Crystal Structures of Botulinum Neurotoxin Subtypes A4 and A5 Cell Binding Domains in Complex with Receptor Ganglioside


Abstract
:1. Introduction
2. Results and Discussion
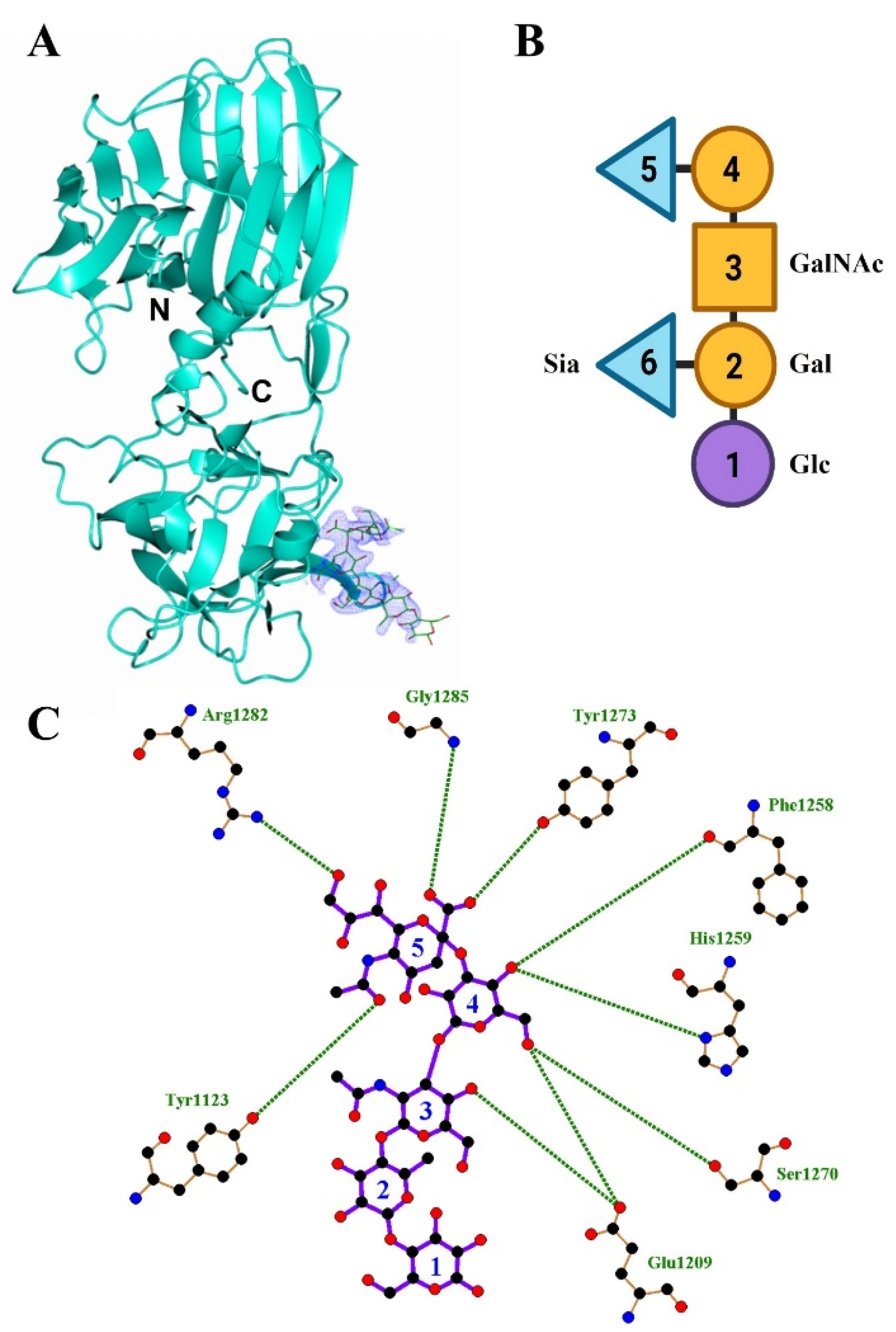
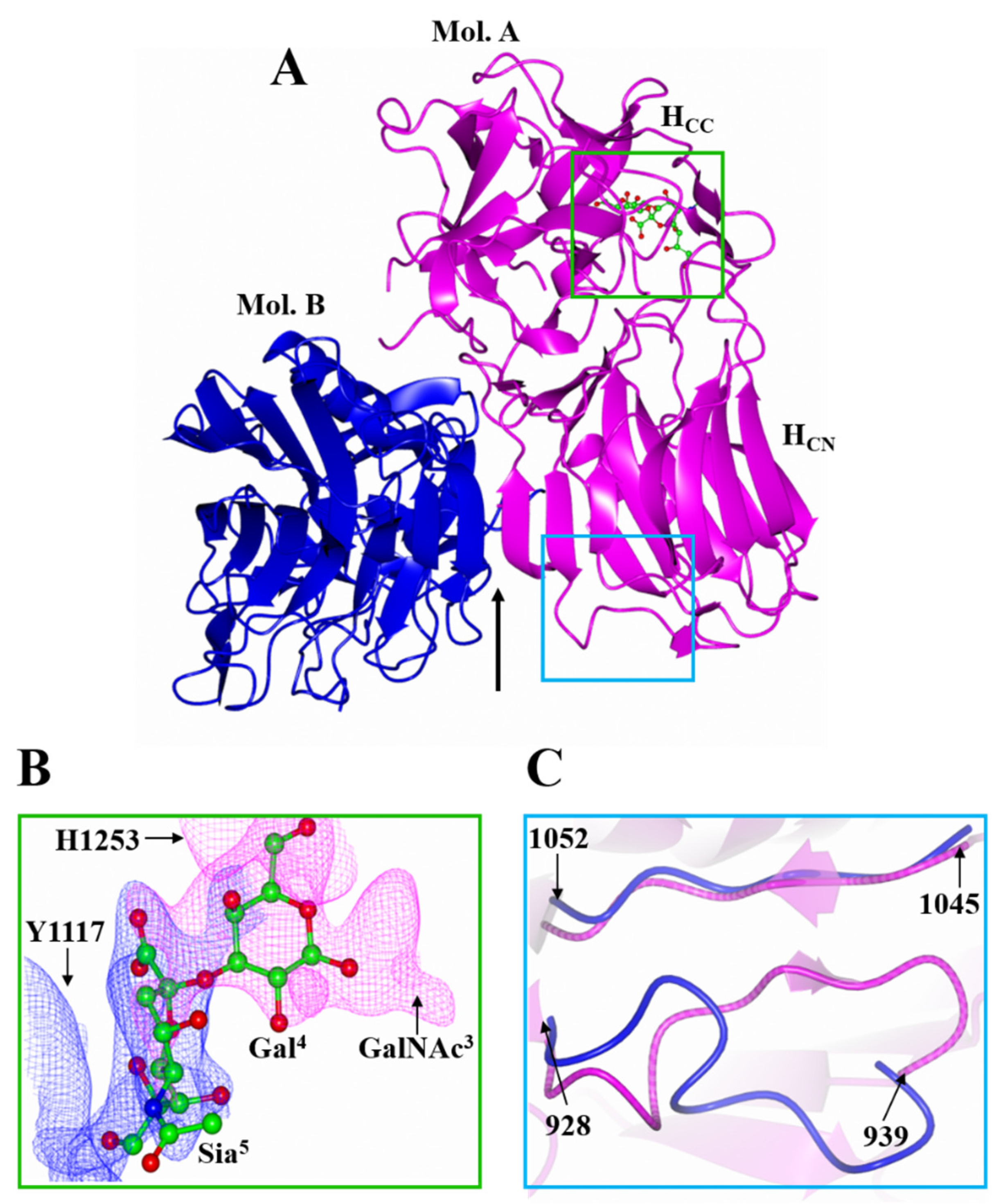
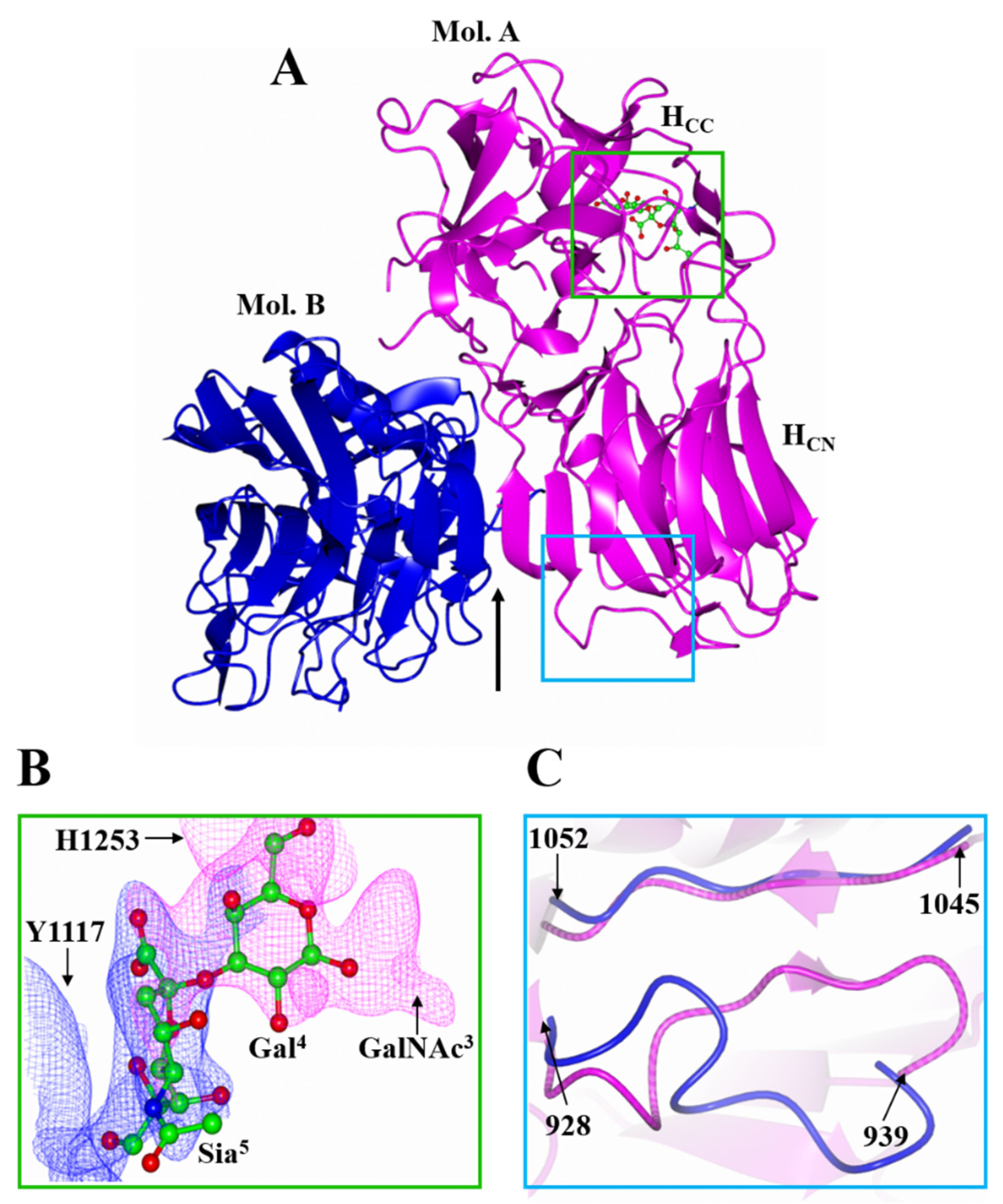
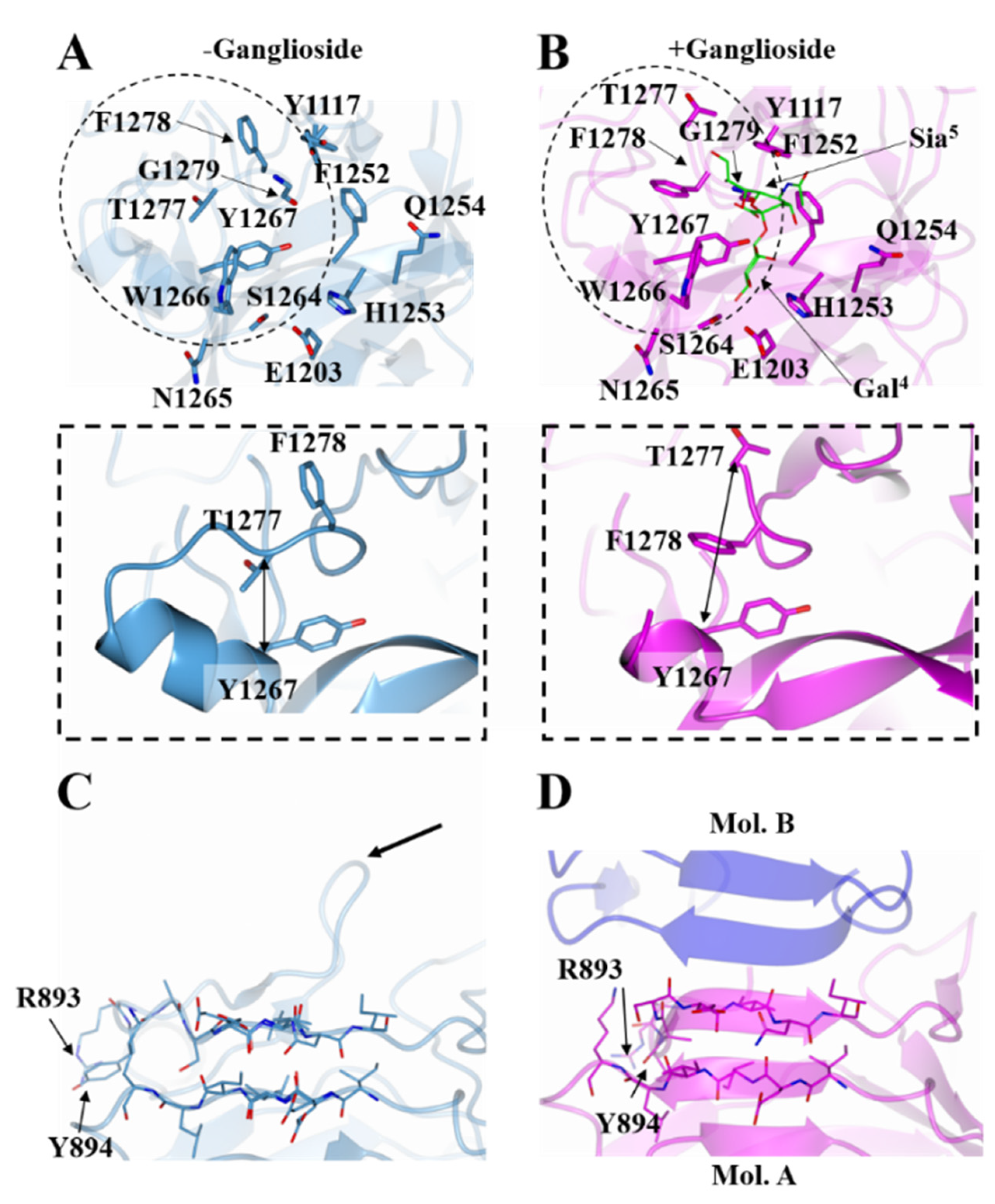
2.1. Structure of HC/A4 in Complex with GD1a Oligosaccharide
2.2. Structure of HC/A5 Co-Crystallised with GM1b Oligosaccharide
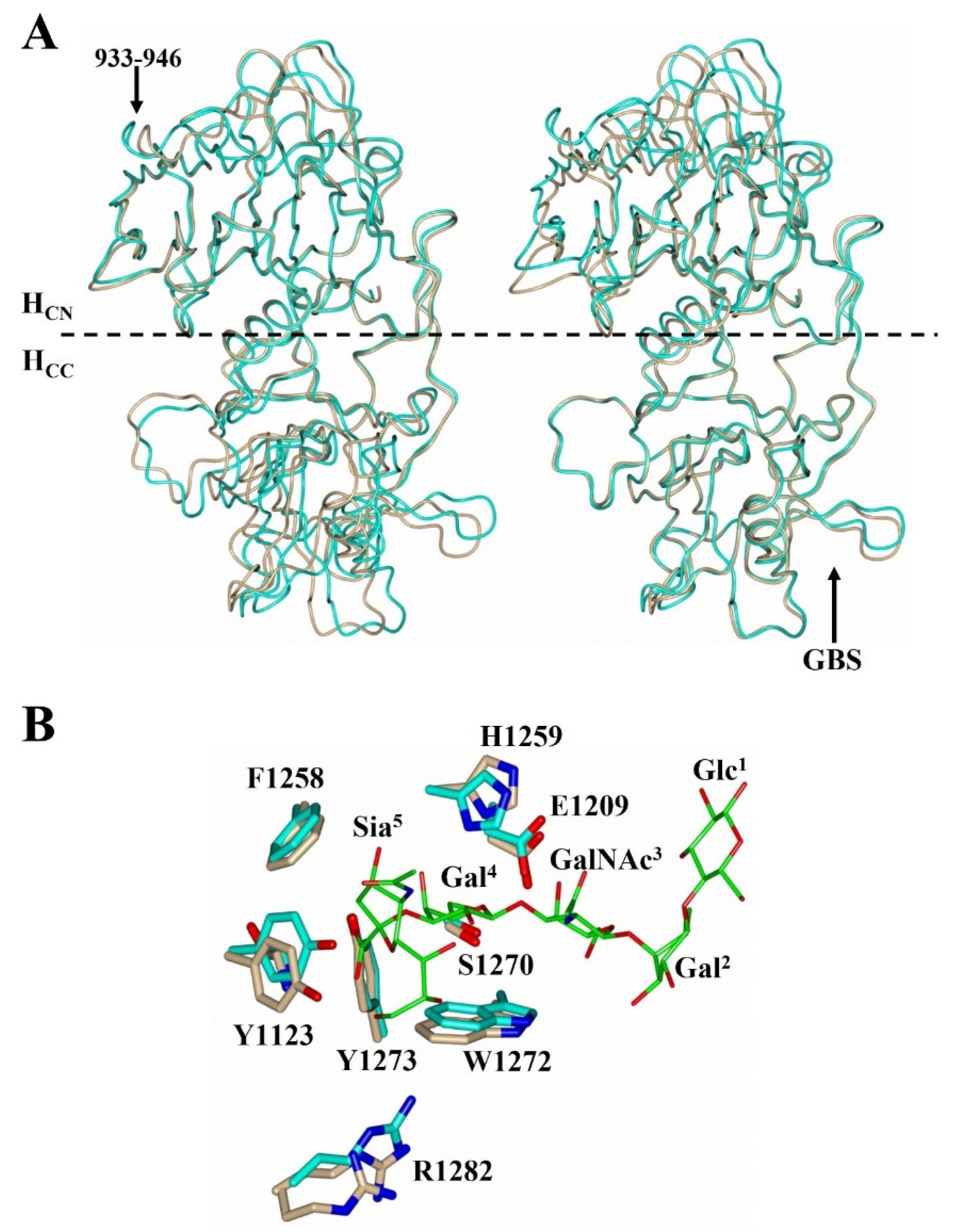
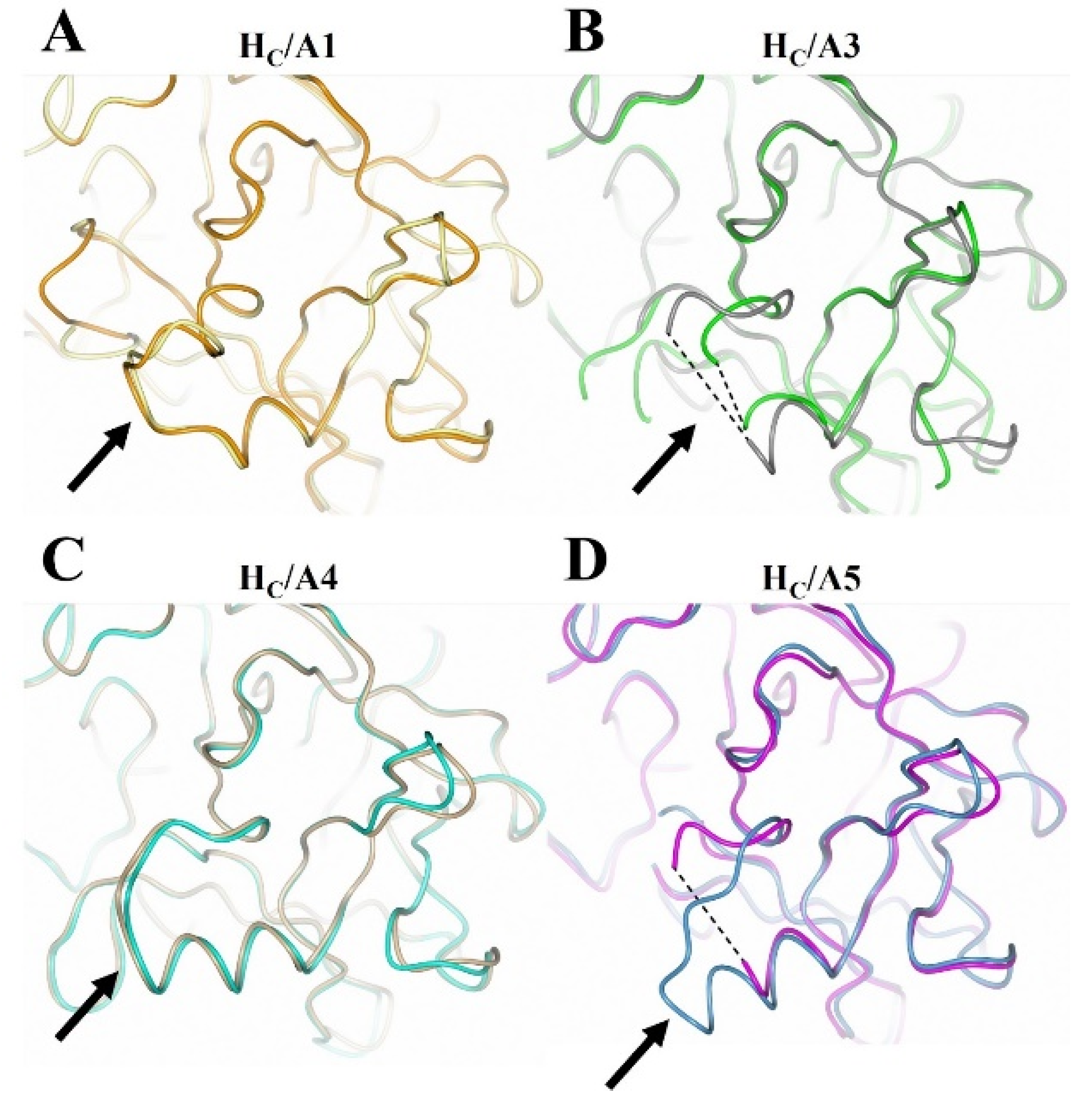
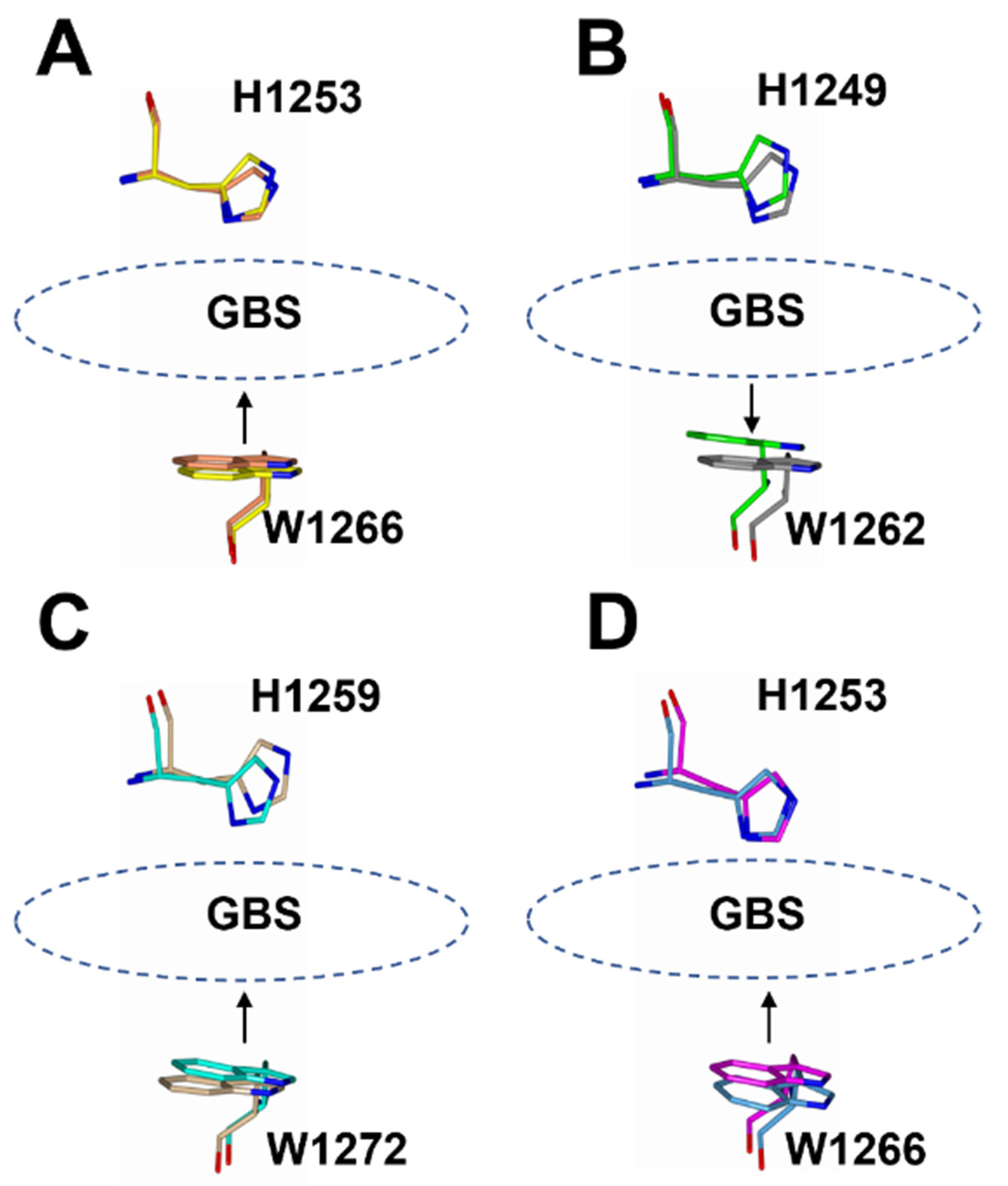
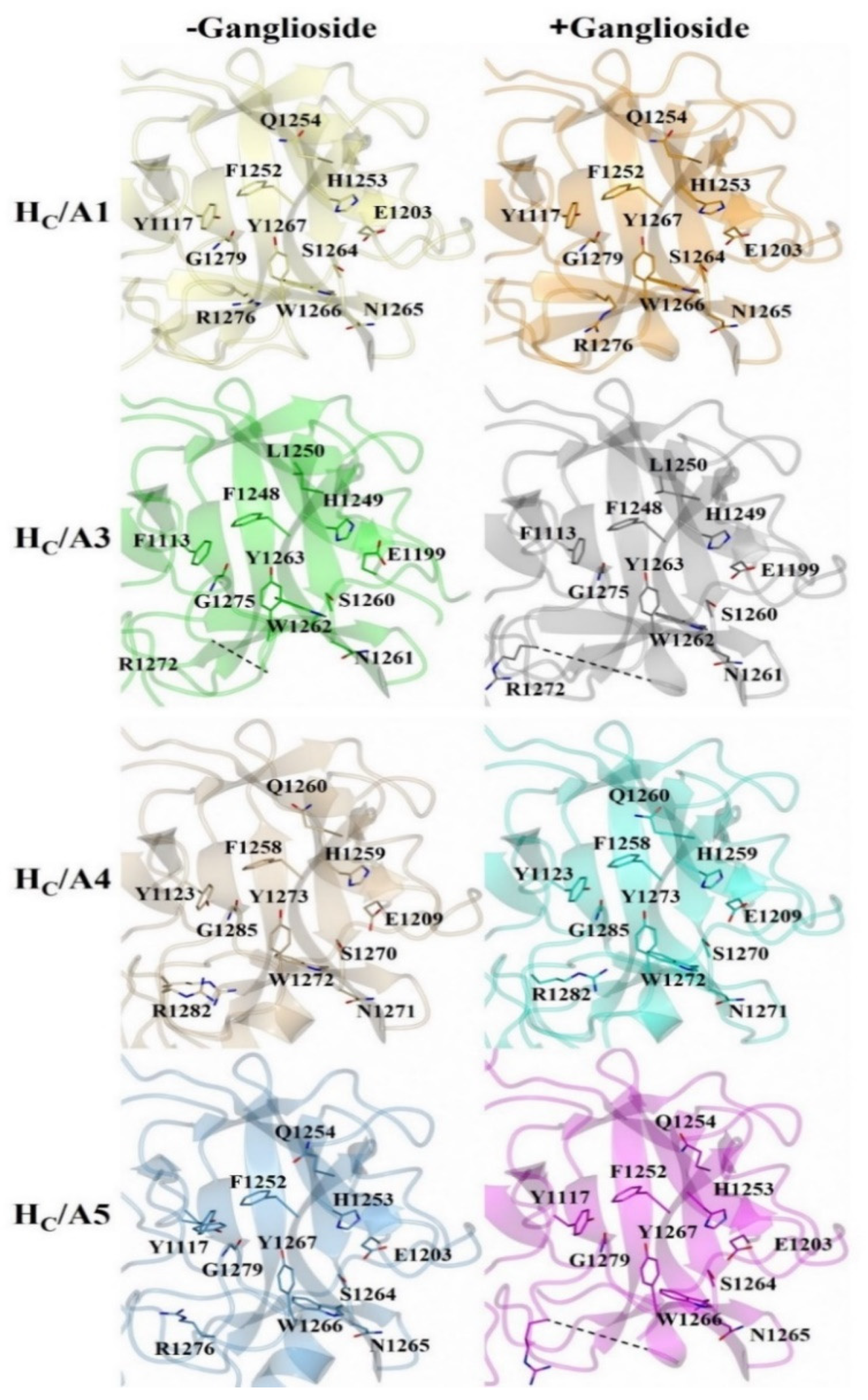
2.3. Structural Variability of HC/A Subtypes at the Ganglioside Binding Site
3. Conclusions
4. Materials and Methods
4.1. Protein Expression and Purification
4.2. Protein Crystallisation
4.3. X-ray Diffraction Data Collection and Structure Determination
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hatheway, C.L. Toxigenic clostridia. Clin. Microbiol. Rev. 1990, 3, 66–98. [Google Scholar] [CrossRef]
- Gill, D.M. Bacterial toxins: A table of lethal amounts. Microbiol. Rev. 1982, 46, 86–94. [Google Scholar] [CrossRef]
- Fonfria, E.; Maignel, J.; Lezmi, S.; Martin, V.; Splevins, A.; Shubber, S.; Kalinichev, M.; Foster, K.; Picaut, P.; Krupp, J. The expanding therapeutic utility of botulinum neurotoxins. Toxins 2018, 10, 208. [Google Scholar] [CrossRef] [Green Version]
- Lacy, D.B.; Tepp, W.; Cohen, A.C.; DasGupta, B.R.; Stevens, R.C. Crystal structure of botulinum neurotoxin type A and implications for toxicity. Nat. Struct. Biol. 1998, 5, 898–902. [Google Scholar] [CrossRef]
- Pellizzari, R.; Rossetto, O.; Schiavo, G.; Montecucco, C. Tetanus and botulinum neurotoxins: Mechanism of action and therapeutic uses. Philos. Trans. R. Soc. B Biol. Sci. 1999, 354, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Foster, K.A. The dual-receptor recognition of botulinum neurotoxins. In Molecular Aspects of Botulinum Neurotoxin; Springer: Berlin, Germany, 2014; pp. 129–150. [Google Scholar]
- Schmid, M.F.; Robinson, J.P.; Dasgupta, B. Direct visualization of botulinum neurotoxin-induced channels in phospholipid vesicles. Nature 1993, 364, 827–830. [Google Scholar] [CrossRef] [PubMed]
- Hoch, D.H.; Romero-Mira, M.; Ehrlich, B.E.; Finkelstein, A.; DasGupta, B.R.; Simpson, L.L. Channels formed by botulinum, tetanus, and diphtheria toxins in planar lipid bilayers: Relevance to translocation of proteins across membranes. Proc. Natl. Acad. Sci. USA 1985, 82, 1692–1696. [Google Scholar] [CrossRef] [Green Version]
- Montal, M. Translocation of botulinum neurotoxin light chain protease by the heavy chain protein-conducting channel. Toxicon 2009, 54, 565–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiavo, G.; Rossetto, O.; Santucci, A.; DasGupta, B.R.; Montecucco, C. Botulinum neurotoxins are zinc proteins. J. Biol. Chem. 1992, 267, 23479–23483. [Google Scholar] [CrossRef]
- Lu, B. The destructive effect of botulinum neurotoxins on the SNARE protein: SNAP-25 and synaptic membrane fusion. PeerJ 2015, 3, e1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanizawa, Y.; Fujisawa, T.; Mochizuki, T.; Kaminuma, E.; Suzuki, Y.; Nakamura, Y.; Tohno, M. Draft genome sequence of Weissella oryzae SG25T, isolated from fermented rice grains. Genome Announc. 2014, 2, 3–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunt, J.; Carter, A.T.; Stringer, S.C.; Peck, M.W. Identification of a novel botulinum neurotoxin gene cluster in Enterococcus. FEBS Lett. 2018, 592, 310–317. [Google Scholar] [CrossRef] [Green Version]
- Contreras, E.; Masuyer, G.; Qureshi, N.; Chawla, S.; Dhillon, H.S.; Lee, H.L.; Chen, J.; Stenmark, P.; Gill, S.S. A neurotoxin that specifically targets Anopheles mosquitoes. Nat. Commun. 2019, 10, 2869. [Google Scholar] [CrossRef] [PubMed]
- Rasetti-Escargueil, C.; Lemichez, E.; Popoff, M.R. Variability of botulinum toxins: Challenges and opportunities for the future. Toxins 2018, 10, 374. [Google Scholar] [CrossRef] [Green Version]
- Hill, K.K.; Smith, T.J.; Helma, C.H.; Ticknor, L.O.; Foley, B.T.; Svensson, R.T.; Brown, J.L.; Johnson, E.A.; Smith, L.A.; Okinaka, R.T.; et al. Genetic diversity among botulinum neurotoxin-producing clostridial strains. J. Bacteriol. 2007, 189, 818–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, J.R.; Liu, S.M.; Acharya, K.R. Variations in the botulinum neurotoxin binding domain and the potential for novel therapeutics. Toxins 2018, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Pellett, S.; Tepp, W.H.; Whitemarsh, R.C.M.M.; Bradshaw, M.; Johnson, E.A. In vivo onset and duration of action varies for botulinum neurotoxin A subtypes 1–5. Toxicon 2015, 107, 37–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Krilich, J.; Pellett, S.; Baudys, J.; Tepp, W.H.; Barr, J.R.; Johnson, E.A.; Kalb, S.R. Comparison of the catalytic properties of the botulinum neurotoxin subtypes A1 and A5. Biochim. Biophys. Acta-Proteins Proteom. 2013, 1834, 2722–2728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arndt, J.W.; Jacobson, M.J.; Abola, E.E.; Forsyth, C.M.; Tepp, W.H.; Marks, J.D.; Johnson, E.A.; Stevens, R.C. A structural perspective of the sequence variability within botulinum neurotoxin subtypes A1–A4. J. Mol. Biol. 2006, 362, 733–742. [Google Scholar] [CrossRef]
- Ledeen, R. Gangliosides of the neuron. Trends Neurosci. 1985, 8, 169–174. [Google Scholar] [CrossRef]
- Hamark, C.; Berntsson, R.P.A.; Masuyer, G.; Henriksson, L.M.; Gustafsson, R.; Stenmark, P.; Widmalm, G. Glycans confer specificity to the recognition of ganglioside receptors by botulinum neurotoxin A. J. Am. Chem. Soc. 2017, 139, 218–230. [Google Scholar] [CrossRef]
- Gregory, K.S.; Liu, S.M.; Acharya, K.R. Crystal structure of botulinum neurotoxin subtype A3 cell binding domain in complex with GD1a co-receptor ganglioside. FEBS Open Biol. 2020, 10, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.R.; Rees, J.; Liu, S.M.; Acharya, K.R. High resolution crystal structures of Clostridium botulinum neurotoxin A3 and A4 binding domains. J. Struct. Biol. 2018, 202, 113–117. [Google Scholar] [CrossRef]
- Davies, J.R.; Britton, A.; Liu, S.M.; Acharya, K.R. High-resolution crystal structures of the botulinum neurotoxin binding domains from subtypes A5 and A6. FEBS Open Biol. 2020, 10, 1474–1481. [Google Scholar] [CrossRef] [PubMed]
- Popoff, M.R.; Connan, C. Absorption and transport of botulinum neurotoxins. In Molecular Aspects of Botulinum Neurotoxin; Springer: Berlin, Germany, 2014; pp. 35–68. [Google Scholar]
- Elez, K.; Bonvin, A.M.J.J.; Vangone, A. Biological vs. crystallographic protein interfaces: An overview of computational approaches for their classification. Crystals 2020, 10, 114. [Google Scholar] [CrossRef] [Green Version]
- Liebschner, D.; Afonine, P.V.; Moriarty, N.W.; Poon, B.K.; Sobolev, O.V.; Terwilliger, T.C.; Adams, P.D. Polder maps: Improving OMIT maps by excluding bulk solvent. Acta Crystallogr. Sect. D Struct. Biol. 2017, 73, 148–157. [Google Scholar] [CrossRef] [Green Version]
- Stenmark, P.; Dupuy, J.; Imamura, A.; Kiso, M.; Stevens, R.C. Crystal structure of botulinum neurotoxin type A in complex with the cell surface co-receptor GT1b-insight into the toxin-neuron interaction. PLoS Pathog. 2008, 4, e1000129. [Google Scholar] [CrossRef] [PubMed]
- Whitemarsh, R.C.M.; Tepp, W.H.; Johnson, E.A.; Pellett, S. Persistence of botulinum neurotoxin a subtypes 1–5 in primary rat spinal cord cells. PLoS ONE 2014, 9, e90252. [Google Scholar] [CrossRef] [Green Version]
- Whitemarsh, R.C.M.; Tepp, W.H.; Bradshaw, M.; Lin, G.; Pier, C.L.; Scherf, J.M.; Johnson, E.A.; Pellett, S. Characterization of botulinum neurotoxin A Subtypes 1 through 5 by investigation of activities in mice, in neuronal cell cultures, and in vitro. Infect. Immun. 2013, 81, 3894–3902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterman, D.G.; Winter, G.; Gildea, R.J.; Parkhurst, J.M.; Brewster, A.S.; Sauter, N.K.; Evans, G. Diffraction-geometry refinement in the DIALS framework. Acta Crystallogr. Sect. D Struct. Biol. 2016, 72, 558–575. [Google Scholar] [CrossRef] [Green Version]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.W.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liebschner, D.; Afonine, P.V.; Baker, M.L.; Bunkoczi, G.; Chen, V.B.; Croll, T.I.; Hintze, B.; Hung, L.W.; Jain, S.; McCoy, A.J.; et al. Macromolecular structure determination using X-rays, neutrons and electrons: Recent developments in Phenix. Acta Crystallogr. Sect. D Struct. Biol. 2019, 75, 861–877. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 2018, 27, 293–315. [Google Scholar] [CrossRef]
- McNicholas, S.; Potterton, E.; Wilson, K.S.; Noble, M.E.M. Presenting your structures: The CCP4mg molecular-graphics software. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 386–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Beamline | I04 | |
| Wavelength used | 0.9795 Å | |
| Protein | HC/A4:GD1a | Hc/A5:GM1b |
| Crystallographic Statistics | ||
| Space group | P61 | P21 |
| Unit cell dimensions: | ||
| a, b, c (Å) | 94.68, 94.68, 181.21 | 44.16, 129.40, 78.05 |
| α, β, γ (°) | 90.00, 90.00, 120.00 | 90.00, 102.96, 90.00 |
| Resolution range (Å) | 90.60–2.30 (2.38–2.30) | 129.40–2.30 (2.49–2.40) |
| Rmerge | 0.269 (1.866) | 0.305 (1.586) |
| Rpim | 0.043 (0.366) | 0.126 (0.746) |
| <I/σ(I)> | 12.1 (1.9) | 4.3 (0.8) |
| CC1/2 | 0.998 (0.644) | 0.982 (0.575) |
| Completeness (%) | 99.7 (97.5) | 100.0 (100.0) |
| No. observed reflections | 1,594,764 (97,629) | 227,781 (22,543) |
| No. unique reflections | 40,672 (3862) | 33,451 (3528) |
| Multiplicity | 39.2 (25.3) | 6.8 (6.4) |
| Refinement Statistics | ||
| Rwork/Rfree | 0.203/0.248 | 0.222/0.262 |
| RMSD bond lengths (Å) | 0.004 | 0.003 |
| RMSD bond angles (°) | 0.68 | 0.55 |
| Ramachandran plot statistics (%): | ||
| Favoured | 95.44 | 95.38 |
| Allowed | 4.56 | 4.34 |
| Outliers | 0.00 | 1.02 |
| Average B-Factors (Å2): | ||
| Protein atoms | 42.28 | 39.10 |
| Solvent atoms | 35.04 | 32.36 |
| GD1a atoms | 72.59 | 70.99 |
| No. Atoms: | 7069 | 6286 |
| Protein | 6696 | 6094 |
| Solvent | 237 | 160 |
| GD1a/GM1b carbohydrates | 136 | 32 |
| PDB code | 7QPT | 7QPU |
| Monosaccharide | H-Bonding Residue (Distance in Å) | |||
|---|---|---|---|---|
| HC/A5:GM1b | HC/A4:GD1a | HC/A3:GD1a | HC/A1:GD1a | |
| Sia6 | N/A | Not modelled | Not modelled | Trp 1266 (3.5) |
| Sia5 | Tyr 1117 (2.8) | Tyr 1123 (2.8) | Tyr 1117 (2.9) | |
| Tyr 1267 (2.7) | Tyr 1273 (2.5) | Tyr 1263 (2.7) | Tyr 1267-H2O (2.5, 3.5) | |
| Gly 1279 (3.2) | Gly 1285 (3.1) | Gly 1275 (2.9) | Gly 1279-H2O Δ (2.6 2.8) | |
| Leu 1250-H2O Δ (2.9, 2.8) | ||||
| Arg 1282 (3.8) | Arg 1276 -H2O Δ (2.8, 2.8) | |||
| Gal4 | Glu 1203 (2.6) | Glu 1209 (2.4) | Glu 1199 (2.7) | Glu 1203 (2.8) |
| Phe 1252 (2.8) | Phe 1258 (2.8) | Phe 1248 (2.5) | Phe 1252 (2.7) | |
| His 1253 (3.1) | His 1259 (2.7) | His 1249 (3.1) | His 1253 (2.7) | |
| Ser 1264 (2.9) | Ser 1270 (2.5) | Ser 1260 (2.7) | Ser 1264 (2.8) | |
| Leu 1250-H2O Δ (2.9, 3.0) | ||||
| GalNAc3 | Not modelled | Glu 1209 (2.6) | Glu 1199 (2.5) | Glu 1203 (2.5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gregory, K.S.; Mojanaga, O.O.; Liu, S.M.; Acharya, K.R. Crystal Structures of Botulinum Neurotoxin Subtypes A4 and A5 Cell Binding Domains in Complex with Receptor Ganglioside. Toxins 2022, 14, 129. https://doi.org/10.3390/toxins14020129
Gregory KS, Mojanaga OO, Liu SM, Acharya KR. Crystal Structures of Botulinum Neurotoxin Subtypes A4 and A5 Cell Binding Domains in Complex with Receptor Ganglioside. Toxins. 2022; 14(2):129. https://doi.org/10.3390/toxins14020129
Chicago/Turabian StyleGregory, Kyle S., Otsile O. Mojanaga, Sai Man Liu, and K. Ravi Acharya. 2022. "Crystal Structures of Botulinum Neurotoxin Subtypes A4 and A5 Cell Binding Domains in Complex with Receptor Ganglioside" Toxins 14, no. 2: 129. https://doi.org/10.3390/toxins14020129
APA StyleGregory, K. S., Mojanaga, O. O., Liu, S. M., & Acharya, K. R. (2022). Crystal Structures of Botulinum Neurotoxin Subtypes A4 and A5 Cell Binding Domains in Complex with Receptor Ganglioside. Toxins, 14(2), 129. https://doi.org/10.3390/toxins14020129