Development and Characterization of Anti-Naja ashei Three-Finger Toxins (3FTxs)-Specific Monoclonal Antibodies and Evaluation of Their In Vitro Inhibition Activity

 ,
,  ,
,  , ,
, ,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
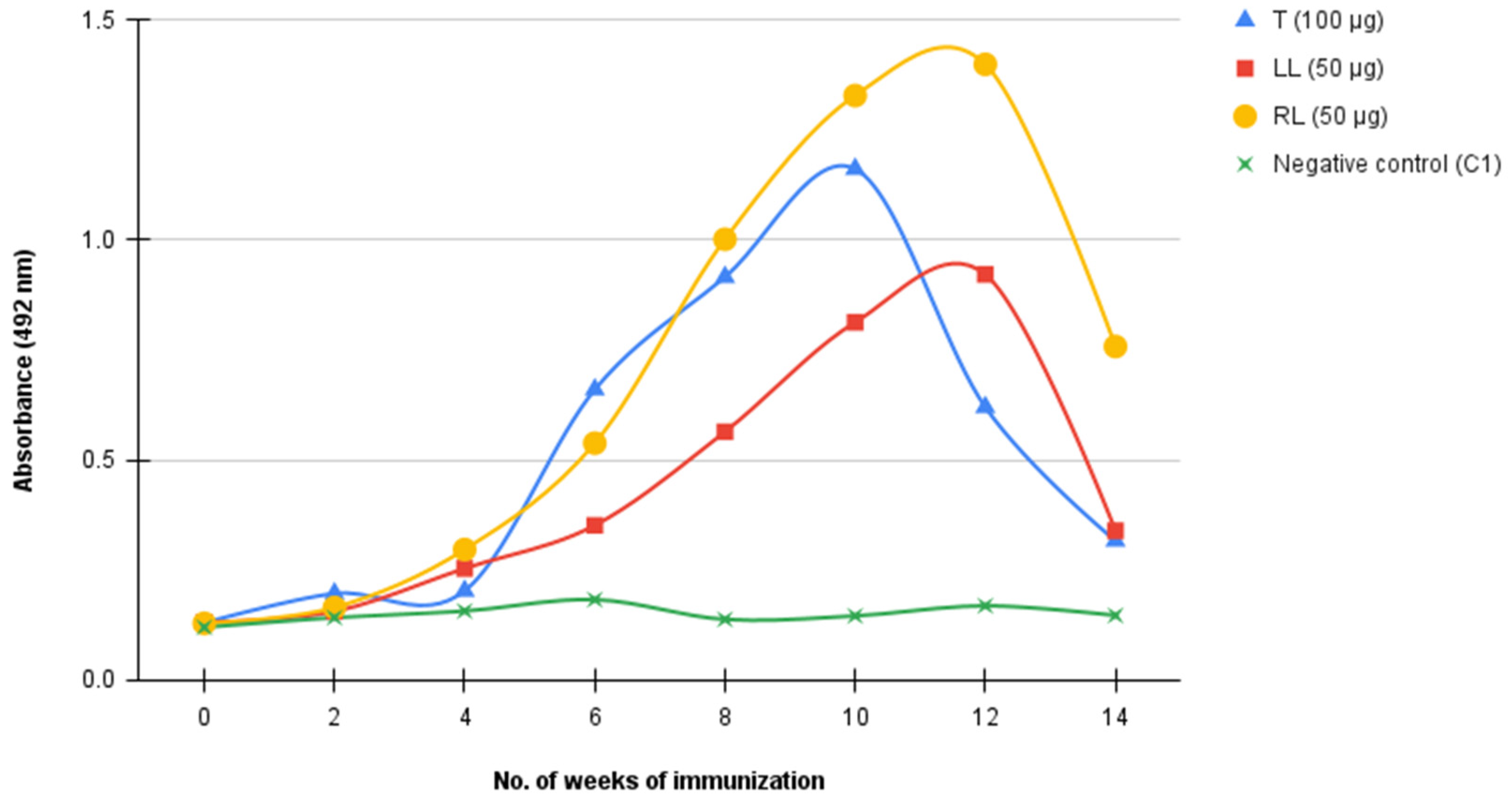
2.1. Immune Response against 3FTxs Antigen and Antibody Titer Determination
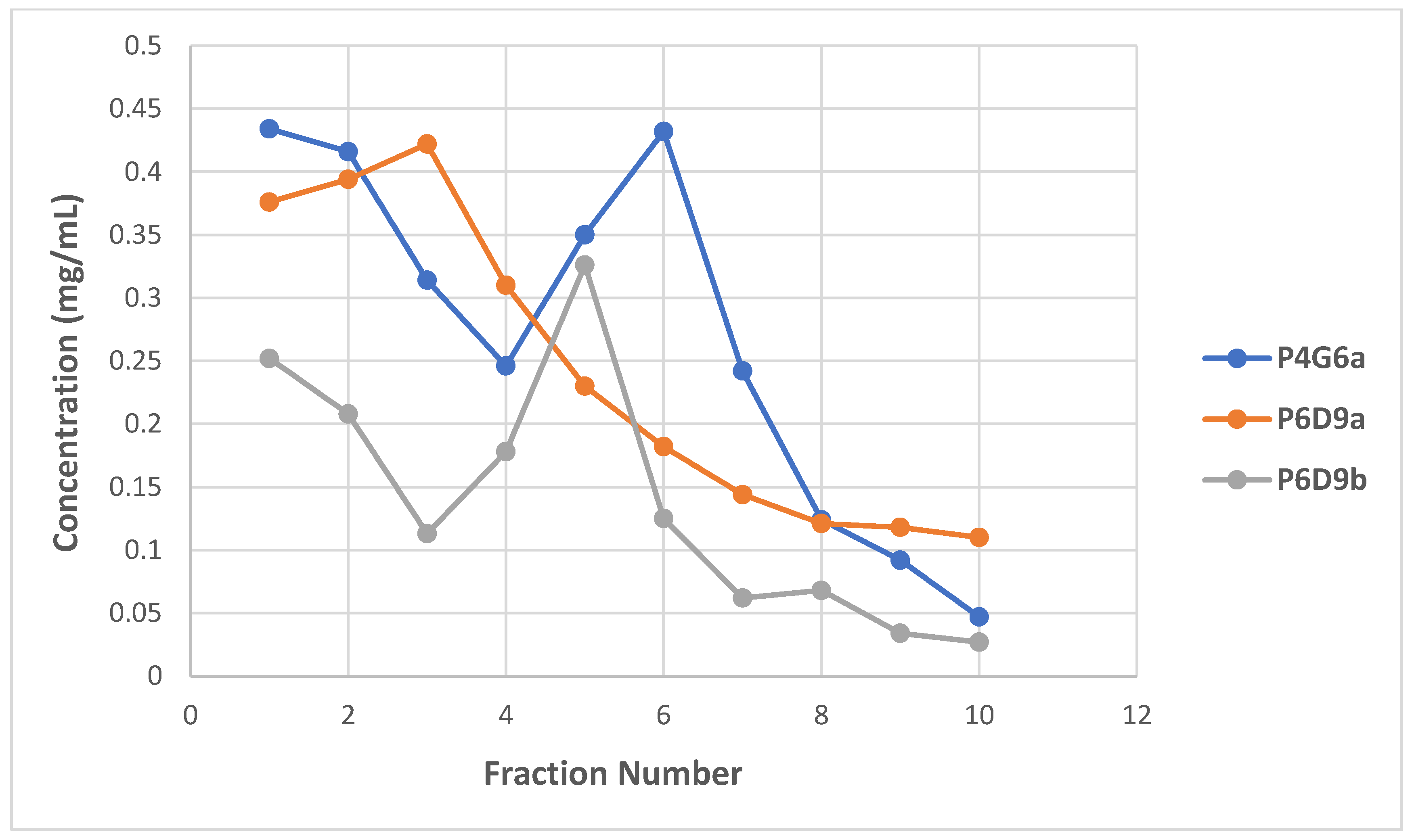
2.2. Characterization of mAbs
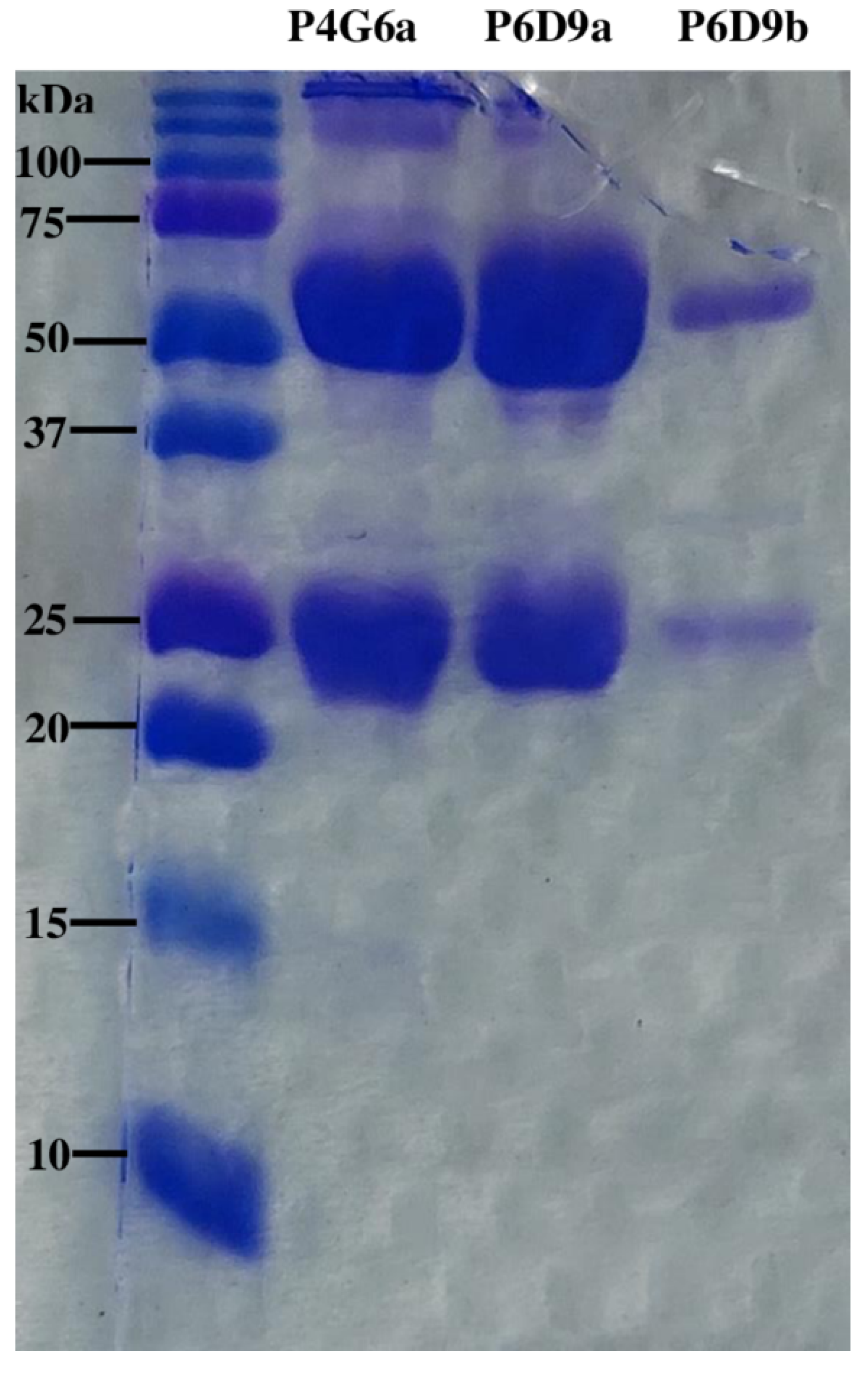
SDS-PAGE Analysis of mAbs
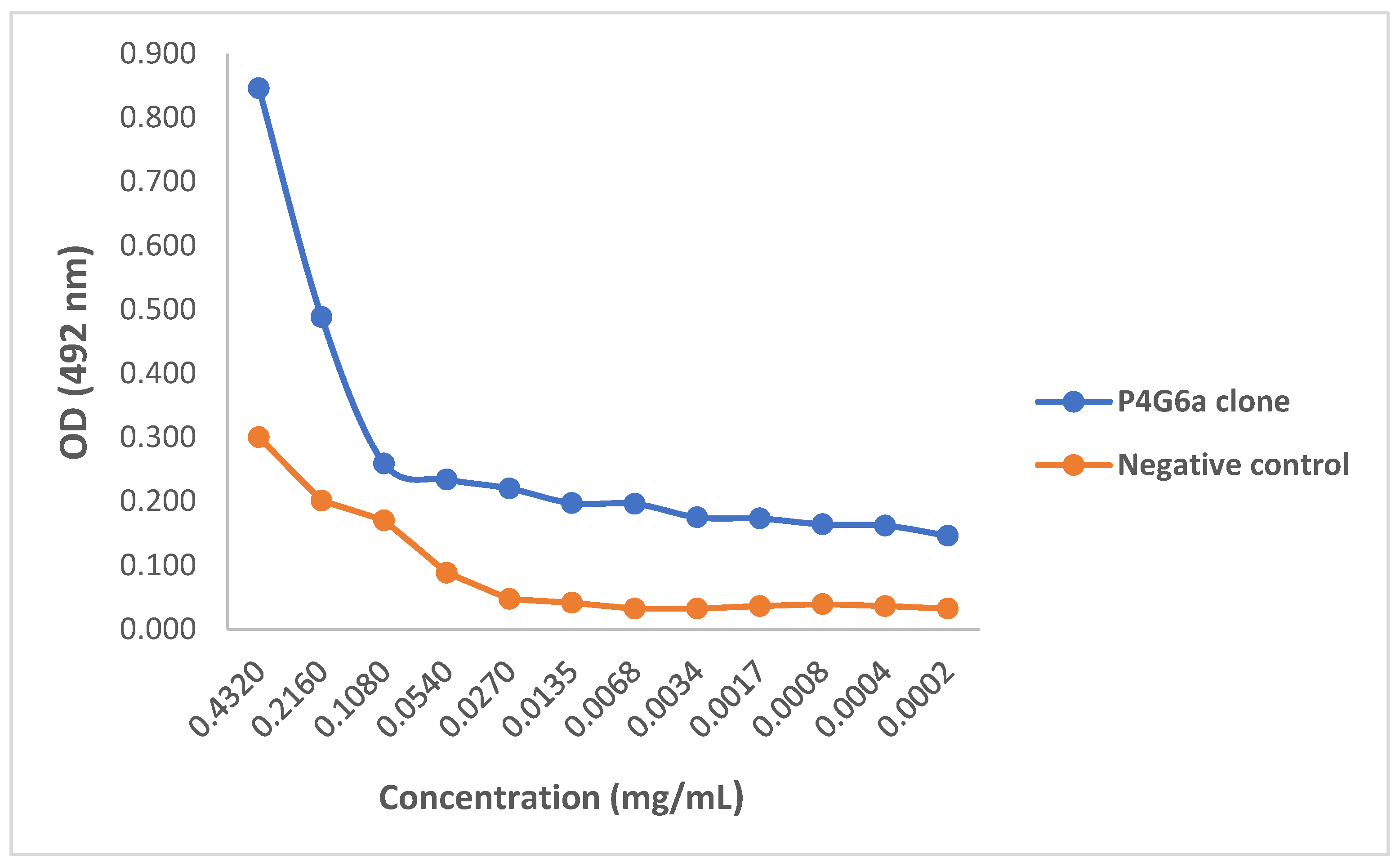
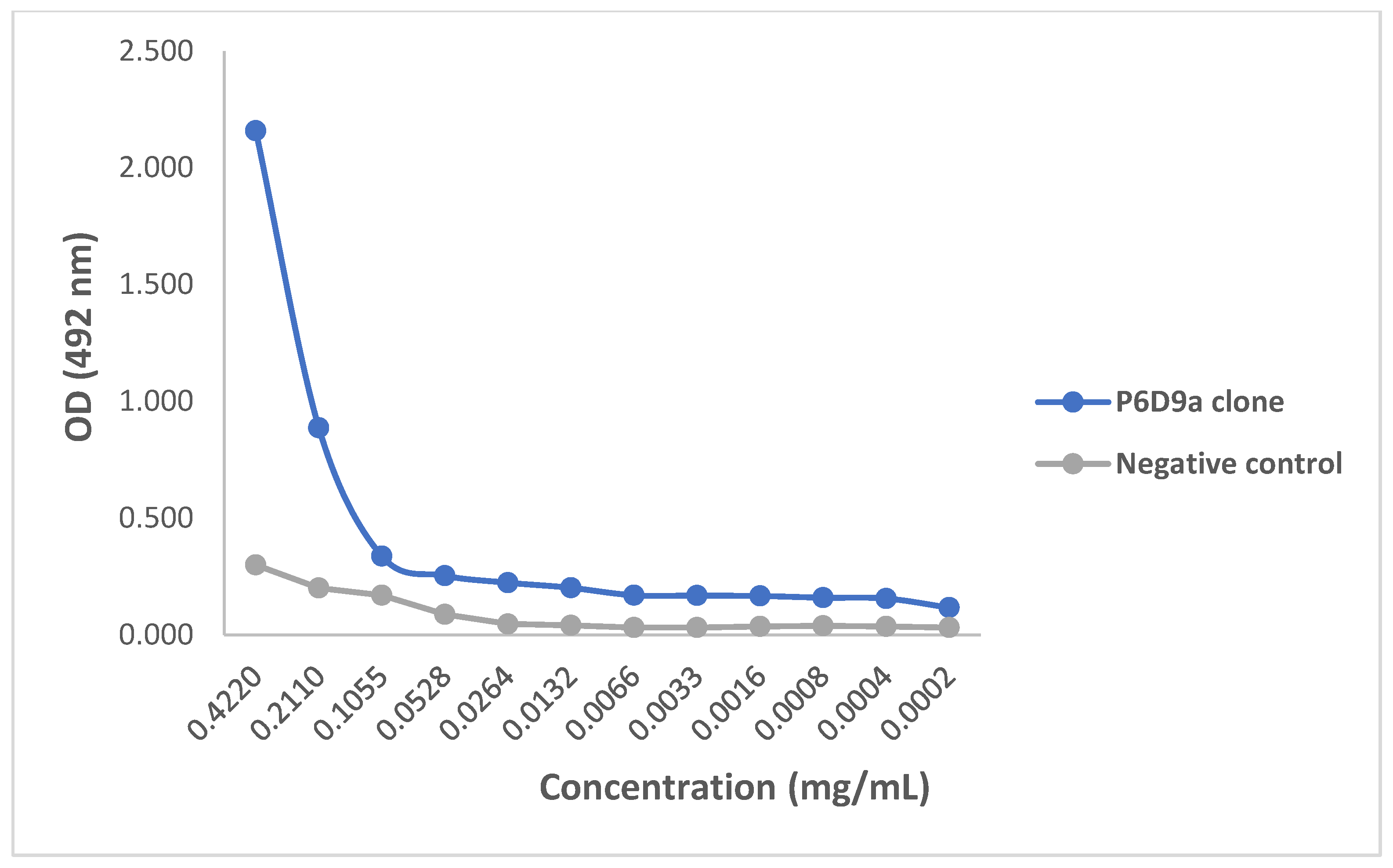
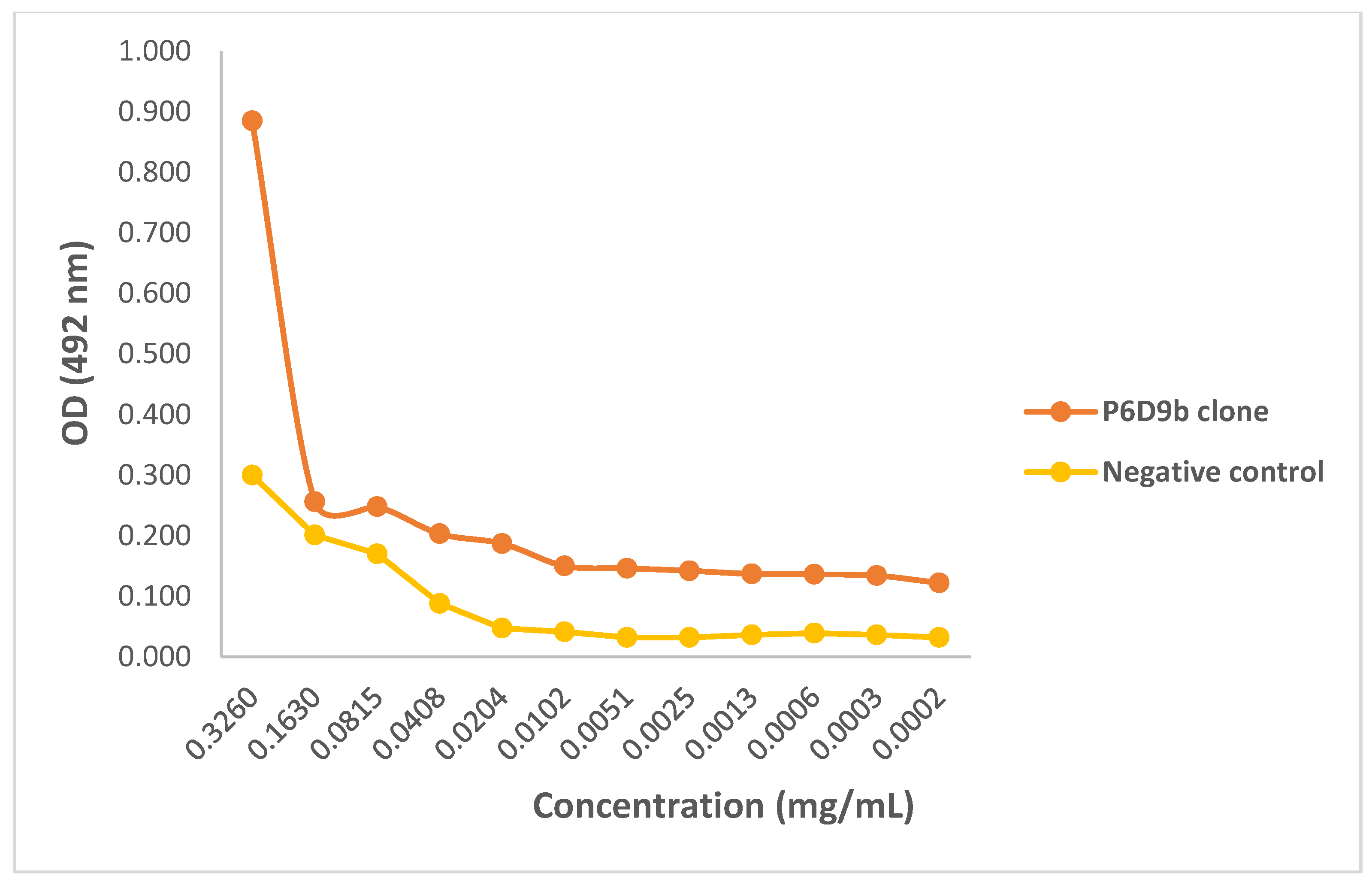
2.3. Evaluating Binding Efficacy of Purified mAbs
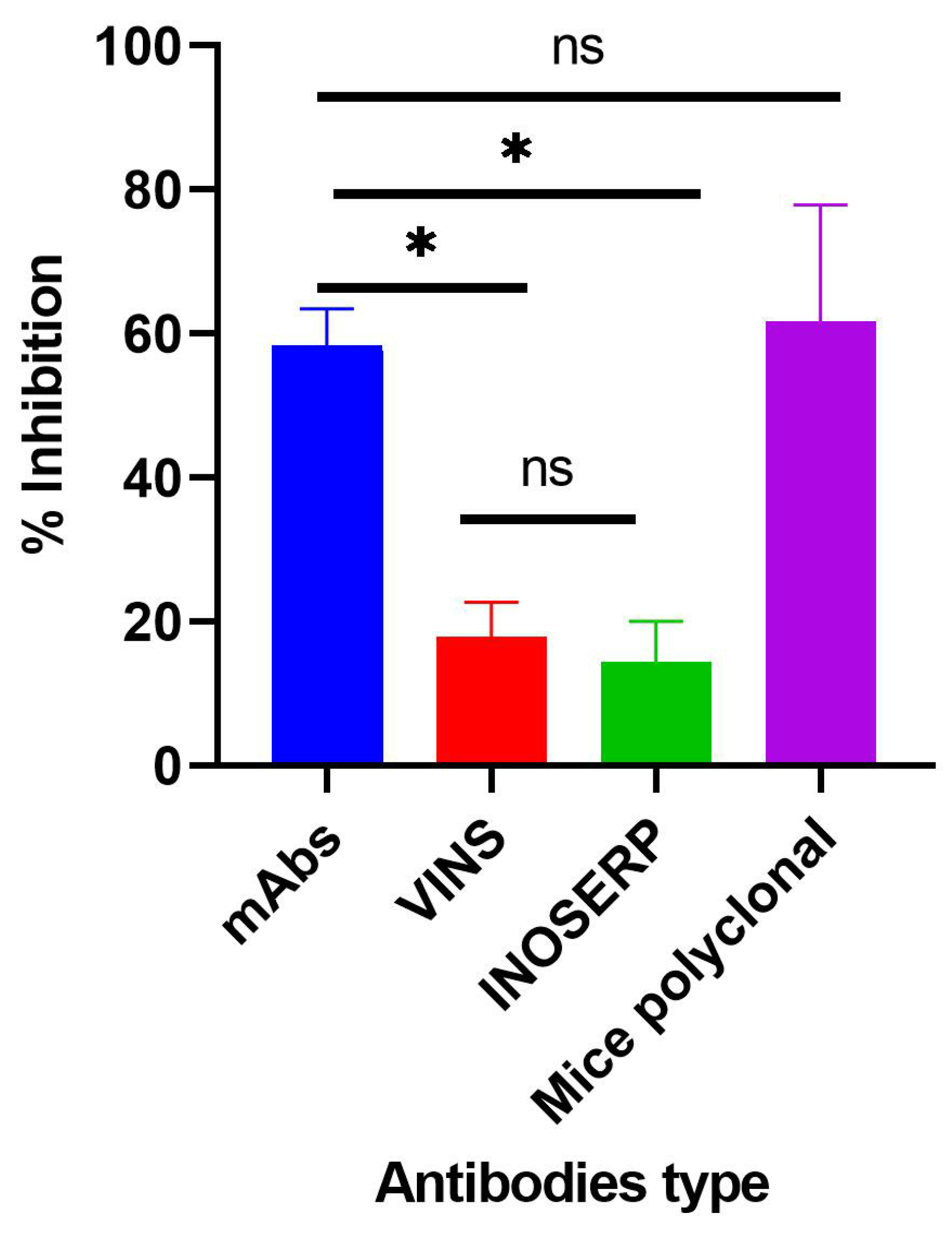
2.4. Evaluating In Vitro Activity of Purified mAbs
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Three-Finger Toxins and Venoms
5.2. Antivenoms
5.3. Animals
5.4. Ethical Review
5.5. Optimization of ELISA Parameters
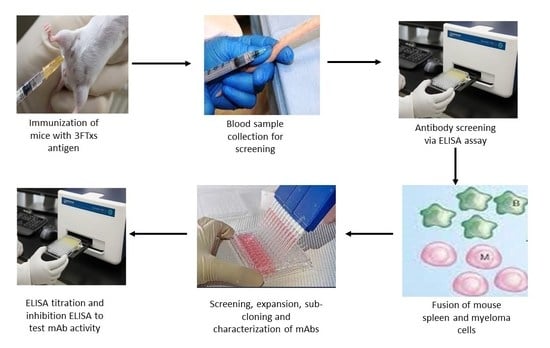
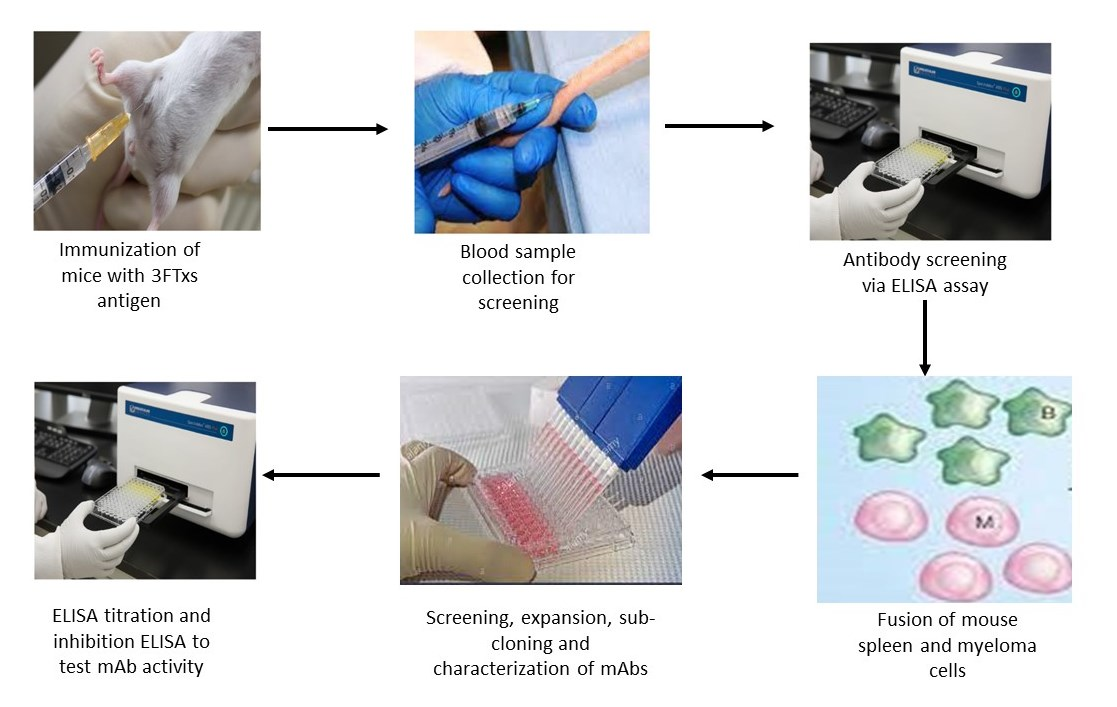
5.6. Generation of Monoclonal Antibodies
5.7. Screening of Serum and Hybridoma Supernatants for Antibodies
5.8. Characterization of mAbs
SDS-PAGE Analysis of mAbs
5.9. Monoclonal Antibody Titration Using ELISA
5.10. Evaluating Inhibitory Activity of Generated mAbs
5.11. Statistical Analysis
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Prim. 2017, 3, nrdp201763. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-H.; Lee, Y.-C.; Liang, M.-H.; Leu, S.-J.; Lin, L.-T.; Chiang, J.-R.; Yang, Y.-Y. Antibodies against Venom of the Snake Deinagkistrodon acutus. Appl. Environ. Microbiol. 2016, 82, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Sajevic, T.; Leonardi, A.; Križaj, I. Haemostatically active proteins in snake venoms. Toxicon 2011, 57, 627–645. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Snakebite Envenoming. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/snakebite-envenoming (accessed on 15 January 2022).
- Nkwescheu, A.; Mbasso, L.C.D.; Pouth, F.B.B.; Dzudie, A.; Billong, S.C.; Ngouakam, H.; Diffo, J.L.D.; Eyongorock, H.; Mbacham, W. Snakebite in bedroom kills a physician in Cameroon: A case report. Pan Afr. Med. J. 2016, 24, 231. [Google Scholar] [CrossRef]
- Harrison, R.A.; Oluoch, G.O.; Ainsworth, S.; Alsolaiss, J.; Bolton, F.; Arias, A.-S.; Gutiérrez, J.-M.; Rowley, P.; Kalya, S.; Ozwara, H.; et al. Preclinical antivenom-efficacy testing reveals potentially disturbing deficiencies of snakebite treatment capability in East Africa. PLoS Negl. Trop. Dis. 2017, 11, e0005969. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Warrell, D.A.; Williams, D.J.; Jensen, S.; Brown, N.; Calvete, J.J.; Harrison, R.A. The Need for the Global Snakebite Initiative The Need for Full Integration of Snakebite Envenoming within a Global Strategy to Combat the Neglected Tropical Diseases: The Way Forward. PLoS Negl. Trop. Dis. 2013, 7, e2162. [Google Scholar] [CrossRef]
- Ministry of Health. Guidelines for Prevention, Diagnosis and Management of Snakebite Envenoming in Kenya. Nairobi 2019. Available online: https://kma.co.ke/Documents/Snakebite%20Envenoming%20in%20Kenya.pdf (accessed on 20 March 2020).
- WHO. Who Expert Committee on Biological Standardization. 2017. Available online: https://www.who.int/groups/expert-committee-on-biological-standardization (accessed on 18 February 2020).
- Okumu, M.O.; Mbaria, J.M.; Gikunju, J.K.; Mbuthia, P.G.; Madadi, V.O.; Ochola, F.O. Enzymatic activity and brine shrimp lethality of venom from the large brown spitting cobra (Naja ashei) and its neutralization by antivenom. BMC Res. Notes 2020, 13, 1–7. [Google Scholar] [CrossRef]
- Hus, K.K.; Buczkowicz, J.; Petrilla, V.; Petrillová, M.; Łyskowski, A.; Legáth, J.; Bocian, A. First Look at the Venom of Naja ashei. Molecules 2018, 23, 609. [Google Scholar] [CrossRef]
- Hus, K.K.; Marczak, Ł.; Petrilla, V.; Petrillová, M.; Legáth, J.; Bocian, A. Different Research Approaches in Unraveling the Venom Proteome of Naja ashei. Biomolecules 2020, 10, 1282. [Google Scholar] [CrossRef]
- Bermúdez-Méndez, E.; Fuglsang-Madsen, A.; Føns, S.; Lomonte, B.; Gutiérrez, J.M.; Laustsen, A.H. Innovative Immunization Strategies for Antivenom Development. Toxins 2018, 10, 452. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M.; Doley, R. Structure, function and evolution of three-finger toxins: Mini proteins with multiple targets. Toxicon 2010, 56, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Williams, H.F.; Layfield, H.J.; Vallance, T.; Patel, K.; Bicknell, A.B.; Trim, S.A.; Vaiyapuri, S. The Urgent Need to Develop Novel Strategies for the Diagnosis and Treatment of Snakebites. Toxins 2019, 11, 363. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.L.; Fan, H.W.; França, F.O.; Jorge, M.T.; Leite, R.P.; Nishioka, S.A.; Avila, A.; Sano-Martins, I.S.; Tomy, S.C.; Santoro, M.L. Randomized comparative trial of three antivenoms in the treatment of envenoming by lance-headed vipers (Bothrops jararaca) in São Paulo, Brazil. Q. J. Med. 1993, 86, 315–325. [Google Scholar] [PubMed]
- Otero-Patiño, R.; Diaz, A.; Higashi, H.G.; Cardoso, J.L.; Murcia, M.; Garcia, L.F.; Silva-Diaz, J.F.; Sierra, A.; Medina, M.C.; Castañeda, N.; et al. A randomized, blinded, comparative trial of one pepsin-digested and two whole IgG antivenoms for Bothrops snake bites in Uraba, Colombia. The Regional Group on Antivenom Therapy Research (REGATHER). Am. J. Trop. Med. Hyg. 1998, 58, 183–189. [Google Scholar] [CrossRef]
- Chippaux, J.-P.; Goyffon, M. Antivenom serotherapy: Its applications, its limitations, its future. Bull. Société Pathol. Exot. 1991, 84, 286–297. [Google Scholar]
- Chippaux, J.P. Production and use of snake antivenom. In Handbook of Natural Toxins, 5th ed.; Tu, T., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 1991; pp. 529–555. [Google Scholar]
- Harrison, R.A.; Cook, D.A.; Renjifo, C.; Casewell, N.R.; Currier, R.B.; Wagstaff, S.C. Research strategies to improve snakebite treatment: Challenges and progress. J. Proteom. 2011, 74, 1768–1780. [Google Scholar] [CrossRef]
- Frauches, T.S.; Petretski, J.H.; Arnholdt, A.C.; Lasunskaia, E.B.; de Carvalho, E.C.; Kipnis, T.L.; da Silva, W.D.; Kanashiro, M.M. Bothropic antivenom based on monoclonal antibodies, is it possible? Toxicon 2013, 71, 49–56. [Google Scholar] [CrossRef]
- Harrison, R. Development of venom toxin-specific antibodies by DNA immunisation: Rationale and strategies to improve therapy of viper envenoming. Vaccine 2004, 22, 1648–1655. [Google Scholar] [CrossRef]
- Tan, C.H.; Tan, K.Y.; Lim, S.E.; Tan, N.H. Venomics of the beaked sea snake, Hydrophis schistosus: A minimalist toxin arsenal and its cross-neutralization by heterologous antivenoms. J. Proteom. 2015, 126, 121–130. [Google Scholar] [CrossRef]
- Tan, K.Y.; Tan, C.H.; Fung, S.Y.; Tan, N.H. Neutralization of the Principal Toxins from the Venoms of Thai Naja kaouthia and Malaysian Hydrophis schistosus: Insights into Toxin-Specific Neutralization by Two Different Antivenoms. Toxins 2016, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Engmark, M.; Clouser, C.; Timberlake, S.; Vigneault, F.; Gutiérrez, J.M.; Lomonte, B. Exploration of immunoglobulin transcriptomes from mice immunized with three-finger toxins and phospholipases A2 from the Central American coral snake, Micrurus nigrocinctus. PeerJ 2017, 5, e2924. [Google Scholar] [CrossRef] [PubMed]
- Moroz-Perlmutter, C.; Goldblum, N.; De Vries, A.; Gitter, S. Detoxification of Snake Venoms and Venom Fractions by Formaldehyde. Exp. Biol. Med. 1963, 112, 595–598. [Google Scholar] [CrossRef]
- Lakkappa, D.B.; Menon, J.; Kumar, D.; Oommen, V.O.; Joseph, K.J. Snake Venom Detection Kit (SVDK): Update on Current Aspects and Challenges. In Clinical Toxinology in Asia Pacific and Africa; Gopalakrishnakone, P., Faiz, A., Fernando, R., Gnanathasan, C.A., Habib, A.G., Yang, C., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 379–400. [Google Scholar]
- Leow, C.H.; Jones, M.; Cheng, Q.; Mahler, S.; McCarthy, J. Production and characterization of specific monoclonal antibodies binding the Plasmodium falciparum diagnostic biomarker, histidine-rich protein 2. Malar. J. 2014, 13, 277. [Google Scholar] [CrossRef] [PubMed]
- Stiles, B.G.; Sexton, F.W.; Guest, S.B.; Olson, M.A.; Hack, D.C. Characterization of monoclonal antibodies against Naja naja oxiana neurotoxin I. Biochem. J. 1994, 303, 163–170. [Google Scholar] [CrossRef][Green Version]
- Dyba, B.; Rudolphi-Szydło, E.; Barbasz, A.; Czyżowska, A.; Hus, K.; Petrilla, V.; Petrillová, M.; Legáth, J.; Bocian, A. Effects of 3FTx Protein Fraction from Naja ashei Venom on the Model and Native Membranes: Recognition and Implications for the Mechanisms of Toxicity. Molecules 2021, 26, 2164. [Google Scholar] [CrossRef]
- Manson, E.Z.; Mutinda, K.C.; Gikunju, J.K.; Bocian, A.; Hus, K.K.; Petrílla, V.; Legáth, J.; Kimotho, J.H. Development of an Inhibition Enzyme-Linked Immunosorbent Assay (ELISA) Prototype for Detecting Cytotoxic Three-Finger Toxins (3FTxs) in African Spitting Cobra Venoms. Molecules 2022, 27, 888. [Google Scholar] [CrossRef]
- Himananto, O.; Thummabenjapone, P.; Luxananil, P.; Kumpoosiri, M.; Hongprayoon, R.; Kositratana, W.; Gajanandana, O. Novel and Highly Specific Monoclonal Antibody to Acidovorax citrulli and Development of ELISA-Based Detection in Cucurbit Leaves and Seed. Plant Dis. 2011, 95, 1172–1178. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manson, E.Z.; Kyama, M.C.; Kimani, J.; Bocian, A.; Hus, K.K.; Petrilla, V.; Legáth, J.; Kimotho, J.H. Development and Characterization of Anti-Naja ashei Three-Finger Toxins (3FTxs)-Specific Monoclonal Antibodies and Evaluation of Their In Vitro Inhibition Activity. Toxins 2022, 14, 285. https://doi.org/10.3390/toxins14040285
Manson EZ, Kyama MC, Kimani J, Bocian A, Hus KK, Petrilla V, Legáth J, Kimotho JH. Development and Characterization of Anti-Naja ashei Three-Finger Toxins (3FTxs)-Specific Monoclonal Antibodies and Evaluation of Their In Vitro Inhibition Activity. Toxins. 2022; 14(4):285. https://doi.org/10.3390/toxins14040285
Chicago/Turabian StyleManson, Ernest Z., Mutinda C. Kyama, Josephine Kimani, Aleksandra Bocian, Konrad K. Hus, Vladimír Petrilla, Jaroslav Legáth, and James H. Kimotho. 2022. "Development and Characterization of Anti-Naja ashei Three-Finger Toxins (3FTxs)-Specific Monoclonal Antibodies and Evaluation of Their In Vitro Inhibition Activity" Toxins 14, no. 4: 285. https://doi.org/10.3390/toxins14040285
APA StyleManson, E. Z., Kyama, M. C., Kimani, J., Bocian, A., Hus, K. K., Petrilla, V., Legáth, J., & Kimotho, J. H. (2022). Development and Characterization of Anti-Naja ashei Three-Finger Toxins (3FTxs)-Specific Monoclonal Antibodies and Evaluation of Their In Vitro Inhibition Activity. Toxins, 14(4), 285. https://doi.org/10.3390/toxins14040285