; James, S.J. The Use and Performance of Household Refrigerators: A Review. Compr. Rev. Food Sci. Food Saf. 2017, 16, 160–179. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic, J.; Djekic, I.; Smigic, N.; Tomic, N.; Rajkovic, A. Temperature Profile and Hygiene in Household Refrigerators in Belgrade, Serbia and Their Relation to Consumers Food Safety Knowledge and Characteristics of the Refrigerators. Food Control 2022, 136, 108813. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in 2012. EFSA J. 2014, 12, 3547. [Google Scholar]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). Analysis of the Baseline Survey on the Prevalence of Listeria monocytogenes in Certain Ready-to-Eat Foods in the EU, 2010–2011 Part A: Listeria Monocytogenes Prevalence Estimates. EFSA J. 2013, 11, 3241. [Google Scholar] [CrossRef]
- Forghani, F.; Kim, J.B.; Oh, D.H. Enterotoxigenic Profiling of Emetic Toxin- and Enterotoxin-Producing Bacillus cereus, isolated from Food, Environmental, and Clinical Samples by Multiplex PCR. J. Food Sci. 2014, 79, M2288–M2293. [Google Scholar] [CrossRef]
- Wijnands, L.M.; Dufrenne, J.B.; Rombouts, F.M.; in’t Veld, P.H.; van Leusden, F.M. Prevalence of Potentially Pathogenic Bacillus cereus in Food Commodities in The Netherlands. J. Food Prot. 2006, 69, 2587–2594. [Google Scholar] [CrossRef]
- Berthold-pluta, A.; Pluta, A.; Garbowska, M. Prevalence and Toxicity Characterization of Bacillus cereus in Food Products from Poland. Foods 2019, 8, 269. [Google Scholar] [CrossRef] [Green Version]
- Biesta-Pieters, E.G.; Dissel, S.; Reij, M.W.; Zwietering, M.H.; In’t Veld, P.H. Characterization and Exposure Assessment of Emetic Bacillus cereus and Cereulide Production in Food Products on the Dutch Market. J. Food Prot. 2016, 79, 230–238. [Google Scholar] [CrossRef]
- Böhm, M.; Huptas, C.; Krey, V.M.; Scherer, S. Massive Horizontal Gene Transfer, Strictly Vertical Inheritance and Ancient Duplications Differentially Shape the Evolution of Bacillus cereus Enterotoxin Operons hbl, cytK and nhe. BMC Evol. Biol. 2015, 15, 246. [Google Scholar] [CrossRef] [Green Version]
- Fiedler, G.; Schneider, C.; Igbinosa, E.O.; Kabisch, J.; Brinks, E.; Becker, B.; Stoll, D.A.; Cho, G.S.; Huch, M.; Franz, C.M.A.P. Antibiotics Resistance and Toxin Profiles of Bacillus cereus-Group Isolates from Fresh Vegetables from German Retail Markets. BMC Microbiol. 2019, 19, 250. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Bang, J.; Kim, H.; Kim, Y.; Kim, B.; Beuchat, L.R.; Ryu, J. Bacillus cereus and Bacillus thuringiensis Spores in Korean Rice: Prevalence and Toxin Production as Affected by Production Area and Degree of Milling. Food Microbiol. 2014, 42, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Glasset, B.; Herbin, S.; Guillier, L.; Cadel-Six, S.; Vignaud, M.; Grout, J.; Pairaud, S.; Michel, V.; Hennekinne, J.; Ramarao, N.; et al. Bacillus cereus-Induced Food-Borne Outbreaks in France, 2007 to 2014: Epidemiology and Genetic Characterisation. Eurosurveillance 2016, 21, 30413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burtscher, J.; Etter, D.; Biggel, M.; Schlaepfer, J.; Johler, S. Further Insights Into the Toxicity of Bacillus cytotoxicus Based on Toxin Gene Profiling and Vero Cell Cytotoxicity Assays. Toxins 2021, 13, 234. [Google Scholar] [CrossRef] [PubMed]
- Cairo, J.; Gherman, I.; Day, A.; Cook, P.E. Bacillus cytotoxicus—A Potentially Virulent Food-Associated Microbe. J. Appl. Microbiol. 2022, 132, 31–40. [Google Scholar] [CrossRef]
- Gamage, N.W.; Bamforth, J.; Ashfaq, T.; Bernard, K.; Gräfenhan, T.; Walkowiak, S. Profiling of Bacillus cereus on Canadian Grain. PLoS ONE 2021, 16, e0259209. [Google Scholar] [CrossRef]
- Samapundo, S.; Heyndrickx, M.; Xhaferi, R.; Devlieghere, F. Incidence, Diversity and Toxin Gene Characteristics of Bacillus cereus Group Strains Isolated from Food Products Marketed in Belgium. Int. J. Food Microbiol. 2011, 150, 34–41. [Google Scholar] [CrossRef]
- Ankolekar, C.; Rahmati, T.; Labbé, R.G. Detection of Toxigenic Bacillus cereus and Bacillus thuringiensis Spores in U.S. Rice. Int. J. Food Microbiol. 2009, 128, 460–466. [Google Scholar] [CrossRef]
- Koné, K.M.; Douamba, Z.; De Halleux, M.; Bougoudogo, F.; Mahillon, J. Prevalence and Diversity of the Thermotolerant Bacterium Bacillus cytotoxicus Among Dried Food Products. J. Food Prot. 2019, 82, 1210–1216. [Google Scholar] [CrossRef]
- Yu, S.; Yu, P.; Wang, J.; Li, C.; Guo, H.; Liu, C.; Kong, L.; Yu, L.; Wu, S.; Lei, T.; et al. A Study on Prevalence and Characterization of Bacillus cereus in Ready-to-Eat Foods in China. Front. Microbiol. 2020, 10, 3043. [Google Scholar] [CrossRef] [Green Version]
- Rana, N.; Panda, A.K.; Pathak, N.; Gupta, T.; Thakur, S.D. Bacillus cereus: Public Health Burden Associated with Ready-to-Eat Foods in Himachal Pradesh, India. J. Food Sci. Technol. 2020, 57, 2293–2302. [Google Scholar] [CrossRef] [PubMed]
- Dufrenne, J.; Tatini, S.; Notermans, S. Stability of Spores of Bacillus cereus Stored on Silicagel. Int. J. Food Microbiol. 1994, 23, 111–116. [Google Scholar] [CrossRef]
- Messelhäusser, U.; Frenzel, E.; Blöchinger, C.; Zucker, R.; Kämpf, P.; Ehling-Schulz, M. Emetic Bacillus cereus are More Volatile than Thought: Recent Foodborne Outbreaks and Prevalence Studies in Bavaria (2007–2013). BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, H.; Schaller, G.; von Wiese, W.; Terplan, G. Bacillus cereus in Infant Foods and Dried Milk Products. Int. J. Food Microbiol. 1994, 23, 1–15. [Google Scholar] [CrossRef]
- Tatsinkou Fossi, B.; Tatah Kihla Akoachere, J.F.; Nchanji, G.T.; Wanji, S. Occurrence, Heat and Antibiotic Resistance Profile of Bacillus cereus Isolated from Raw Cow and Processed Milk in Mezam Division, Cameroon. Int. J. Dairy Technol. 2017, 70, 43–51. [Google Scholar] [CrossRef]
- Pei, X.; Yang, S.; Zhan, L.; Zhu, J.; Song, X.; Hu, X.; Liu, G.; Ma, G.; Li, N.; Yang, D. Prevalence of Bacillus cereus in Powdered Infant and Powdered Follow-Up Formula in China. Food Control 2018, 93, 101–105. [Google Scholar] [CrossRef]
- Liu, X.Y.; Hu, Q.; Xu, F.; Ding, S.Y.; Zhu, K. Characterization of Bacillus cereus in Dairy Products in China. Toxins 2020, 12, 454. [Google Scholar] [CrossRef]
- Heini, N.; Stephan, R.; Ehling-Schulz, M.; Johler, S. Characterization of Bacillus cereus Group Isolates from Powdered Food Products. Int. J. Food Microbiol. 2018, 283, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Park, K.M.; Kim, H.J. Biofilm Formation of Low-Temperature-Tolerant Bacillus cereus Isolated from Green Leaf Lettuce in the Cold Chain. Foods 2020, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Choma, C.; Guinebretière, M.H.; Carlin, F.; Schmitt, P.; Velge, P.; Granum, P.E.; Nguyen-The, C. Prevalence, Characterization and Growth of Bacillus cereus in Commercial Cooked Chilled Foods Containing Vegetables. J. Appl. Microbiol. 2000, 88, 617–625. [Google Scholar] [CrossRef]
- Carlin, F.; Fricker, M.; Pielaat, A.; Heisterkamp, S.; Shaheen, R.; Salkinoja Salonen, M.; Svensson, B.; Nguyen-the, C.; Ehling-Schulz, M. Emetic Toxin-Producing Strains of Bacillus cereus Show Distinct Characteristics within the Bacillus cereus Group. Int. J. Food Microbiol. 2006, 109, 132–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okanlawon, B.M.; Ogunbanwo, S.T.; Okunlola, A.O. Growth of Bacillus cereus Isolated from Some Traditional Condiments Under Different Regimens. Afr. J. Biotechnol. 2010, 9, 2129–2135. [Google Scholar]
- Jan, S.; Brunet, N.; Techer, C.; Le Maréchal, C.; Koné, A.Z.; Grosset, N.; Cochet, M.F.; Gillard, A.; Gautier, M.; Puterflam, J.; et al. Biodiversity of Psychrotrophic Bacteria of the Bacillus cereus Group Collected in Farm and Egg Product Industry. Food Microbiol. 2011, 28, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Carroll, L.M.; Cheng, R.A.; Wiedmann, M.; Kovac, J. Keeping Up with the Bacillus cereus Group: Taxonomy Through the Genomics Era and Beyond. Crit. Rev. Food Sci. Nutr. 2021, 1–26. [Google Scholar] [CrossRef]
- Merzougui, S.; Cohen, N.; Grosset, N.; Gautier, M.; Lkhider, M. Enterotoxigenic Profiles of Psychrotolerant and Mesophilic Strains of the Bacillus cereus Group Isolated from Food in Morocco. Int. J. Eng. Res. Appl. 2013, 3, 964–970. [Google Scholar]
- Stenfors, L.P.; Granum, P.E. Psychrotolerant Species from the Bacillus cereus Group are not Necessarily Bacillus weihenstephanensis. FEMS Microbiol. Lett. 2001, 197, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Godič Torkar, K.; Seme, K. Antimicrobial Susceptibility, β-Lactamase and Enterotoxin Production in Bacillus cereus Isolates from Clinical and Food Samples. Folia Microbiol. 2009, 54, 233–238. [Google Scholar] [CrossRef]
- Montanhini, M.T.M.; dos Santos Bersot, L. Avaliação do Comportamento Psicrotrófico e Atividade Lipolítica e Proteolítica de Bacillus cereus Isolado de Produtos Lácteos Refrigerados. Acta Sci. Technol. 2013, 35, 163–167. [Google Scholar]
- Fiedoruk, K.; Drewnowska, J.M.; Daniluk, T.; Leszczynska, K.; Iwaniuk, P.; Swiecicka, I. Ribosomal Background of the Bacillus cereus Group Thermotypes. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Bartoszewicz, M.; Bideshi, D.K.; Kraszewska, A.; Modzelewska, E.; Swiecicka, I. Natural Isolates of Bacillus thuringiensis Display Genetic and Psychrotrophic Properties Characteristic of Bacillus weihenstephanensis. J. Appl. Microbiol. 2009, 106, 1967–1975. [Google Scholar] [CrossRef]
- Thorsen, L.; Budde, B.B.; Henrichsen, L.; Martinussen, T.; Jakobsen, M. Cereulide Formation by Bacillus Weihenstephanensis and Mesophilic Emetic Bacillus cereus at Temperature Abuse Depends on Pre-Incubation conditions. Int. J. Food Microbiol. 2009, 134, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Ellouze, M.; Buss Da Silva, N.; Rouzeau-Szynalski, K.; Coisne, L.; Cantergiani, F.; Baranyi, J. Modeling Bacillus cereus Growth and Cereulide Formation in Cereal-, Dairy-, Meat-, Vegetable-Based Food and Culture Medium. Front. Microbiol. 2021, 12, 155. [Google Scholar]
- Finlay, W.J.J.; Logan, N.A.; Sutherland, A.D. Bacillus cereus Produces Most Emetic toxin at Lower Temperatures. Lett. Appl. Microbiol. 2000, 31, 385–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kranzler, M.; Stollewerk, K.; Rouzeau-szynalski, K.; Blayo, L.; Sulyok, M.; Ehling-Schulz, M. Temperature Exerts Control of Bacillus cereus Emetic Toxin Production on Post-Transcriptional Levels. Front. Microbiol. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marxen, S.; Stark, T.D.; Frenzel, E.; Rütschle, A.; Lücking, G.; Pürstinger, G.; Pohl, E.E.; Scherer, S.; Ehling-Schulz, M.; Hofmann, T. Chemodiversity of Cereulide, the Emetic Toxin of Bacillus cereus. Anal. Bioanal. Chem. 2015, 407, 2439–2453. [Google Scholar] [CrossRef]
- European Food Safety Agency (EFSA). Manual for Reporting on Food-Borne Outbreaks in Accordance with Directive 2003/99/EC for Information Deriving from the Year 2016. EFSA Support. Publ. 2017, 14, 1174E. [Google Scholar] [CrossRef] [Green Version]
- Yang, I.C.; Shih, D.Y.C.; Huang, T.P.; Huang, Y.P.; Wang, J.Y.; Pan, T.M. Establishment of a Novel Multiplex PCR Assay and Detection of Toxigenic Strains of the Species in the Bacillus cereus Group. J. Food Prot. 2005, 68, 2123–2130. [Google Scholar] [CrossRef]
- Wehrle, E.; Didier, A.; Moravek, M.; Dietrich, R.; Märtlbauer, E. Detection of Bacillus cereus with Enteropathogenic Potential by Multiplex Real-Time PCR-Based on SYBR Green I. Mol. Cell. Probes 2010, 24, 124–130. [Google Scholar] [CrossRef]
- Fricker, M.; Messelhäußer, U.; Busch, U.; Scherer, S.; Ehling-Schulz, M. Diagnostic Real-Time PCR Assays for the Detection of Emetic Bacillus cereus Strains in Foods and Recent Food-Borne Outbreaks. Appl. Environ. Microbiol. 2007, 73, 1892–1898. [Google Scholar] [CrossRef] [Green Version]
- Rajkovic, A.; Uyttendaele, M.; Debevere, J. Computer Aided Boar Semen Motility Analysis for Cereulide Detection in Different Food Matrices. Int. J. Food. Microl. 2007, 114, 92–99. [Google Scholar] [CrossRef]
- Delbrassinne, L.; Andjelkovic, M.; Rajkovic, A.; Dubois, P.; Nguessan, E.; Mahillon, J.; van Loco, J. Determination of Bacillus cereus Emetic Toxin in Food Products by Means of LC-MS2. Food Anal. Methods 2012, 5, 969–979. [Google Scholar] [CrossRef]
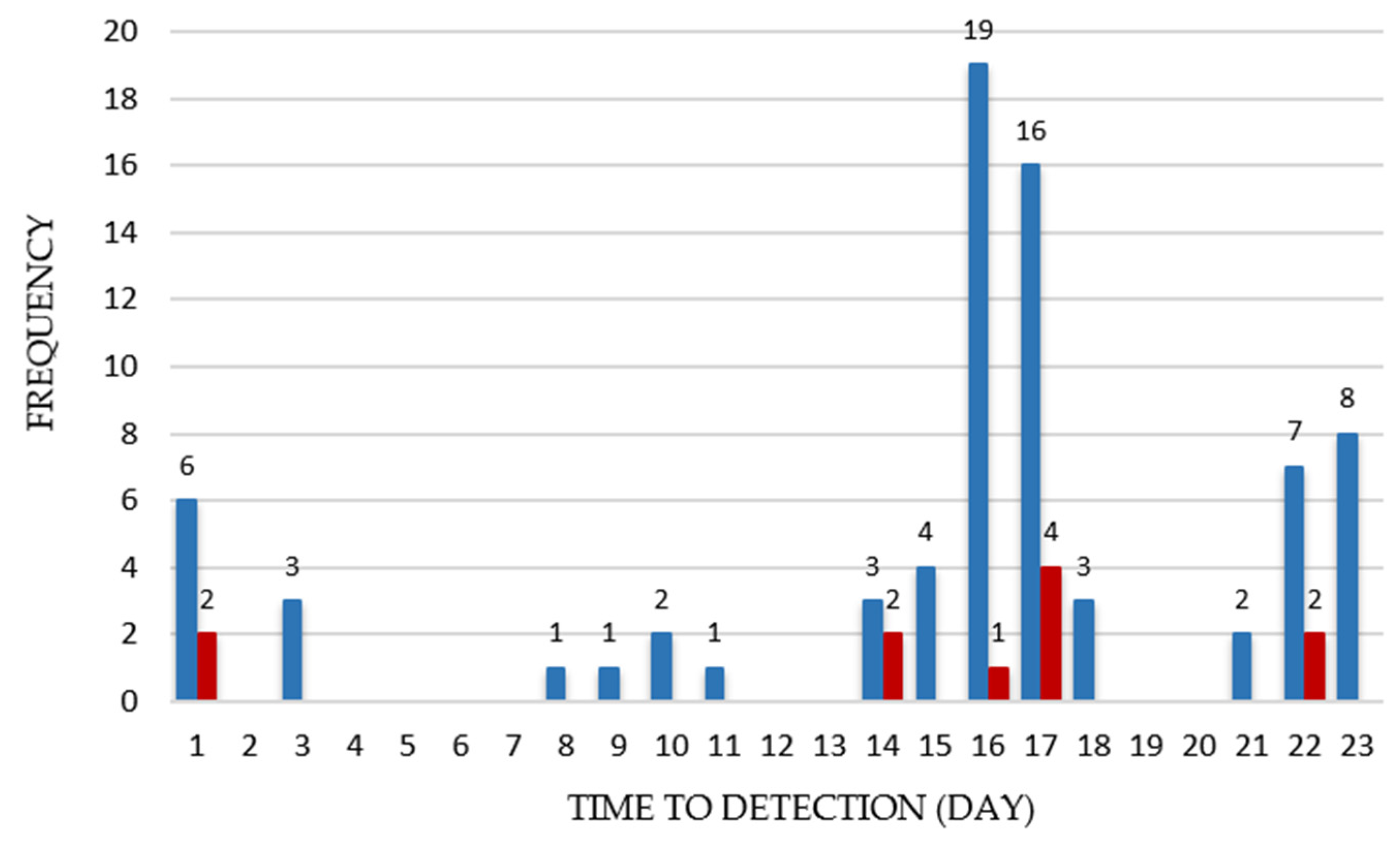

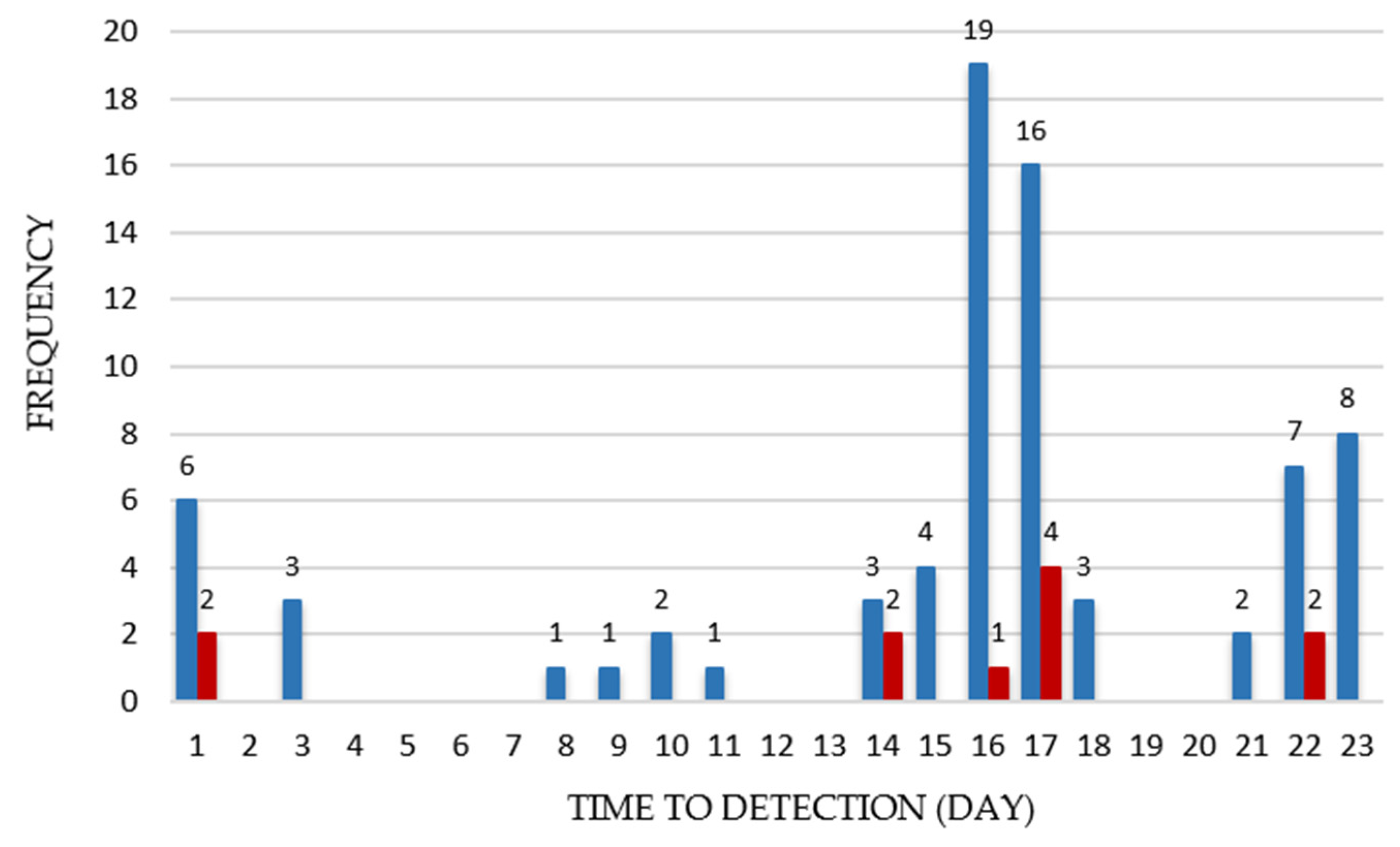{kind=link}
| Food Category | Food Subcategory | Total Number | Number of Positive Samples | |||
|---|---|---|---|---|---|---|
| nheA | hblA | cytK-1 | ces | |||
| Cereal products, including rice and seeds/pulses | 162 | 129 | 108 | 1 | 33 | |
| Cereal grains | 49 | 39 | 35 | - | 11 | |
| Cereal-based products and derivatives | 94 | 66 | 63 | - | 20 | |
| Seeds | 7 | 6 | 6 | |||
| Sprouts | 12 | 6 | 4 | 2 | ||
| Dairy-based products | 26 | 22 | 20 | 3 | 10 | |
| Milk powder and derivative products | 16 | 13 | 13 | 3 | 7 | |
| Coffee creamer | 7 | 7 | 5 | 1 | ||
| Chantilly cream | 3 | 2 | 2 | 2 | ||
| Dehydrated vegetables | Mashed potato flakes | 17 | 14 | 9 | 6 | 7 |
| Herbs and spices | Spices | 12 | 2 | 6 | ||
| Herbal teas | 8 | 6 | 4 | 1 | ||
| Mixed food | Ready-to-eat | 12 | 4 | 2 | ||
| Instant soups | 13 | 11 | 11 | 7 | ||
| Total | 226 | 176 | 162 | 10 | 58 | |
| Temperature (°C) | GDT (Day) | Total Number | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1–5 | 6–10 | 11–15 | 16–20 | 20–25 | Average | Standard Deviation | Median | Interquartile Range | ||
| 4 | 16 | 7 | 11 | 42 | 18 | 11 | 8.3 | 15 | 16 | 94 |
| 7 | 28 | 15 | 21 | 34 | 6 | 10 | 6.3 | 11 | 12 | 104 |
| 10 | 54 | 21 | 19 | 27 | - | 7 | 3.2 | 4 | 4 | 121 |
| Food Product | GDT at 4 °C | Cereulide Production at 37 °C (+/−) | Cereulide Production at 4 °C (ng/mL) |
|---|---|---|---|
| Buckwheat | 17 | + | − |
| Chantilly cream | 1 | + | − |
| Coffee creamer | 16 | + | − |
| Couscous | 14 | − | − |
| Couscous | 14 | − | |
| Milk powder | 1 | − | − |
| Muesli | 17 | − | − |
| Muesli | 17 | + | − |
| Polenta | 17 | − | − |
| Instant vermicelli soup | 22 | + | 0.79 |
| Instant soup | 22 | − | - |
| Food Category | Subcategory | Food Product | Number of Samples |
|---|---|---|---|
| Cereal products, including rice and seeds/pulses | Cereal grains | Buckwheat | 3 |
| Quinoa | 5 | ||
| Millet | 3 | ||
| Bulgur | 3 | ||
| Rice | 29 | ||
| Kamut | 1 | ||
| Cereal-based products and derivatives | Semolina | 4 | |
| Rice waffles | 7 | ||
| Breakfast cereals | 10 | ||
| Couscous | 5 | ||
| Polenta | 6 | ||
| Oat flakes | 5 | ||
| Flour | 13 | ||
| Pasta and pasta-like products | 50 | ||
| Seeds | 7 | ||
| Sprouted seeds | 12 | ||
| Dairy-based products | Milk powder | 8 | |
| Powder containing milk or whey protein (derivative products) | 8 | ||
| Coffee creamer | 7 | ||
| Chantilly cream | 3 | ||
| Dehydrated vegetables | Processed potato flakes | 17 | |
| Herbs and spices | Spices | 12 | |
| Herbal teas | 8 | ||
| Mixed food | Ready-to-eat | Pasta salads | 8 |
| Rice salads | 4 | ||
| Soups | Instant soups | 13 | |
| Total | 250 | ||
| Genes | Primers | Annealing Temperature (°C) | Reference |
|---|---|---|---|
| hblA_Taq | F: ATT AAT ACA GGG GAT GGA GAA ACT T R: TGA TCC TAA TAC TTC TTC TAG ACG CTT P: FAM/TGACTGCAA/ZEN/GAG CTCTTTATT | 52 | [77] |
| nheA_SYBR_G | F: TTC AAA TTC AAA AGA ATG TTG AAG AAG G R: GAT TTG TTT GCT TAT TCA TTT CAT CAC | 60 | [78] |
| cytK1_SYBR_G | F: GCT TTG TAT AAG CAA CTT GGA TAG R: AGC CTC TGT AAC ACC AAG C | 60 | |
| ces_SYBR_G | F: CAC GCC GAA AGT GAT TAT ACC AA R: CAC GAT AAA ACC ACT GAG ATA GTG | 60 | [79] |
| ces_Taq | F: CGC CGA AAG TGA TTA TAC CAA R: TAT GCC CCG TTC TCA AAC TG P: FAM/GGG AAA ATA ACG AGA AAT GCA/TAMRA | 60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jovanovic, J.; Tretiak, S.; Begyn, K.; Rajkovic, A. Detection of Enterotoxigenic Psychrotrophic Presumptive Bacillus cereus and Cereulide Producers in Food Products and Ingredients. Toxins 2022, 14, 289. https://doi.org/10.3390/toxins14040289
Jovanovic J, Tretiak S, Begyn K, Rajkovic A. Detection of Enterotoxigenic Psychrotrophic Presumptive Bacillus cereus and Cereulide Producers in Food Products and Ingredients. Toxins. 2022; 14(4):289. https://doi.org/10.3390/toxins14040289
Chicago/Turabian StyleJovanovic, Jelena, Svitlana Tretiak, Katrien Begyn, and Andreja Rajkovic. 2022. "Detection of Enterotoxigenic Psychrotrophic Presumptive Bacillus cereus and Cereulide Producers in Food Products and Ingredients" Toxins 14, no. 4: 289. https://doi.org/10.3390/toxins14040289
APA StyleJovanovic, J., Tretiak, S., Begyn, K., & Rajkovic, A. (2022). Detection of Enterotoxigenic Psychrotrophic Presumptive Bacillus cereus and Cereulide Producers in Food Products and Ingredients. Toxins, 14(4), 289. https://doi.org/10.3390/toxins14040289