Cloning and Characterization of Three Novel Enzymes Responsible for the Detoxification of Zearalenone


Abstract
:1. Introduction
2. Results
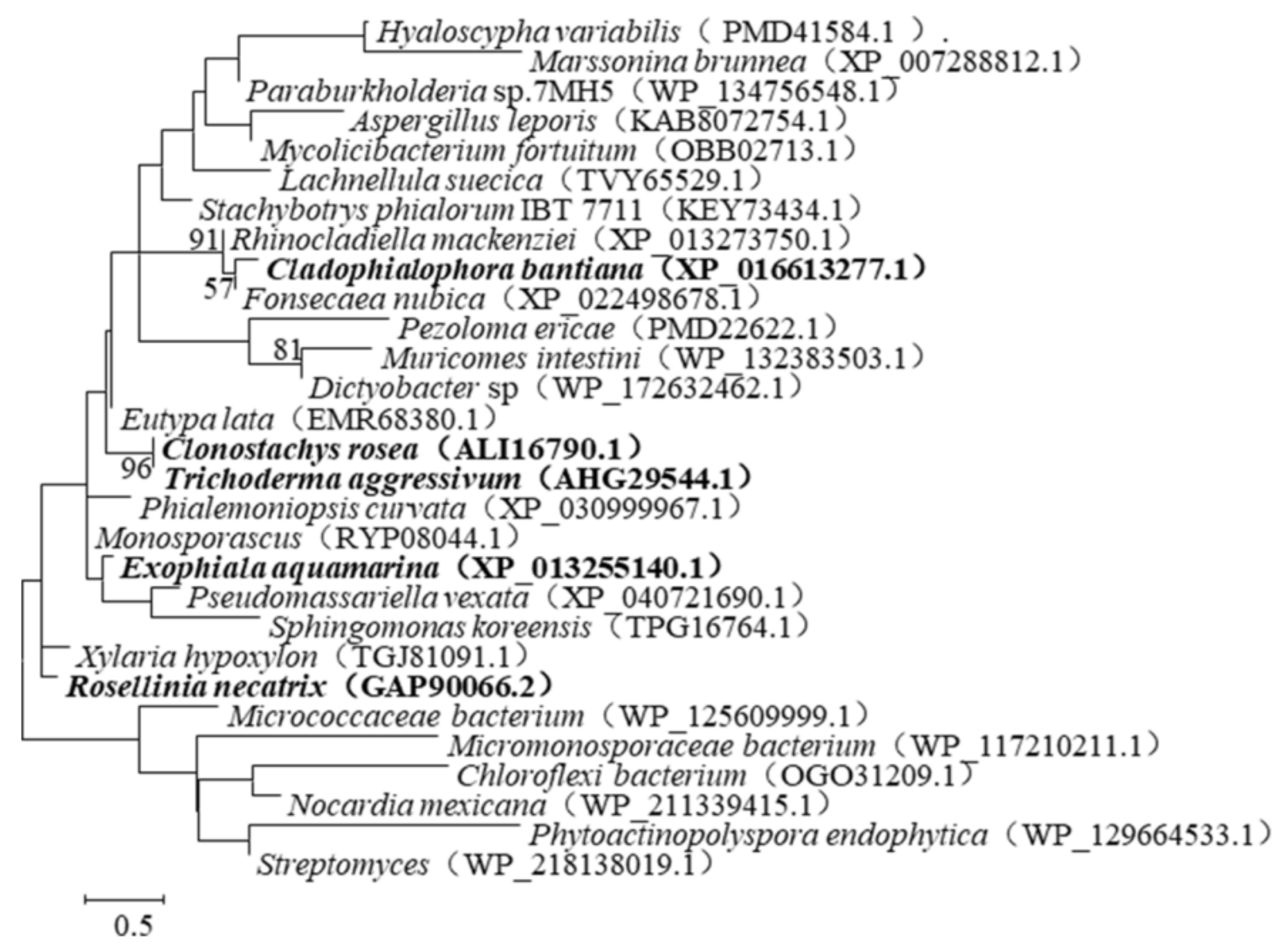
2.1. Selection of the ZEN-Degrading Enzymes
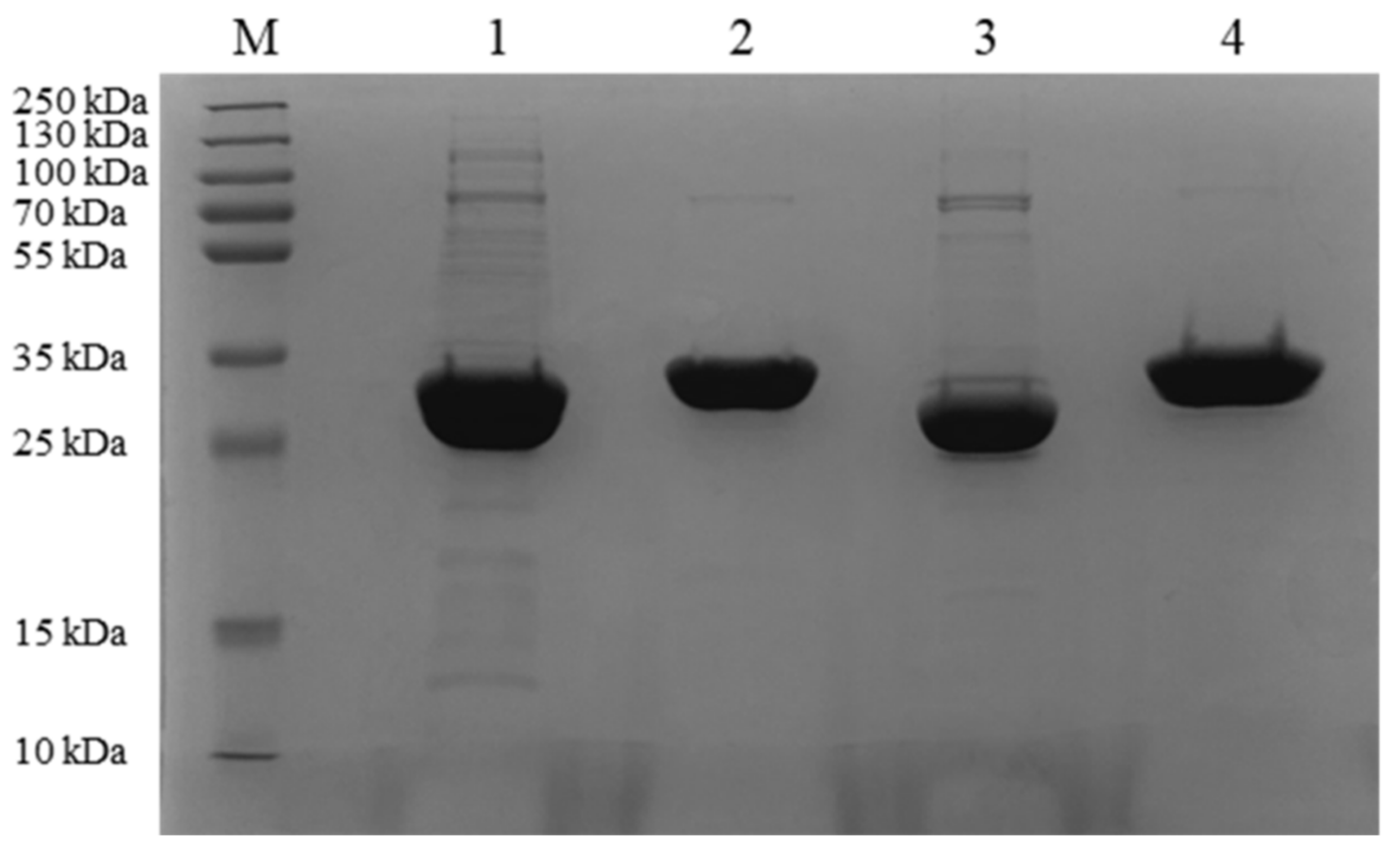
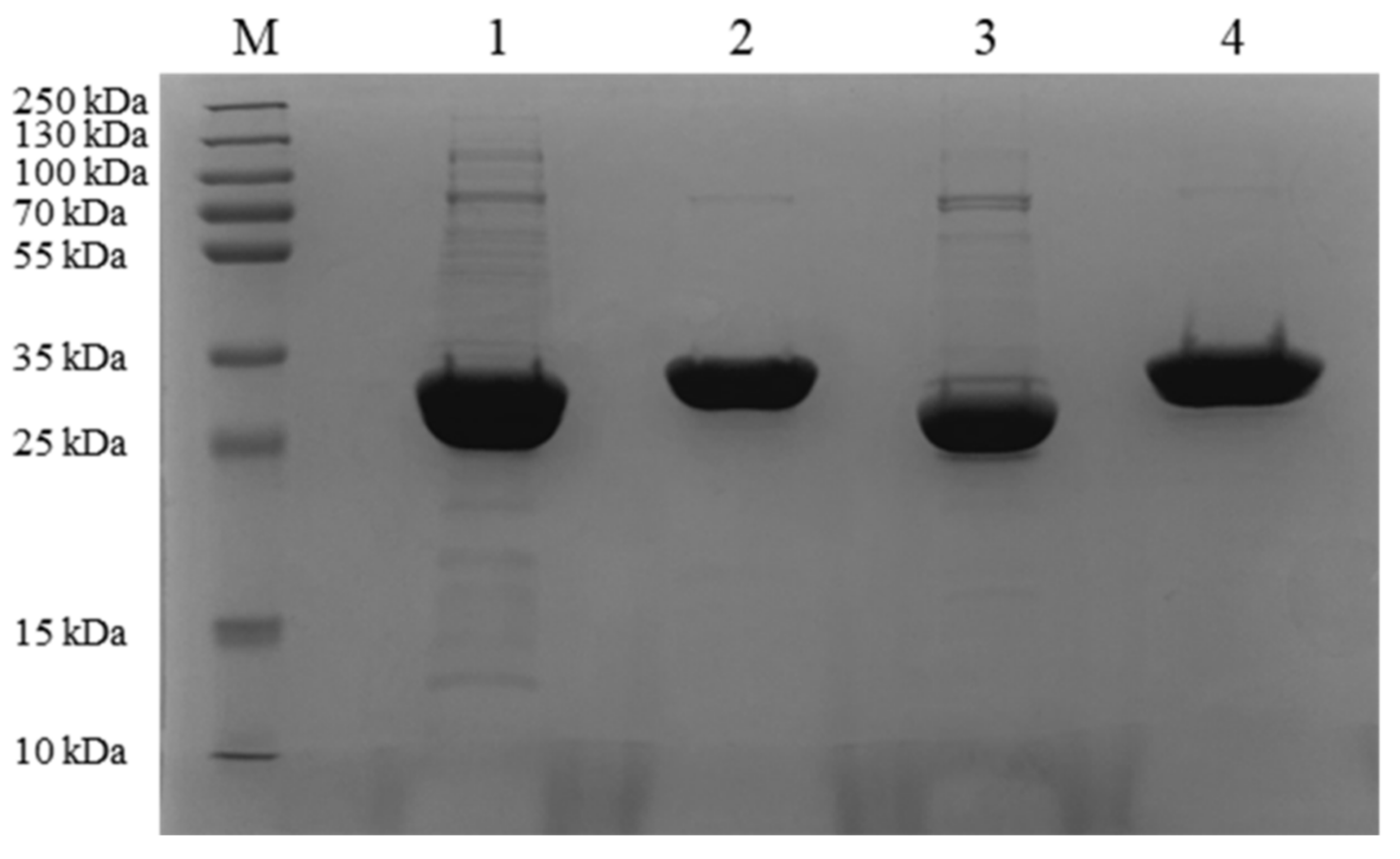
2.2. Expression and Purification of the Four Proteins
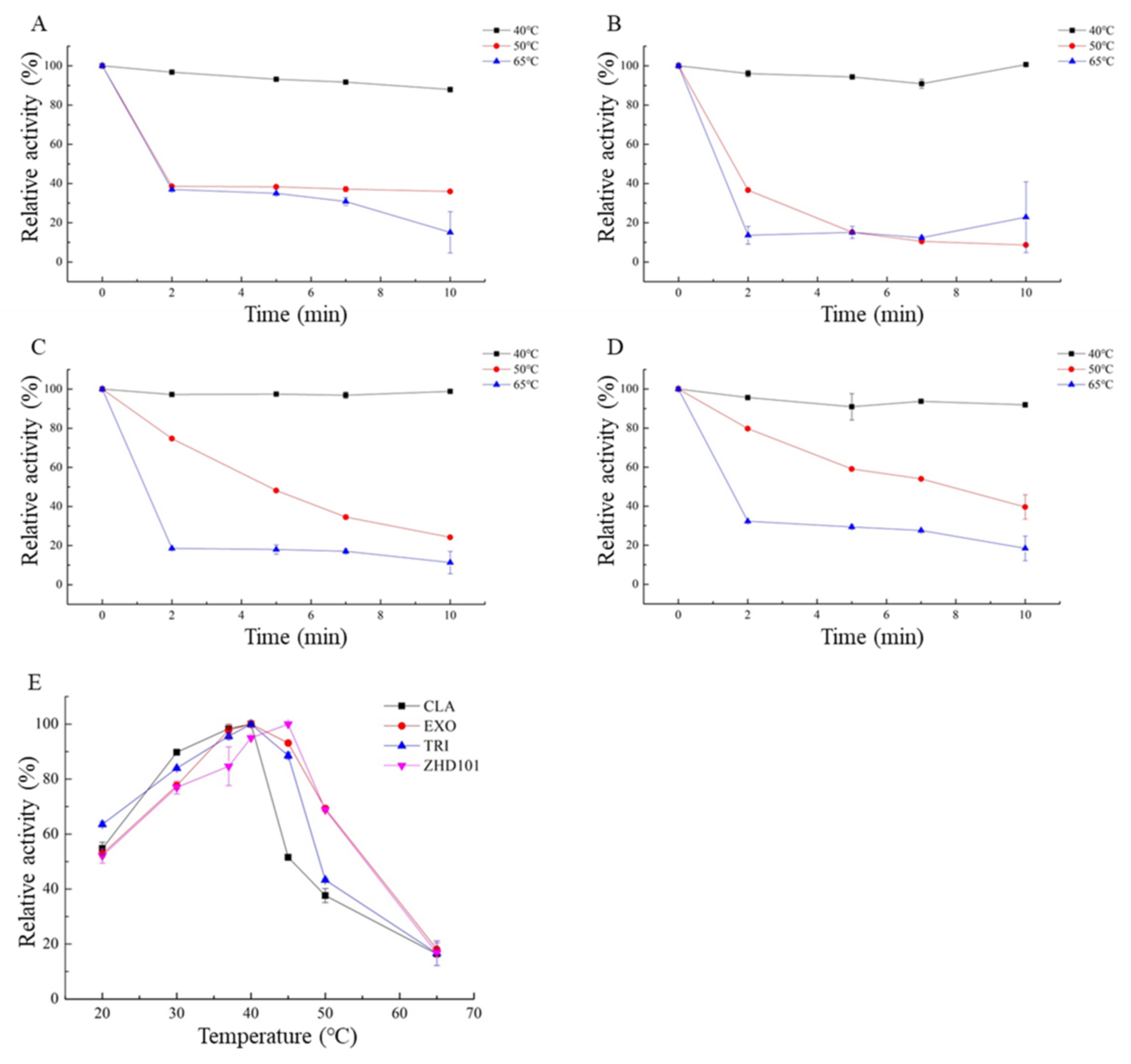
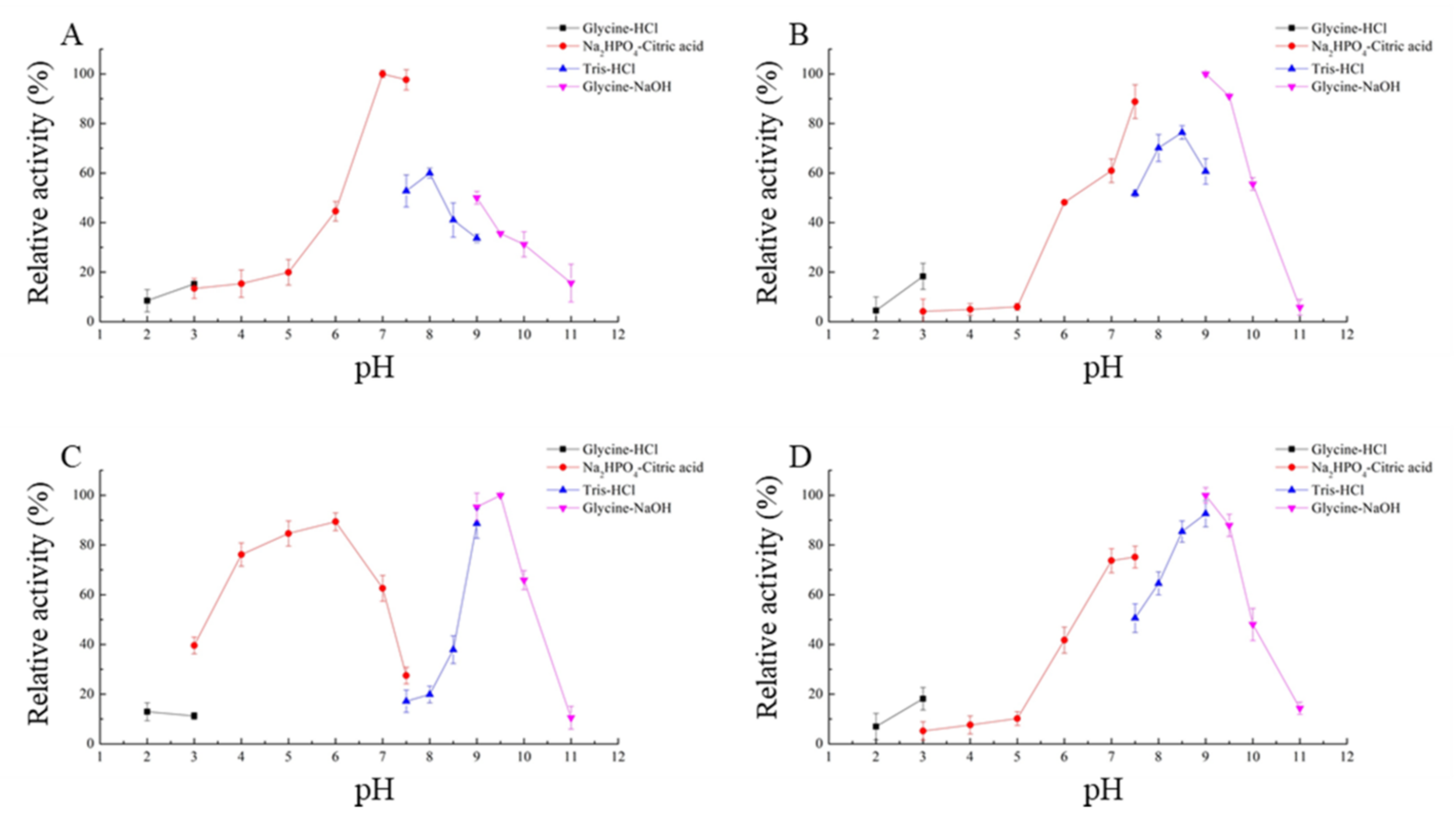
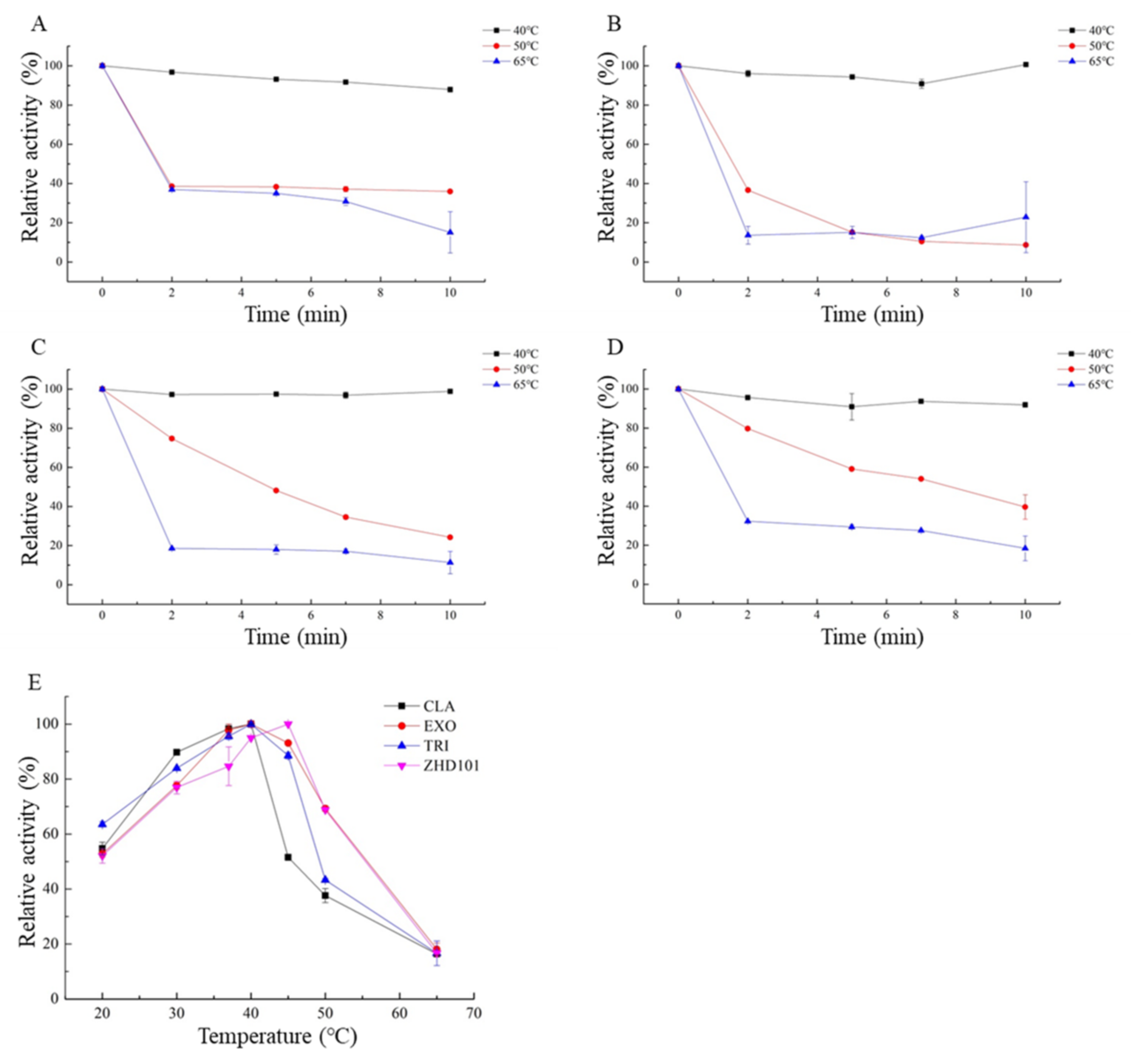
2.3. Enzymatic Properties of the Four ZEN-Degrading Enzymes
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plasmids, Strains, Chemicals, and Medium
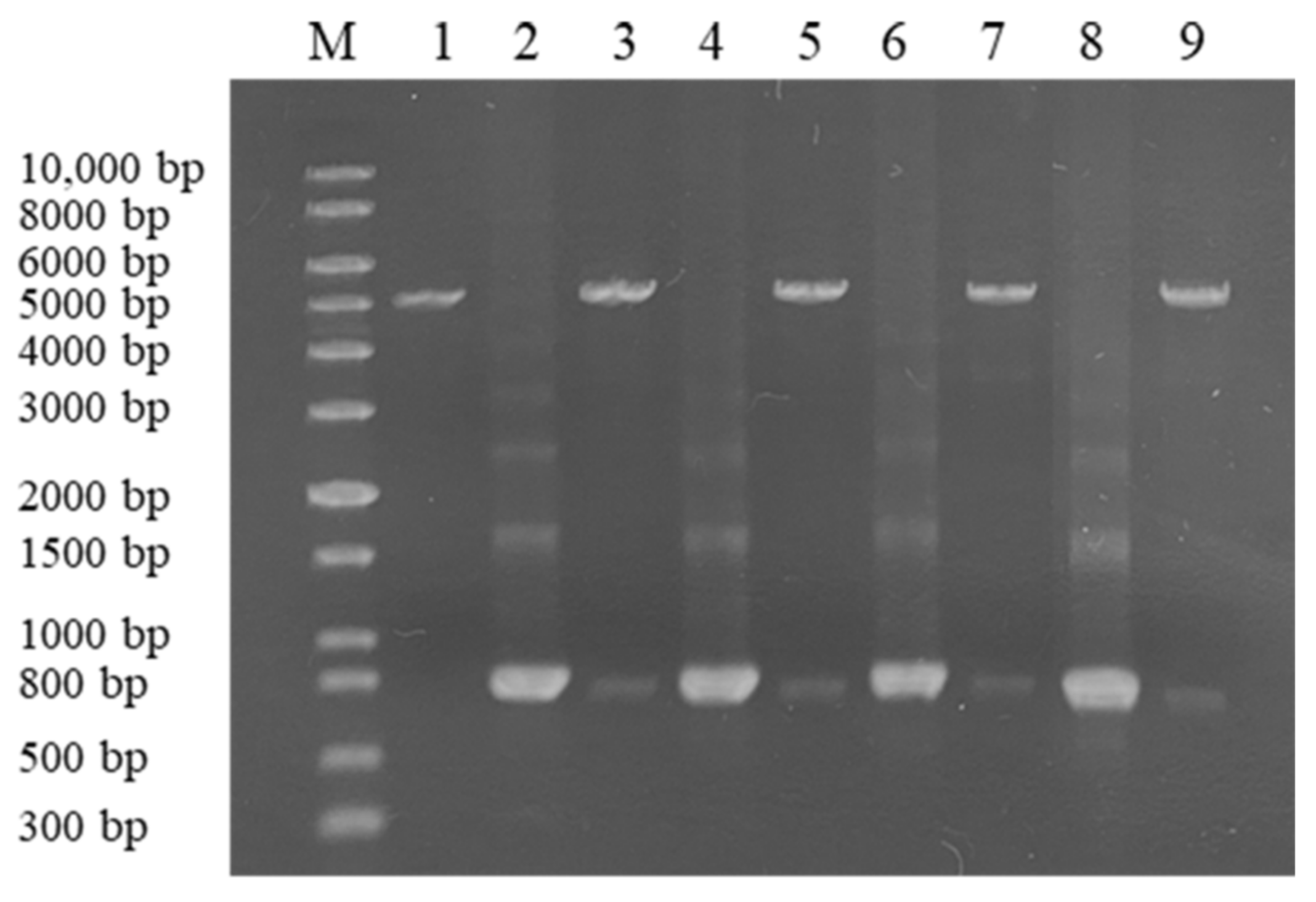
5.2. Cloning of the Four ZEN-Degrading Enzyme Genes and Expression in E. coli
5.3. Enzyme Purification and Molecular Mass Determination
5.4. Enzymatic Substrate Degrading Activity
5.5. Effects of Temperature and pH on the Enzyme Activity and Stability
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stob, M.; Baldwin, R.S.; Tuite, J.; Andrews, F.N.; Gillette, K.G. Solation of an anabolic, uterotrophic compound from corn infected with Gibberella zeae. Nature 1962, 196, 1318. [Google Scholar] [CrossRef] [PubMed]
- Alshannaq, A.; Yu, J.-H. Occurrence, toxicity, and analysis of major mycotoxins in food. Int. J. Environ. 2017, 14, 632. [Google Scholar] [CrossRef] [Green Version]
- Kuiper-Goodman, T.; Scott, P.M.; Watanabe, H. Risk assessment of the mycotoxin zearalenone. Regul. Toxicol. Pharm 1987, 7, 253–306. [Google Scholar] [CrossRef]
- Bulgaru, C.V.; Marin, D.E.; Pistol, G.C.; Taranu, I. Zearalenone and the immune response. Toxins 2021, 13, 248. [Google Scholar] [CrossRef] [PubMed]
- Maaroufi, K.; Chekir, L.; Creppy, E.E.; Ellouz, F.; Bacha, H. Zearalenone induces modifications of haematological and biochemical parameters in rats. Toxicon 1996, 34, 535–540. [Google Scholar] [CrossRef]
- Wang, Y.C.; Deng, J.L.; Xu, S.W.; Peng, X.; Zuo, Z.C.; Cui, H.M.; Wang, Y.; Ren, Z.H. Effects of zearalenone on il-2, il-6, and ifn-γmrna levels in the splenic lymphocytes of chickens. Sci. World J. 2012, 2012, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scudamore, K.A.; Patel, S. Survey for aflatoxins, ochratoxin a, zearalenone and fumonisins in maize imported into the united kingdom. Food Addit. Contam. 2000, 17, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.I.; Almeida, N.G.; Carvalho, K.L.; Gonçalves, G.A.A.; Silva, C.N.; Santos, E.A.; Garcia, J.C.; Vargas, E.A. Co-occurrence of aflatoxins b1, b2, g1and g2, ochratoxin a, zearalenone, deoxynivalenol, and citreoviridin in rice in brazil. Food Addit. Contam. Part A 2012, 29, 694–703. [Google Scholar] [CrossRef]
- Pleadin, J.; Sokolović, M.; Perši, N.; Zadravec, M.; Jaki, V.; Vulić, A. Contamination of maize with deoxynivalenol and zearalenone in croatia. Food Control 2012, 28, 94–98. [Google Scholar] [CrossRef]
- Rai, A.; Dixit, S.; Singh, S.P.; Gautam, N.K.; Das, M.; Tripathi, A. Presence of zearalenone in cereal grains and its exposure risk assessment in indian population. J. Food Sci. 2018, 83, 3126–3133. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Molto, J.C.; Man, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Ropejko, K.; Twarużek, M. Zearalenone and its metabolites—general overview, occurrence, and toxicity. Toxins 2021, 13, 35. [Google Scholar] [CrossRef] [PubMed]
- Mahato, D.K.; Devi, S.; Pandhi, S.; Sharma, B.; Maurya, K.K.; Mishra, S.; Dhawan, K.; Selvakumar, R.; Kamle, M.; Mishra, A.K.; et al. Occurrence, impact on agriculture, human health, and management strategies of zearalenone in food and feed: A review. Toxins 2021, 13, 92. [Google Scholar] [CrossRef] [PubMed]
- Kabak, B.; Dobson, A.D.W.; Var, I. Strategies to prevent mycotoxin contamination of food and animal feed: A review. Crit. Rev. Food Sci. Nutr. 2006, 46, 593–619. [Google Scholar] [CrossRef] [PubMed]
- Calado, T.; Abrunhosa, L.; Cabo Verde, S.; Alté, L.; Venâncio, A.; Fernández-Cruz, M.L. Effect of gamma-radiation on zearalenone—degradation, cytotoxicity and estrogenicity. Foods 2020, 9, 1687. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, Y.; Ji, J.; Wu, H.; Pi, F.; Zhang, Y.; Sun, X. Chemical and toxicological alterations of zearalenone under ozone treatment. Food Addit. Contam. Part A 2018, 36, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Lian, C.; Xi, Y.; Sun, Z.; Zheng, S. Evaluation of nonionic surfactant modified montmorillonite as mycotoxins adsorbent for aflatoxin b1 and zearalenone. J. Colloid Interface Sci. 2018, 518, 48–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalska, K.; Habrowska-Górczyńska, D.E.; Piastowska-Ciesielska, A.W. Zearalenone as an endocrine disruptor in humans. Environ. Toxicol. Pharmacol. 2016, 48, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Qi, L.; Liu, Y.; Wang, R.; Yang, D.; Li, K. Effects of electron beam irradiation on zearalenone and ochratoxin a in naturally contaminated corn and corn quality parameters. Toxins 2017, 9, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, J.; He, B.; Zhang, L.; Li, P.; Zhang, Q.; Ding, X.; Zhang, W. A structure identification and toxicity assessment of the degradation products of aflatoxin b1 in peanut oil under uv irradiation. Toxins 2016, 8, 332. [Google Scholar] [CrossRef] [PubMed]
- Ryu, D.; Hanna, M.A.; Bullerman, L.B. Stability of zearalenone during extrusion of corn grit. J. Food Prot. 1999, 62, 1482–1484. [Google Scholar] [CrossRef]
- Bueno, D.J.; Marco, L.D.; Oliver, G.; Bardón, A. In vitro binding of zearalenone to different adsorbents. J. Food Prot. 2005, 68, 613–615. [Google Scholar] [CrossRef] [PubMed]
- Kriszt, R.; Krifaton, C.; Szoboszlay, S.; Cserháti, M.; Kriszt, B.; Kukolya, J.; Czéh, A.; Fehér-Tóth, S.; Török, L.; Szőke, Z.; et al. A new zearalenone biodegradation strategy using non-pathogenic Rhodococcus pyridinivorans k408 strain. PLoS ONE 2012, 7, e43608. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.Q.; Yang, F.; Yang, P.L.; Liu, J.; Lv, Z.H. Microbial reduction of zearalenone by a new isolated Lysinibacillus sp. Zj-2016-1. World Mycotoxin J. 2018, 11, 571–578. [Google Scholar] [CrossRef]
- Chen, S.-W.; Wang, H.-T.; Shih, W.-Y.; Ciou, Y.-A.; Chang, Y.-Y.; Ananda, L.; Wang, S.-Y.; Hsu, J.-T. Application of zearalenone (zen)-detoxifying bacillus in animal feed decontamination through fermentation. Toxins 2019, 11, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakeya, H.; Takahashi-Ando, N.; KIimura, M.; Onose, R. Biotransformation of the mycotoxin, zearalenone, to a non-estrogenic compound by a fungal strain of Clonostachys sp. Biosci. Biotechnol. Biochem. 2002, 66, 2723–2726. [Google Scholar] [CrossRef] [Green Version]
- Banu, I.; Lupu, A.; Aprodu, I. Degradation of zearalenone by laccase enzyme. Sci. Study Res. 2013, 14, 79–84. [Google Scholar]
- Zheng, Y.; Liu, W.; Chen, C.-C.; Hu, X.; Liu, W.; Ko, T.-P.; Tang, X.; Wei, H.; Huang, J.-W.; Guo, R.-T. Crystal structure of a mycoestrogen-detoxifying lactonase from Rhinocladiella mackenziei: Molecular insight into zhd substrate selectivity. ACS Catal. 2018, 8, 4294–4298. [Google Scholar] [CrossRef]
- De Oliveira Garcia, S.; Sibaja, K.V.M.; Nogueira, W.V.; Feltrin, A.C.P.; Pinheiro, D.F.A.; Cerqueira, M.B.R.; Furlong, E.B.; Garda-Buffon, J. Peroxidase as a simultaneous degradation agent of ochratoxin a and zearalenone applied to model solution and beer. Food Res. Int. 2020, 131, 109039. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Ando, N.; Kimura, M.; Kakeya, H.; Osada, H.; Yamaguchi, I. A novel lactonohydrolase responsible for the detoxification of zearalenone: Enzyme purification and gene cloning. Biochem. J. 2002, 365, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Takahashi-Ando, N.; Ohsato, S.; Shibata, T.; Hamamoto, H.; Yamaguchi, I.; Kimura, M. Metabolism of zearalenone by genetically modified organisms expressing the detoxification gene from Clonostachys rosea. Appl. Environ. Microbiol. 2004, 70, 3239–3245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higa-Nishiyama, A.; Takahashi-Ando, N.; Shimizu, T.; Kudo, T.; Yamaguchi, I.; Kimura, M. A model transgenic cereal plant with detoxification activity for the estrogenic mycotoxin zearalenone. Transgenic Res. 2005, 14, 713–717. [Google Scholar] [CrossRef]
- Takahashi-Ando, N.; Tokai, T.; Hamamoto, H.; Yamaguchi, I.; Kimura, M. Efficient decontamination of zearalenone, the mycotoxin of cereal pathogen, by transgenic yeasts through the expression of a synthetic lactonohydrolase gene. Appl. Microbiol. Biotechnol. 2005, 67, 838–844. [Google Scholar] [CrossRef]
- Xiang, L.; Wang, Q.H.; Zhou, Y.L.; Yin, L.F.; Zhang, G.M.; Ma, Y.H. High-level expression of a zen-detoxifying gene by codon optimization and biobrick in Pichia pastoris. Microbiol. Res. 2016, 193, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.C.; Hsu, T.C.; Cheng, K.C.; Liu, J.R. Expression of the clonostachys rosea lactonohydrolase gene by lactobacillus reuteri to increase its zearalenone-removing ability. Microb. Cell Fact. 2017, 16, 69. [Google Scholar] [CrossRef]
- Yoon, S.H.; Jeong, H.; Kwon, S.K.; Kim, J.F. Genomics, Biological Features, and Biotechnological Applications of Escherichia Coli B: “Is B for Better?!”; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–17. [Google Scholar]
- Wang, M.; Yin, L.; Hu, H.; Selvaraj, J.N.; Zhou, Y.; Zhang, G. Expression, functional analysis and mutation of a novel neutral zearalenone-degrading enzyme. Int. J. Biol. Macromol. 2018, 118, 1284–1292. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, W.; Wu, H.; Zhang, W.; Mu, W. Identification of a potent enzyme for the detoxification of zearalenone. J. Agric. Food Chem. 2020, 68, 376–383. [Google Scholar] [CrossRef] [PubMed]
- Chai, C.; Chang, X.; Wang, N. Cloning of zen-degrading enzyme zhd795 and study on degradation activity. J. Food Sci. Technol. 2018, 36, 28–32. [Google Scholar]
- Cheng, B.; Shi, W.; Luo, J.; Peng, F.; Wan, C.; Wei, H. Cloning of zearalenone-degraded enzyme gene(zen-jjm) and its expression and activity analysis. J. Agric. Biotechnol. 2010, 18, 225–230. [Google Scholar]
- Aiko, V.; Mehta, A. Occurrence, detection and detoxification of mycotoxins. J. Biosci. 2015, 40, 943–954. [Google Scholar] [CrossRef]
- Grenier, B.; Loureiro-Bracarense, A.P.; Leslie, J.F.; Oswald, I.P. Physical and Chemical Methods for Mycotoxin Decontamination in Maize; Wiley: New York, NY, USA, 2014; pp. 116–127. [Google Scholar]
- Wang, J.X.; Xie, Y.L. Review on microbial degradation of zearalenone and aflatoxins. Grain Oil Sci. Technol. 2020, 3, 117–125. [Google Scholar] [CrossRef]
- Wang, N.; Wu, W.W.; Pan, J.W.; Long, M. Detoxification strategies for zearalenone using microorganisms: A review. Microorganisms 2019, 7, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinyiro, S.E.; Wokadala, C.; Xu, D.; Yao, W. Adsorption and degradation of zearalenone by bacillus strains. Folia Microbiol. 2011, 56, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.S.; Yu, D.; Liu, N.; Wu, A. Degrading ochratoxin a and zearalenone mycotoxins using a multifunctional recombinant enzyme. Toxins 2019, 11, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viksoe-Nielsen, A.; Soerensen, B.H. Process for Degrading Zearalenone in a Feed Product Employing Lacasse. European Patent WO 2009/077447 A1, PCT/EP2008/067409, 25 June 2009. [Google Scholar]
- Karlovsky, P.; Suman, M.; Berthiller, F.; De Meester, J.; Eisenbrand, G.; Perrin, I.; Oswald, I.P.; Speijers, G.; Chiodini, A.; Recker, T.; et al. Impact of food processing and detoxification treatments on mycotoxin contamination. Mycotoxin Res. 2016, 32, 179–205. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wu, H.; Tang, Y.; Qiu, L. Cloning, expression of a peroxiredoxin gene from acinetobacter sp. SM04 and characterization of its recombinant protein for zearalenone detoxification. Microbiol. Res. 2012, 167, 121–126. [Google Scholar] [CrossRef]
- Bi, K.; Zhang, W.; Xiao, Z.; Zhang, D. Characterization, expression and application of a zearalenone degrading enzyme from Neurospora crassa. AMB Express 2018, 8, 194. [Google Scholar] [CrossRef] [Green Version]
- Hui, R.; Hu, X.; Liu, W.; Liu, W.; Zheng, Y. Characterization and crystal structure of a novel zearalenone hydrolase from Cladophialophora bantiana. Acta Crystallogr. F 2017, 73, 515–519. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Amplicon Gene | Sequences (5′-3′) | Restriction Site |
|---|---|---|---|
| CLA-F | cla | ATAGGATCCATGGCTCCAGAAAGATTGAG | BamHI |
| CLA-R | CGCAAGCTTTCACAAGTACTTTCTAGTAGTTT | HindIII | |
| EXO-F | exo | ATAGGATCCATGAGAACCAGATCCAACAT | BamHI |
| EXO-R | CGCAAGCTTTCACAAGTACTTTCTAGTTTTTT | HindIII | |
| TRI-F | tri | TCAGGATCCATGAGAATCAGATCCACCAT | BamHI |
| TRI-R | GGCAAGCTTTCACAAGTACTTCTGAGTAAACT | HindIII | |
| ZHD101-F | zhd101 | ATAGGATCCATGAGAACGCGGAGCACGAT | BamHI |
| ZHD101-R | CGCAAGCTTCTACAGATGTTTCTGCGTCGTTT | HindIII |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Liu, X.; Zhang, Y.; Zhang, X.; Huang, H. Cloning and Characterization of Three Novel Enzymes Responsible for the Detoxification of Zearalenone. Toxins 2022, 14, 82. https://doi.org/10.3390/toxins14020082
Zhang Y, Liu X, Zhang Y, Zhang X, Huang H. Cloning and Characterization of Three Novel Enzymes Responsible for the Detoxification of Zearalenone. Toxins. 2022; 14(2):82. https://doi.org/10.3390/toxins14020082
Chicago/Turabian StyleZhang, Yi, Xiaomeng Liu, Yunpeng Zhang, Xiaolin Zhang, and He Huang. 2022. "Cloning and Characterization of Three Novel Enzymes Responsible for the Detoxification of Zearalenone" Toxins 14, no. 2: 82. https://doi.org/10.3390/toxins14020082
APA StyleZhang, Y., Liu, X., Zhang, Y., Zhang, X., & Huang, H. (2022). Cloning and Characterization of Three Novel Enzymes Responsible for the Detoxification of Zearalenone. Toxins, 14(2), 82. https://doi.org/10.3390/toxins14020082