Allergen Content of Therapeutic Preparations for Allergen-Specific Immunotherapy of European Paper Wasp Venom Allergy

 , ,
, ,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
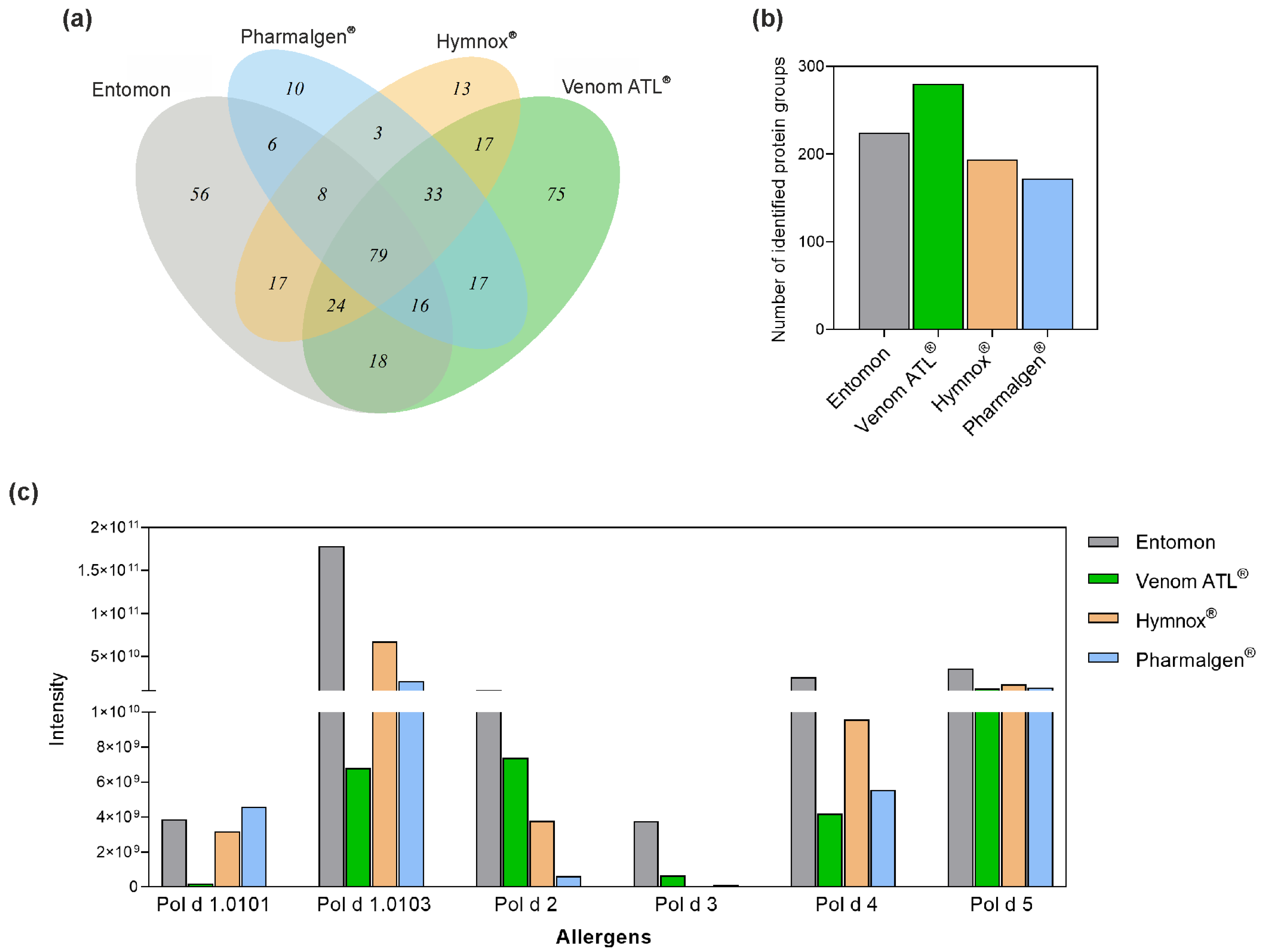
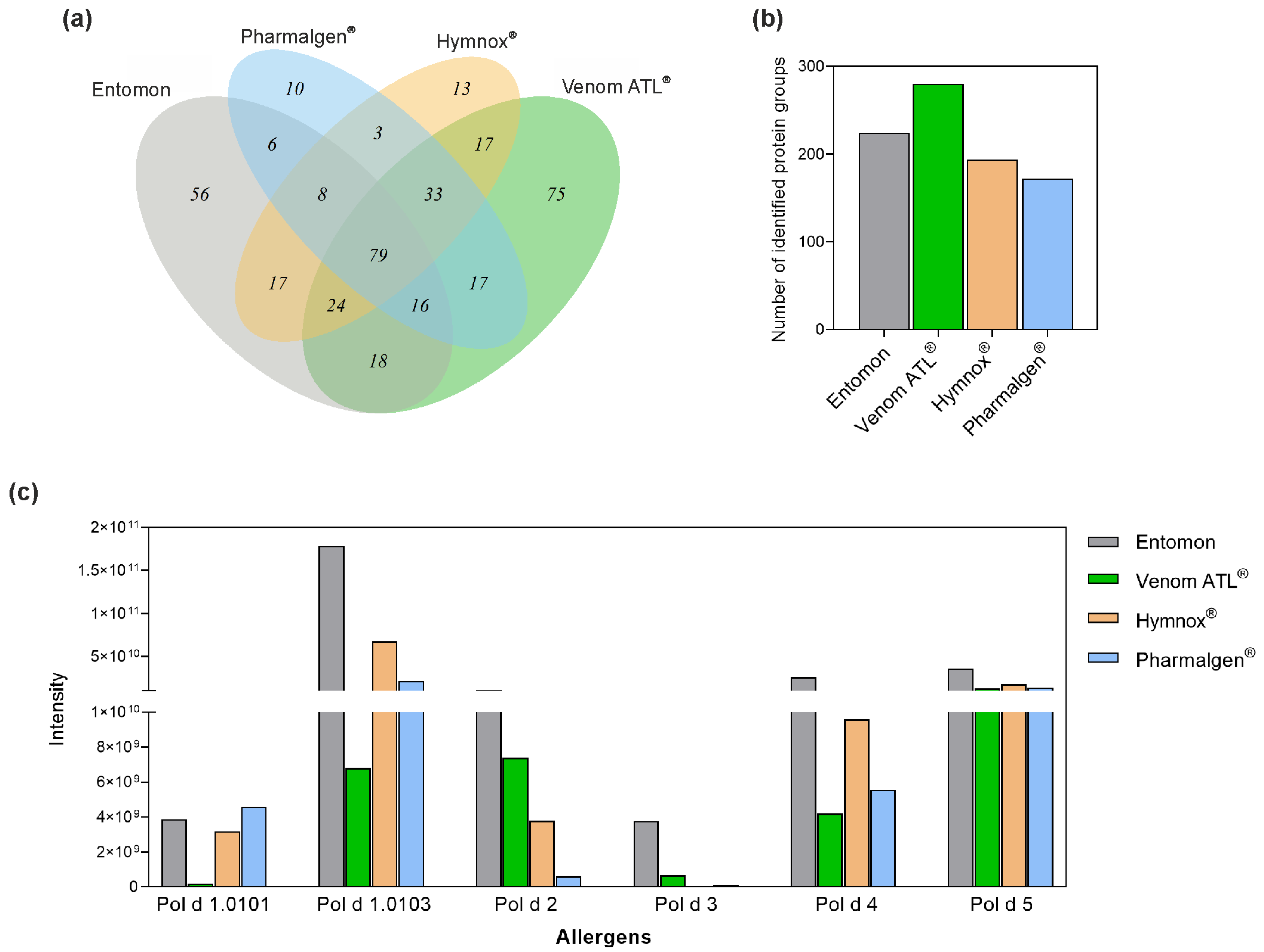
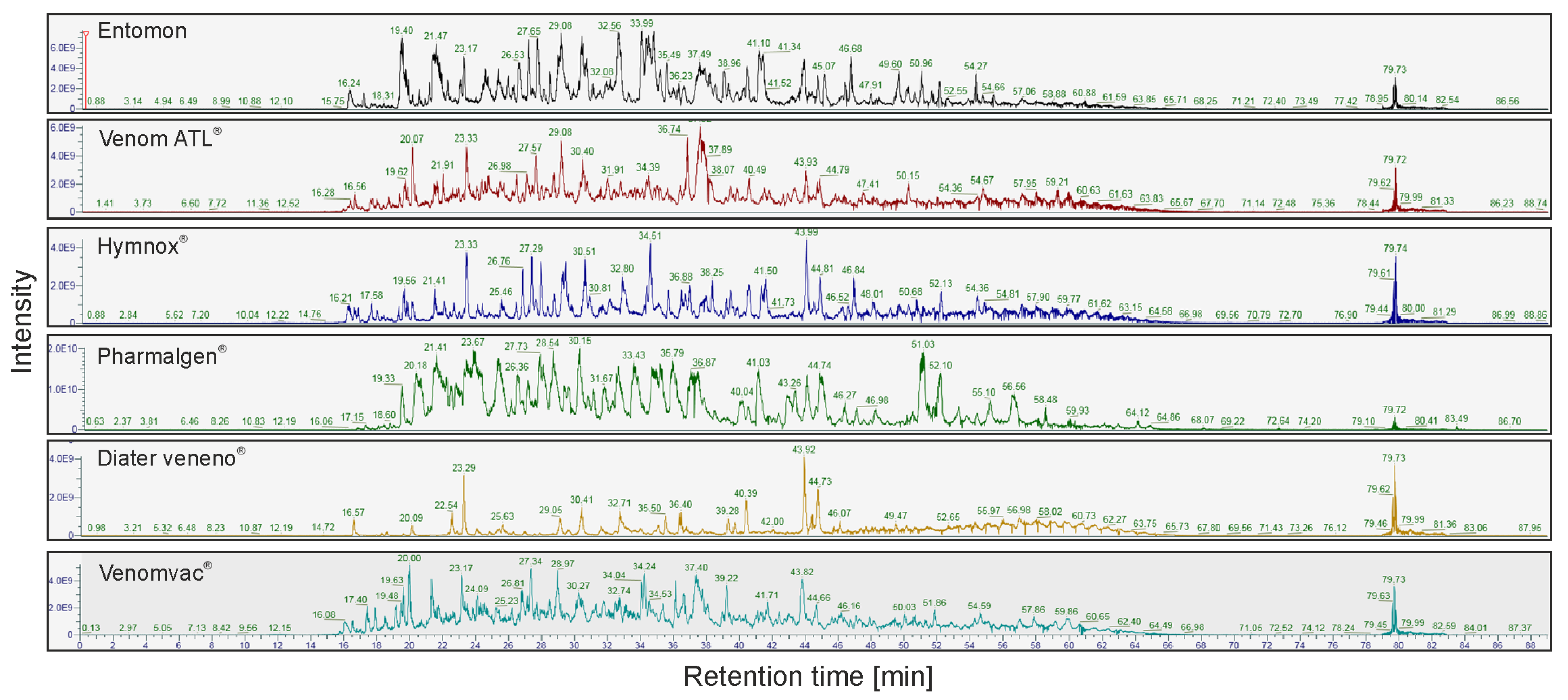
2.1. LC-MS/MS of P. dominula Venom-Containing Venom Preparations
2.2. SDS-PAGE Analysis of Polistes Venom Preparations
2.3. LC-MS of Polistes Venom Preparations
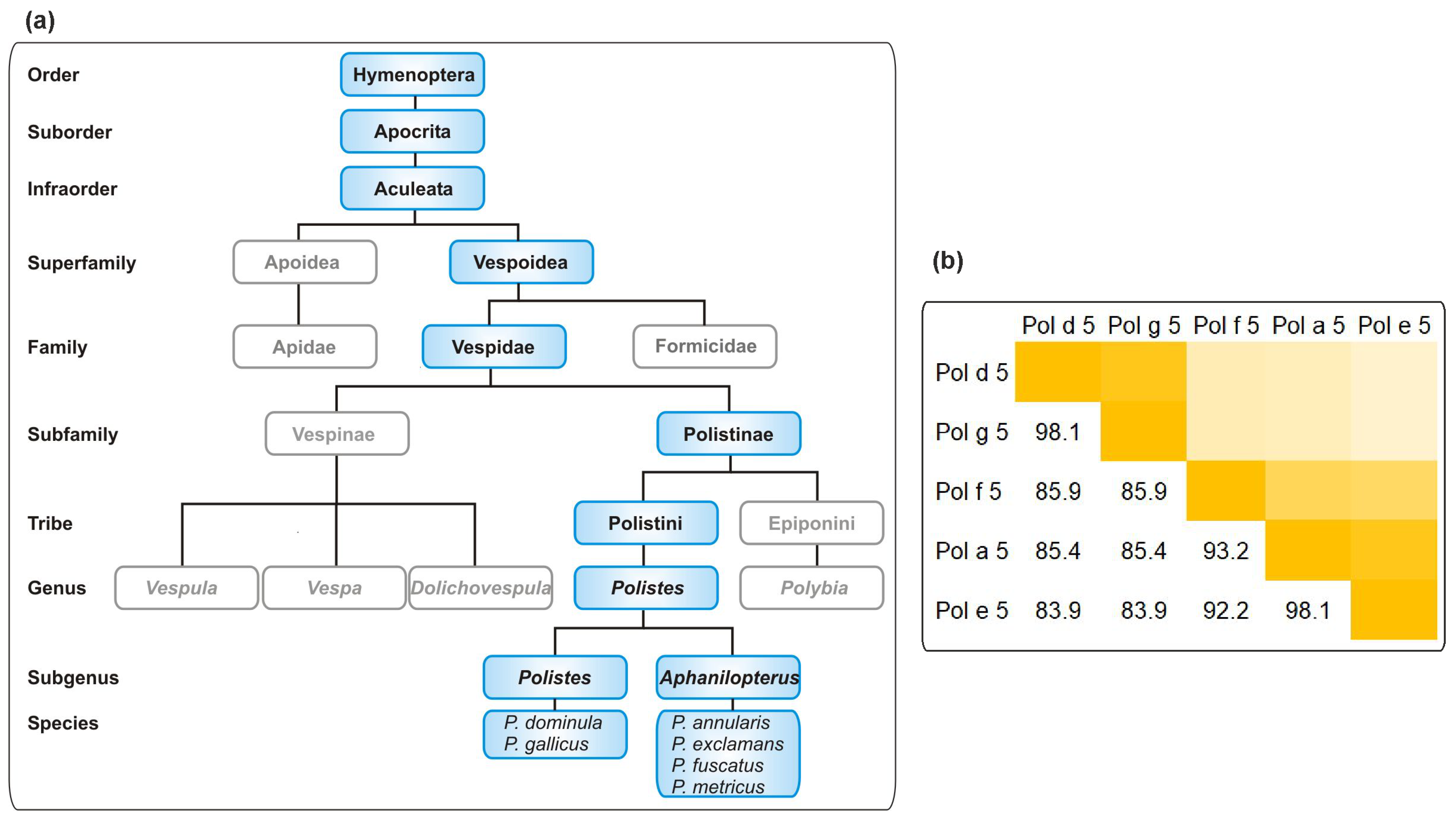
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Products
5.2. SDS-PAGE
5.3. Sample Preparation
5.4. Liquid Chromatography-Mass Spectrometry
5.5. Data Processing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bilo, M.B.; Ollert, M.; Blank, S. The role of component-resolved diagnosis in Hymenoptera venom allergy. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Blank, S.; Bilo, M.B.; Ollert, M. Component-resolved diagnostics to direct in venom immunotherapy: Important steps towards precision medicine. Clin. Exp. Allergy 2018, 48, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Blank, S.; Grosch, J.; Ollert, M.; Bilo, M.B. Precision Medicine in Hymenoptera Venom Allergy: Diagnostics, Biomarkers, and Therapy of Different Endotypes and Phenotypes. Front. Immunol. 2020, 11, 579409. [Google Scholar] [CrossRef] [PubMed]
- Grosch, J.; Eberlein, B.; Waldherr, S.; Pascal, M.; San Bartolome, C.; De La Roca Pinzon, F.; Dittmar, M.; Hilger, C.; Ollert, M.; Biedermann, T.; et al. Characterization of New Allergens from the Venom of the European Paper Wasp Polistes dominula. Toxins 2021, 13, 559. [Google Scholar] [CrossRef]
- Grosch, J.; Hilger, C.; Bilo, M.B.; Kler, S.; Schiener, M.; Dittmar, G.; Bernardin, F.; Lesur, A.; Ollert, M.; Schmidt-Weber, C.B.; et al. Shedding Light on the Venom Proteomes of the Allergy-Relevant Hymenoptera Polistes dominula (European Paper Wasp) and Vespula spp. (Yellow Jacket). Toxins 2020, 12, 323. [Google Scholar] [CrossRef]
- Schiener, M.; Hilger, C.; Eberlein, B.; Pascal, M.; Kuehn, A.; Revets, D.; Planchon, S.; Pietsch, G.; Serrano, P.; Moreno-Aguilar, C.; et al. The high molecular weight dipeptidyl peptidase IV Pol d 3 is a major allergen of Polistes dominula venom. Sci. Rep. 2018, 8, 1318. [Google Scholar] [CrossRef]
- Buck, M.; Marshall, S.A.; Cheung, D.K.B. Identification atlas of the Vespidae (Hymenoptera, Aculeata) of the northeastern Nearctic Region. Can. J. Arthropod Identif. 2008, 5, 1–492. [Google Scholar]
- Cervo, R.; Zacchi, F.; Turillazzi, S. Polistes dominulus (Hymenoptera, Vespidae) invading North America: Some hypotheses for its rapid spread. Insects Sociaux 2000, 47, 155–157. [Google Scholar] [CrossRef]
- Eardley, C.; Koch, F.; Wood, A.R. Polistes dominulus (Christ, 1791) (Hymenoptera: Polistinae: Vespidae) newly recorded from South Africa: Short communication. Afr. Entomol. 2009, 17, 226–227. [Google Scholar] [CrossRef]
- Gamboa, G.J.; Noble, M.A.; Thom, M.C.; Togal, J.L.; Srinivasan, R.; Murphy, B.D. The comparative biology of two sympatric paper wasps in Michigan, the native Polistes fuscatus and the invasive Polistes dominulus (Hymenoptera, Vespidae). Insects Sociaux 2003, 51, 153–157. [Google Scholar] [CrossRef]
- Weiner, S.A.; Noble, K.; Upton, C.T.; Flynn, G.; Woods, W.A.; Starks, P.T. The cost of flight: A role in the Polistes dominulus invasion. Insects Sociaux 2012, 59, 81–86. [Google Scholar] [CrossRef]
- Demain, J.G.; Gessner, B.D.; McLaughlin, J.B.; Sikes, D.S.; Foote, J.T. Increasing insect reactions in Alaska: Is this related to changing climate? Allergy Asthma Proc. 2009, 30, 238–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blank, S.; Bazon, M.L.; Grosch, J.; Schmidt-Weber, C.B.; Brochetto-Braga, M.R.; Bilo, M.B.; Jakob, T. Antigen 5 Allergens of Hymenoptera Venoms and Their Role in Diagnosis and Therapy of Venom Allergy. Curr. Allergy Asthma Rep. 2020, 20, 58. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, J.M. Distributional Checklist of Species of the Genus Polistes. Am. Museum Novit. 1996, 3188, 1–40. [Google Scholar]
- Nguyen, L.T.P.; Carpenter, J. Taxonomic notes on the paper wasps of the subgenus Polistes (Gyrostoma) (Hymenoptera, Vespidae, Polistinae) occurring in Vietnam, with description of a new species. J. Hymenopt. Res. 2019, 74, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Caruso, B.; Bonadonna, P.; Severino, M.G.; Manfredi, M.; Dama, A.; Schiappoli, M.; Rizzotti, P.; Senna, G.; Passalacqua, G. Evaluation of the IgE cross-reactions among vespid venoms. A possible approach for the choice of immunotherapy. Allergy 2007, 62, 561–564. [Google Scholar] [CrossRef]
- Monsalve, R.I.; Vega, A.; Marques, L.; Miranda, A.; Fernandez, J.; Soriano, V.; Cruz, S.; Dominguez-Noche, C.; Sanchez-Morillas, L.; Armisen-Gil, M.; et al. Component-resolved diagnosis of vespid venom-allergic individuals: Phospholipases and antigen 5s are necessary to identify Vespula or Polistes sensitization. Allergy 2012, 67, 528–536. [Google Scholar] [CrossRef]
- Quercia, O.; Cova, V.; Martini, M.; Cortellini, G.; Murzilli, F.; Bignardi, D.; Cilia, M.; Scarpa, A.; Bilo, M.B. CAP-Inhibition, Molecular Diagnostics, and Total IgE in the Evaluation of Polistes and Vespula Double Sensitization. Int. Arch. Allergy Immunol. 2018, 177, 365–369. [Google Scholar] [CrossRef]
- Sturm, G.J.; Varga, E.M.; Roberts, G.; Mosbech, H.; Bilo, M.B.; Akdis, C.A.; Antolin-Amerigo, D.; Cichocka-Jarosz, E.; Gawlik, R.; Jakob, T.; et al. EAACI guidelines on allergen immunotherapy: Hymenoptera venom allergy. Allergy 2018, 73, 744–764. [Google Scholar] [CrossRef] [Green Version]
- Jakob, T.; Müller, U.; Helbling, A.; Spillner, E. Component resolved diagnostics for hymenoptera venom allergy. Curr. Opin. Allergy Clin. Immunol. 2017, 17, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Frick, M.; Fischer, J.; Helbling, A.; Rueff, F.; Wieczorek, D.; Ollert, M.; Pfutzner, W.; Muller, S.; Huss-Marp, J.; Dorn, B.; et al. Predominant Api m 10 sensitization as risk factor for treatment failure in honey bee venom immunotherapy. J. Allergy Clin. Immunol. 2016, 138, 1663–1671 e1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonertz, A.; Roberts, G.C.; Hoefnagel, M.; Timon, M.; Slater, J.E.; Rabin, R.L.; Bridgewater, J.; Pini, C.; Pfaar, O.; Akdis, C.; et al. Challenges in the implementation of EAACI guidelines on allergen immunotherapy: A global perspective on the regulation of allergen products. Allergy 2018, 73, 64–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rueff, F.; Przybilla, B.; Bilo, M.B.; Muller, U.; Scheipl, F.; Seitz, M.J.; Aberer, W.; Bodzenta-Lukaszyk, A.; Bonifazi, F.; Campi, P.; et al. Clinical effectiveness of hymenoptera venom immunotherapy: A prospective observational multicenter study of the European academy of allergology and clinical immunology interest group on insect venom hypersensitivity. PLoS ONE 2013, 8, e63233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rueff, F.; Vos, B.; Oude Elberink, J.; Bender, A.; Chatelain, R.; Dugas-Breit, S.; Horny, H.P.; Kuchenhoff, H.; Linhardt, A.; Mastnik, S.; et al. Predictors of clinical effectiveness of Hymenoptera venom immunotherapy. Clin. Exp. Allergy 2014, 44, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Blank, S.; Etzold, S.; Darsow, U.; Schiener, M.; Eberlein, B.; Russkamp, D.; Wolf, S.; Graessel, A.; Biedermann, T.; Ollert, M.; et al. Component-resolved evaluation of the content of major allergens in therapeutic extracts for specific immunotherapy of honeybee venom allergy. Hum. Vaccin Immunother. 2017, 13, 2482–2489. [Google Scholar] [CrossRef]
- Blank, S.; Seismann, H.; Michel, Y.; McIntyre, M.; Cifuentes, L.; Braren, I.; Grunwald, T.; Darsow, U.; Ring, J.; Bredehorst, R.; et al. Api m 10, a genuine A. mellifera venom allergen, is clinically relevant but underrepresented in therapeutic extracts. Allergy 2011, 66, 1322–1329. [Google Scholar] [CrossRef]
- Brunetto, B.; Tinghino, R.; Braschi, M.C.; Antonicelli, L.; Pini, C.; Iacovacci, P. Characterization and comparison of commercially available mite extracts for in vivo diagnosis. Allergy 2010, 65, 184–190. [Google Scholar] [CrossRef]
- Focke, M.; Marth, K.; Flicker, S.; Valenta, R. Heterogeneity of commercial timothy grass pollen extracts. Clin. Exp. Allergy 2008, 38, 1400–1408. [Google Scholar] [CrossRef]
- Heneberg, P.; Riegerova, K.; Rihova, A.; Simcikova, D.; Kucera, P. Updates on the surface antigens of basophils: CD16 on basophils of patients with respiratory or insect venom allergy and the rejection of CD203c and CD63 externalization decoupling by bisindolylmaleimides. Clin. Exp. Allergy 2019, 49, 54–67. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.F.; Du Toit, G.; Douiri, A.; Radulovic, S.; Stephens, A.; Turcanu, V.; Lack, G. Distinct parameters of the basophil activation test reflect the severity and threshold of allergic reactions to peanut. J. Allergy Clin. Immunol. 2015, 135, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Marsh, D.G.; Goodfriend, L.; King, T.P.; Lowenstein, H.; Platts-Mills, T.A. Allergen nomenclature. Bull. World Health Organ. 1986, 64, 767–774. [Google Scholar] [PubMed]
- Radauer, C.; Nandy, A.; Ferreira, F.; Goodman, R.E.; Larsen, J.N.; Lidholm, J.; Pomes, A.; Raulf-Heimsoth, M.; Rozynek, P.; Thomas, W.R.; et al. Update of the WHO/IUIS Allergen Nomenclature Database based on analysis of allergen sequences. Allergy 2014, 69, 413–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blank, S.; Bilò, M.B.; Grosch, J.; Schmidt-Weber, C.B.; Ollert, M.; Jakob, T. Marker allergens in Hymenoptera venom allergy—Characteristics and potential use in precision medicine. Allergo J. Int. 2021, 30, 26–38. [Google Scholar] [CrossRef]
- Spiric, J.; Paulus, K.E.; Schwaben, L.; Vieths, S.; Junker, A.; Mahler, V.; Reuter, A. Complete compositional analysis of honeybee venom therapeutic products by mass spectrometry. Confirmation of all known Api m allergens in one blow. Allergy 2018, 73, 342. [Google Scholar]
- Schiener, M.; Eberlein, B.; Moreno-Aguilar, C.; Pietsch, G.; Serrano, P.; McIntyre, M.; Schwarze, L.; Russkamp, D.; Biedermann, T.; Spillner, E.; et al. Application of recombinant antigen 5 allergens from seven allergy-relevant Hymenoptera species in diagnostics. Allergy 2017, 72, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Casset, A.; Mari, A.; Purohit, A.; Resch, Y.; Weghofer, M.; Ferrara, R.; Thomas, W.R.; Alessandri, C.; Chen, K.W.; de Blay, F.; et al. Varying allergen composition and content affects the in vivo allergenic activity of commercial Dermatophagoides pteronyssinus extracts. Int. Arch. Allergy Immunol. 2012, 159, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Curin, M.; Reininger, R.; Swoboda, I.; Focke, M.; Valenta, R.; Spitzauer, S. Skin prick test extracts for dog allergy diagnosis show considerable variations regarding the content of major and minor dog allergens. Int. Arch. Allergy Immunol. 2011, 154, 258–263. [Google Scholar] [CrossRef]
- Sanchez, F.; Blanca, M.; Fernandez, J.; Miranda, A.; Terrados, A.; Torres, M.J.; Del Cano, A.; Garcia, J.J.; Juarez, C. Comparative study between European and American species of Polistes using sera from European sensitized subjects. Clin. Exp. Allergy 1995, 25, 281–287. [Google Scholar] [CrossRef]
- Severino, M.G.; Campi, P.; Macchia, D.; Manfredi, M.; Turillazzi, S.; Spadolini, I.; Bilo, M.B.; Bonifazi, F. European Polistes venom allergy. Allergy 2006, 61, 860–863. [Google Scholar] [CrossRef]
- Bonadonna, P.; Caruso, B.; Labardi, D.; Dama, A.; Senna, G.; Passalacqua, G. Treatment with American Polistes venom was ineffective in an Italian patient allergic to European Polistes. Allergy 2007, 62, 966–967. [Google Scholar] [CrossRef]
- Dalmau Duch, G.; Gazquez Garcia, V.; Gaig Jane, P.; Galan Nieto, A.; Monsalve Clemente, R.I. Importance of controlled sting challenge and component-resolved diagnosis in the success of venom immunotherapy. J. Investig. Allergol. Clin. Immunol. 2012, 22, 135–136. [Google Scholar] [PubMed]
- Savi, E.; Incorvaia, C.; Boni, E.; Mauro, M.; Peveri, S.; Pravettoni, V.; Quercia, O.; Reccardini, F.; Montagni, M.; Pessina, L.; et al. Which immunotherapy product is better for patients allergic to Polistes venom? A laboratory and clinical study. PLoS ONE 2017, 12, e0180270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, C.S.; Foehr, S.; Garfield, D.A.; Furlong, E.E.; Steinmetz, L.M.; Krijgsveld, J. Ultrasensitive proteome analysis using paramagnetic bead technology. Mol. Syst. Biol. 2014, 10, 757. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Company | Product Name | Abbreviation | Species |
|---|---|---|---|
| Entomon | Entomon Capillary Extracted Venom® P. dominula | Entomon | P. dominula |
| Allergy Therapeutics | Venom ATL Polistes® | Venom ATL® | P. dominula |
| ROXALL Medicina | Hymnox® | Hymnox® | P. dominula |
| ALK-Abelló | Pharmalgen® | Pharmalgen® | P. dominula |
| Diater Pharmaceutical Laboratory | Diater Veneno de Himenópteros Polistes spp.® | Diater veneno® | Polistes spp.1 |
| LETI Pharma | Venomvac® | Venomvac® | Polistes spp.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grosch, J.; Lesur, A.; Kler, S.; Bernardin, F.; Dittmar, G.; Francescato, E.; Hewings, S.J.; Jakwerth, C.A.; Zissler, U.M.; Heath, M.D.; et al. Allergen Content of Therapeutic Preparations for Allergen-Specific Immunotherapy of European Paper Wasp Venom Allergy. Toxins 2022, 14, 284. https://doi.org/10.3390/toxins14040284
Grosch J, Lesur A, Kler S, Bernardin F, Dittmar G, Francescato E, Hewings SJ, Jakwerth CA, Zissler UM, Heath MD, et al. Allergen Content of Therapeutic Preparations for Allergen-Specific Immunotherapy of European Paper Wasp Venom Allergy. Toxins. 2022; 14(4):284. https://doi.org/10.3390/toxins14040284
Chicago/Turabian StyleGrosch, Johannes, Antoine Lesur, Stéphanie Kler, François Bernardin, Gunnar Dittmar, Elisabetta Francescato, Simon J. Hewings, Constanze A. Jakwerth, Ulrich M. Zissler, Matthew D. Heath, and et al. 2022. "Allergen Content of Therapeutic Preparations for Allergen-Specific Immunotherapy of European Paper Wasp Venom Allergy" Toxins 14, no. 4: 284. https://doi.org/10.3390/toxins14040284
APA StyleGrosch, J., Lesur, A., Kler, S., Bernardin, F., Dittmar, G., Francescato, E., Hewings, S. J., Jakwerth, C. A., Zissler, U. M., Heath, M. D., Ollert, M., Kramer, M. F., Hilger, C., Bilò, M. B., Schmidt-Weber, C. B., & Blank, S. (2022). Allergen Content of Therapeutic Preparations for Allergen-Specific Immunotherapy of European Paper Wasp Venom Allergy. Toxins, 14(4), 284. https://doi.org/10.3390/toxins14040284