Antiparasitic Effects of Potentially Toxic Beetles (Tenebrionidae and Meloidae) from Steppe Zones


 , ,
, ,  and
and 
Abstract
1. Introduction
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Biological Material
5.2. Beetle Extracts
5.3. Bioassays
5.3.1. Nematicidal Activity
5.3.2. Antiprotozoal Activity
5.3.3. Fungicidal Activity
5.3.4. Ixodicidal Activity
5.4. Chemical Profiles of Organic Extracts
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dettner, K. Defensive compounds and drugs from insects. In Insect Molecular Biology and Ecology; Hoffman, K.H., Ed.; Taylor & Francis: Boca Raton, FL, USA, 2014; pp. 39–93. [Google Scholar]
- Dossey, A.T. Insects and their chemical weaponry: New potential for drug discovery. Nat. Prod. Rep. 2010, 27, 1737–1757. [Google Scholar] [CrossRef]
- Deyrup, S.T.; Stagnitti, N.C.; Perpetua, M.J.; Wong-Deyrup, S.W. Drug discovery insights from medicinal beetles in traditional chinese medicine. Biomol. Ther. 2021, 29, 105–126. [Google Scholar] [CrossRef] [PubMed]
- Nishida, R. Sequestration of defensive substances from plants by Lepidoptera. Annu. Rev. Entomol. 2002, 47, 57–92. [Google Scholar] [CrossRef] [PubMed]
- Eisner, T.; Goetz, M.A.; Hill, D.E.; Smedley, S.R.; Meinwald, J. Firefly ‘‘femmes fatales’’ acquire defensive steroids (lucibufagins) from their firefly prey. Proc. Natl. Acad. Sci. USA 1997, 94, 9723–9728. [Google Scholar] [CrossRef]
- Hernandez-Alvarado, J.; Zaragoza-Bastida, A.; Lopez-Rodriguez, G.; Pelaez-Acero, A.; Olmedo-Juarez, A.; Rivero-Perez, N. Antibacterial and antihelmintic activity of secondary metabolites of plants: Approach in Veterinary Medicine. Abanico Vet. 2018, 8, 14–27. [Google Scholar]
- Singh, N.; Kumar, M.; Singh, R.K. Leishmaniasis: Current status of available drugs and new potential drug targets. Asian Pac. J. Trop. Med. 2012, 5, 485–497. [Google Scholar] [CrossRef]
- Thao, N.P.; No, J.H.; Luyen, B.T.T.; Gyongseon, Y.; Byun, S.Y.; Goo, J.; Kim, K.T.; Cuong, N.X.; Nam, N.H.; Minh, C.V.; et al. Secondary Metabolites from Vietnamese Marine Invertebrates with Activity against Trypanosoma brucei and T. cruzi. Molecules 2014, 19, 7869–7880. [Google Scholar] [CrossRef]
- Oliveira, M.; Barreira, L.; Gangadhar, K.N.; Rodrigues, M.J.; Santos, T.; Varela, J.; Custodio, L. Natural products from marine invertebrates against Leishmania parasites: A comprehensive review. Phytochem. Rev. 2016, 15, 663–697. [Google Scholar] [CrossRef]
- Rocha, J.; Peixe, L.; Gomes, N.C.M.; Calado, R. Cnidarians as a source of new marine bioactive compounds-An overview of the last decade and future steps for bioprospecting. Mar. Drugs 2011, 9, 1860–1886. [Google Scholar] [CrossRef]
- Mendoza Meza, D.L.; Sánchez Catalán, K.; Saavedra Ahumada, S. Caracterización bioquímica de extractos acuosos de Ulomoides dermestoides (Coleoptera, Tenebrionidae): Exploración de la inhibición dual de la lipoxigenasa (15-lox) y ciclooxigenasas (Cox-1 y Cox-2). Biosalud 2016, 15, 37–54. [Google Scholar] [CrossRef]
- Whitman, D.W.; Andres, M.F.; Martinez-Diaz, R.A.; Ibanez-Escribano, A.; Olmeda, A.S.; Gonzalez-Coloma, A. Antiparasitic properties of cantharidin and the blister beetle Berberomeloe majalis (Coleoptera: Meloidae). Toxins 2019, 11, 234. [Google Scholar] [CrossRef]
- Chacón-Castro, R.E.; Villalba-Velásquez, V.; Moreira-González, Y. Descripción anatómica de los órganos genitales y glándulas secretoras del escarabajo Ulomoides dermestoides (Fairmere 1893) (Coleoptera: Tenebrionidae). Tecnol. Marcha 2009, 22, 45–65. [Google Scholar]
- Singer, M.; Mace, K.; Bernays, E. Self-medication as adaptive plasticity: Increased ingestion of plant toxins by parasitized caterpillars. PLoS ONE 2009, 4, e4796. [Google Scholar] [CrossRef]
- Huffman, M.A. Animal self-medication and ethno-medicine: Exploration and exploitation of the medicinal properties of plants. Proc. Nutr. Soc. 2003, 62, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Bravo, C.; Bautista, L.M.; Garcia-Paris, M.; Blanco, G.; Alonso, J.C. Males of a strongly polygynous species consume more poisonous food than females. PLoS ONE 2014, 9, e111057. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.P.; Dong, J.; Cai, H.; Wang, W. Cantharidin as an antitumor agent: A retrospective review. Curr. Med. Chem. 2013, 20, 159–166. [Google Scholar] [CrossRef]
- Percino-Daniel, N.; Buckley, D.; Garcia-Paris, M. Pharmacological properties of blister beetles (Coleoptera: Meloidae) promoted their integration into the cultural heritage of native rural Spain as inferred by vernacular names diversity, traditions, and mitochondrial DNA. J. Ethnopharmacol. 2013, 147, 570–583. [Google Scholar] [CrossRef] [PubMed]
- Bravo, C.; Mas-Peinado, P.; Bautista, L.M.; Blanco, G.; Alonso, J.C.; Garcia-Paris, M. Cantharidin is conserved across phylogeographic lineages and present in both morphs of Iberian Berberomeloe blister beetles (Coleoptera, Meloidae). Zool. J. Linn. Soc. 2017, 180, 790–804. [Google Scholar] [CrossRef]
- Sanchez-Barbudo, I.S.; Camarero, P.R.; Garcia-Montijano, M.; Mateo, R. Possible cantharidin poisoning of a great bustard (Otis tarda). Toxicon 2012, 59, 100–103. [Google Scholar] [CrossRef]
- Ghoneim, K. Cantharidin toxicosis to animal and human in the World: A review. Standard Res. J. Toxicol. Environ. Health Sci. 2013, 1, 1–22. [Google Scholar]
- Faragó, S. Spectrum of plant and animal diet of European Great Bustard (Otis tarda tarda)—An overview. Ornis Hung. 2019, 27, 62–84. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, Y.-H.; Luo, Q.; Hu, Y.; Lu, Q.; Yu, W.-X.; Chen, J.-H.; Wang, Y.-J.; Wang, M.-L.; Zhao, Y.; et al. Content and composition analysis of polysaccharides from Blaps rynchopetera and its macrophage phagocytic activity. Open Chem. 2021, 19, 338–346. [Google Scholar] [CrossRef]
- Gunbilig, D.; Boland, W. Defensive agents of Blaps femoralis, a traditional Mongolian medicinal insect. Sci. Pharm. 2009, 77, 597–604. [Google Scholar] [CrossRef]
- Afonso, C.M.M.; Paulmier, I.; Pinto, M.M.M.; Maciel, M.L.; Clément, J.L. Feromonas sexuais de Megachile rotundata: Síntese de análogos structurais. In Proceedings of the 13° Encontro Anual a Sociedade Portuguesa de Química, Lisboa, Portugal, 29 January–1 February 1992. [Google Scholar]
- Moreno-Morales, S.; Crescente, O.; Herrera-Malaver, B.; Hernández, J.V.; Munñoz, M.; Oliveros, M.J. Composición y actividad biológica de los extractos de Ulomoides dermestoides (Tenebrionidae) procesados bajo diferentes condiciones en Cumaná, estado Sucre. FACSALUD-UNEMI 2019, 3, 3–16. [Google Scholar] [CrossRef]
- Howard, R.W.; Blomquist, G.J. Ecological, behavioral, andbiochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef] [PubMed]
- Peeters, C.; Monnin, T.; Malosse, C. Cuticular hydrocarbons correlated with reproductive status in a queenless ant. Proc. R. Soc. B Biol. Sci. 1999, 266, 1323–1327. [Google Scholar] [CrossRef]
- Ginzel, M.D.; Blomquist, G.J.; Millar, J.G.; Hanks, L.M. Role of contact pheromones in mate recognition in Xylotrechus colonus. J. Chem. Ecol. 2003, 29, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R.; Carlson, D.A. cuticular hydrocarbons of the TseTse-flies Glossina morsitans morsitans, G. austeni and G. pallidipes. Insect Biochem. 1986, 16, 403–416. [Google Scholar] [CrossRef]
- Gavris, I.; Bodoki, E.; Verite, P.; Oprean, R. Assessment of perfume ingredients with aphrodisiac potential by gas chromatography—Mass spectrometry. Farmacia 2015, 63, 760–764. [Google Scholar]
- Nikbakhtzadeh, M.R.; Tirgari, S. Cantharidin component of Iranian blister beetles (Col: Meloidae) and their differences between Iranian and exotic species. Iran. J. Public Health 2002, 31, 113–117. [Google Scholar]
- Vivekananthan, T. Identification and isolation of cantharidin and other important in defensive secretion of blister beetle Mylabris pustulata (Thunberg) (Coleoptera: Meloidae) from Tamil Nadu, India. Int. J. Curr. Res. 2014, 3, 232–236. [Google Scholar]
- Nikbakhtzadeh, M.R.; Dettner, K.; Boland, W.; Gaede, G.; Doetterl, S. Intraspecific transfer of cantharidin within selected members of the family Meloidae (Insecta: Coleoptera). J. Insect Physiol. 2007, 53, 890–899. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, C.A.; Howard, R.W.; Oneill, K.M.; Schmidt, J.O. chemistry of male mandibular gland secretions of Philanthus-basilaris cresson and Philanthus-bicinctus (mickel) (hymenoptera, sphecidae). J. Chem. Ecol. 1987, 13, 227–235. [Google Scholar] [CrossRef]
- Akin-Osanaiye, C.B.; Gabriel, A.F.; Alebiosu, R.A. Characterization and antimicrobial screening of ethyl oleate isolated from Phyllanthus amarus (Schum and Thonn). Ann. Biol. Res. 2011, 2, 298–305. [Google Scholar]
- Chen, Y.; Bertrand, C.; Dai, G.; Yuan, J. Biochemical mechanisms of acaricidal activity of 2,4-di-tert-butylphenol and ethyl oleate against the carmine spider mite Tetranychus cinnabarinus. J. Pest Sci. 2018, 91, 405–419. [Google Scholar] [CrossRef]
- Perelman, B.; Chikatunov, V. Intoxication of young crocodiles in captivity due to the ingestión of darkling beetles Blaps nitens Laportei Ardoin (Coleoptera; Tenebrionidae). Isr. J. Vet. Med. 2010, 65, 100–102. [Google Scholar]
- Huffman, M.A. Current evidence for self-medication in primates: A multidisciplinary perspective. In Yearbook of Physical Anthropology, Vol 40—1997; Steegmann, A.T., Ed.; Wiley-Liss, Inc.: Hoboken, NJ, USA, 1997; Volume 40, pp. 171–200. [Google Scholar]
- Masello, J.F.; Martinez, J.; Calderon, L.; Wink, M.; Quillfeldt, P.; Sanz, V.; Theuerkauf, J.; Ortiz-Catedral, L.; Berkunsky, I.; Brunton, D.; et al. Can the intake of antiparasitic secondary metabolites explain the low prevalence of hemoparasites among wild Psittaciformes? Parasit. Vect. 2018, 11, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Gilardi, J.D.; Duffey, S.S.; Munn, C.A.; Tell, L.A. Biochemical functions of geophagy in parrots: Detoxification of dietary toxins and cytoprotective effects. J. Chem. Ecol. 1999, 25, 897–922. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D.; Shaw, R. Sheep self-medicate when challenged with illness-inducing foods. Anim. Behav. 2006, 71, 1131–1139. [Google Scholar] [CrossRef]
- Krishnamani, R.; Mahaney, W.C. Geophagy among primates: Adaptive significance and ecological consequences. Anim. Behav. 2000, 59, 899–915. [Google Scholar] [CrossRef]
- Milan, N.F.; Kacsoh, B.Z.; Schlenke, T.A. Alcohol consumption as self-medication against blood-borne parasites in the Fruit Fly. Curr. Biol. 2012, 22, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Chapuisat, M.; Oppliger, A.; Magliano, P.; Christe, P. Wood ants use resin to protect themselves against pathogens. Proc. R. Soc. B Biol. Sci. 2007, 274, 2013–2017. [Google Scholar] [CrossRef] [PubMed]
- Abbott, J. Self-medication in insects: Current evidence and future perspectives. Ecol. Entomol. 2014, 39, 273–280. [Google Scholar] [CrossRef]
- Schildknecht, H.; Holoubek, K.; Weis, K.H.; Krämer, H. Defensive substances of the arthropods, their isolation and identification. Angew. Chem. Int. 1964, 3, 73–82. [Google Scholar] [CrossRef]
- Tschinkel, W.R. A comparative study of the chemical defensive system of tenebrionid beetles: Chemistry of the secretions. J. Insect Physiol. 1975, 21, 753–783. [Google Scholar] [CrossRef]
- Ikan, R.; Cohen, E.; Shulov, A. benzoquinones and hydroquinones in defence secretions of Blaps sulcata and Blaps wiedemanni. J. Insect Physiol. 1970, 16, 2201–2206. [Google Scholar] [CrossRef]
- Peschke, K.; Eisner, T. Defensive secretion of the tenebrionid beetle, Blaps mucronata: Physical and chemical determinants of effectiveness. J. Comp. Physiol. A 1987, 161, 377–388. [Google Scholar] [CrossRef]
- Morales, M.B.; Martín, C.A. Otis tarda great bustard. BWP Update 2002, 4, 217–232. [Google Scholar]
- Glutz, U.N.; Bauer, K.M.; Bezzel, E. Galliformes und gruiformes. In Handbuch der Vögel Mitteleuropas; Bezzel, E., Glutz, U.N., Bauer, K.M., Eds.; Aula-Verlag: Wiesbaden, Germany, 1994; Volume 4, pp. 566–607. [Google Scholar]
- García-Montijano, M.; Tébar, A.M.; Barreiro, B.; Rodriguez, P.; Alonso, J.C.; Martín, C.A.; Magaña, M.; Alonso, J.A.; Montesinos, A.; Luaces, I. Postmortem findings in wild great bustards (Otis tarda) from Spain: A clinical approach. In Proceedings of the 4th Scientific Meeting EAZWV Joint with the Annual Meeting of the European Wildlife Disease Association (EWDA), Heidelberg, Germany, 8–12 May 2002; p. 6. [Google Scholar]
- Bennett, J.W. An Overview of the Genus Aspergillus. In Aspergillus: Molecular Biology and Genomics; Machida, M., Gomi, K., Eds.; Caister Academic Press: Wymondham, UK, 2010; pp. 1–17. [Google Scholar]
- Escobar, C.; Barcala, M.; Cabrera, J.; Fenoll, C. Overview of Root-Knot Nematodes and Giant Cells. In Plant Nematode Interactions: A View on Compatible Interrelationships; Escobar, C., Fenoll, C., Eds.; Advances in Botanical Research; Academic Press Ltd-Elsevier Science Ltd.: London, UK, 2015; Volume 73, pp. 1–32. [Google Scholar]
- Julio, L.F.; Barrero, A.F.; Mar Herrador del Pino, M.; Arteaga, J.F.; Burillo, J.; Andres, M.F.; Diaz, C.E.; Gonzalez-Coloma, A. Phytotoxic and Nematicidal Components of Lavandula luisieri. J. Nat. Prod. 2016, 79, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Santos-Silva, M.M.; Beati, L.; Santos, A.S.; De Sousa, R.; Nuncio, M.S.; Melo, P.; Santos-Reis, M.; Fonseca, C.; Formosinho, P.; Vilela, C.; et al. The hard-tick fauna of mainland Portugal (Acari: Ixodidae): An update on geographical distribution and known associations with hosts and pathogens. Exp. Appl. Acarol. 2011, 55, 85–121. [Google Scholar] [CrossRef] [PubMed]
- Valcarcel, F.; Gonzalez, J.; Jaime, J.M.T.; Olmeda, A.S. Long term study of ixodid ticks feeding on red deer (Cervus elaphus) in a meso-Mediterranean climate. Exp. Appl. Acarol. 2016, 69, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.; Bilic, I.; Liebhart, D.; Hess, M. Trichomonads in birds—A review. Parasitology 2014, 141, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Andres, M.F.; Gonzalez-Coloma, A.; Sanz, J.; Burillo, J.; Sainz, P. Nematicidal activity of essential oils: A review. Phytochem. Rev. 2012, 11, 371–390. [Google Scholar] [CrossRef]
- Schneider-Orelli, O. Entomologisches Praktikum: Einführung in die Land- und Forstwirtschaftliche Insektenkunde, 2nd ed.; Sauerländer & Co: Aarau, Switzerland, 1947; p. 237. [Google Scholar]
- Puntener, W. Manual for Field Trials in Plant Protection, 2nd ed.; Ciba-Geigy Ltd.: Basle, Switzerland, 1981. [Google Scholar]
- Ruiz-Vasquez, L.; Olmeda, A.S.; Zuniga, G.; Villarroel, L.; Echeverri, L.F.; Gonzalez-Coloma, A.; Reina, M. Insect Antifeedant and Ixodicidal Compounds from Senecio adenotrichius. Chem. Biodivers. 2017, 14, 9. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Family | Species | N° | Fresh Weight (g) | Extract Weight (mg) | Yield (%) |
|---|---|---|---|---|---|
| Tenebrionidae | Tentyria peiroleri | 33 | 4.96 | 100.70 | 2.03 |
| Scaurus uncinus | 8 | 1.45 | 140.90 | 9.72 | |
| Blaps lethifera | 3 | 2.43 | 537.90 | 22.14 | |
| Meloidae | Mylabris quadripunctata | 24 | 2.74 | 78.50 | 2.87 |
| Coleoptera | µg/µL | Mortality a % | Lethal Concentrations b | ||
|---|---|---|---|---|---|
| Family | Species | LC50 µg/µL | LC90 µg/µL | ||
| Tenebrionidae | Tentyria peiroleri | 1.00 | 2.00 ± 1.07 | ||
| Scaurus uncinus | 1.00 | 0.00 | |||
| Blaps lethifera | 1.00 | 0.71 ± 1.03 | |||
| Meloidae | Mylabris quadripunctata | 1.00 | 97.00 ± 1.00 | 0.35 (0.33–0.37) | 0.65 (0.61–0.69) |
| 0.50 | 81.33 ± 1.72 | ||||
| 0.25 | 49.00 ± 4.00 | ||||
| 0.12 | 7.26 ± 3.38 | ||||
| 0.06 | 2.70 ± 1.66 | ||||
| Coleoptera | % Germination Inhibition Aspergillus niger a | % Mortality Hyalomma lusitanicum b | |
|---|---|---|---|
| Family | Species | ||
| Tenebrionidae | Tentyria peiroleri | 9.52 ± 8.46 | 5.08 ± 2.89 |
| Scaurus uncinus | 21.21 ± 8.69 | 11.67 ± 1.67 | |
| Blaps lethifera | 8.65 ± 5.64 | 7.41 ± 3.73 | |
| Meloidae | Mylabris quadripunctata | 0.00 ± 0.00 | 3.70 ± 1.91 |
| Retention Time (min) | Area (%) | Compounds | |||
|---|---|---|---|---|---|
| Mylabris quadripunctata | Tentyria peiroleri | Scaurus uncinus | Blaps lethifera | ||
| 3.06 | - | - | - | 13.34 | 1-Tridecanol |
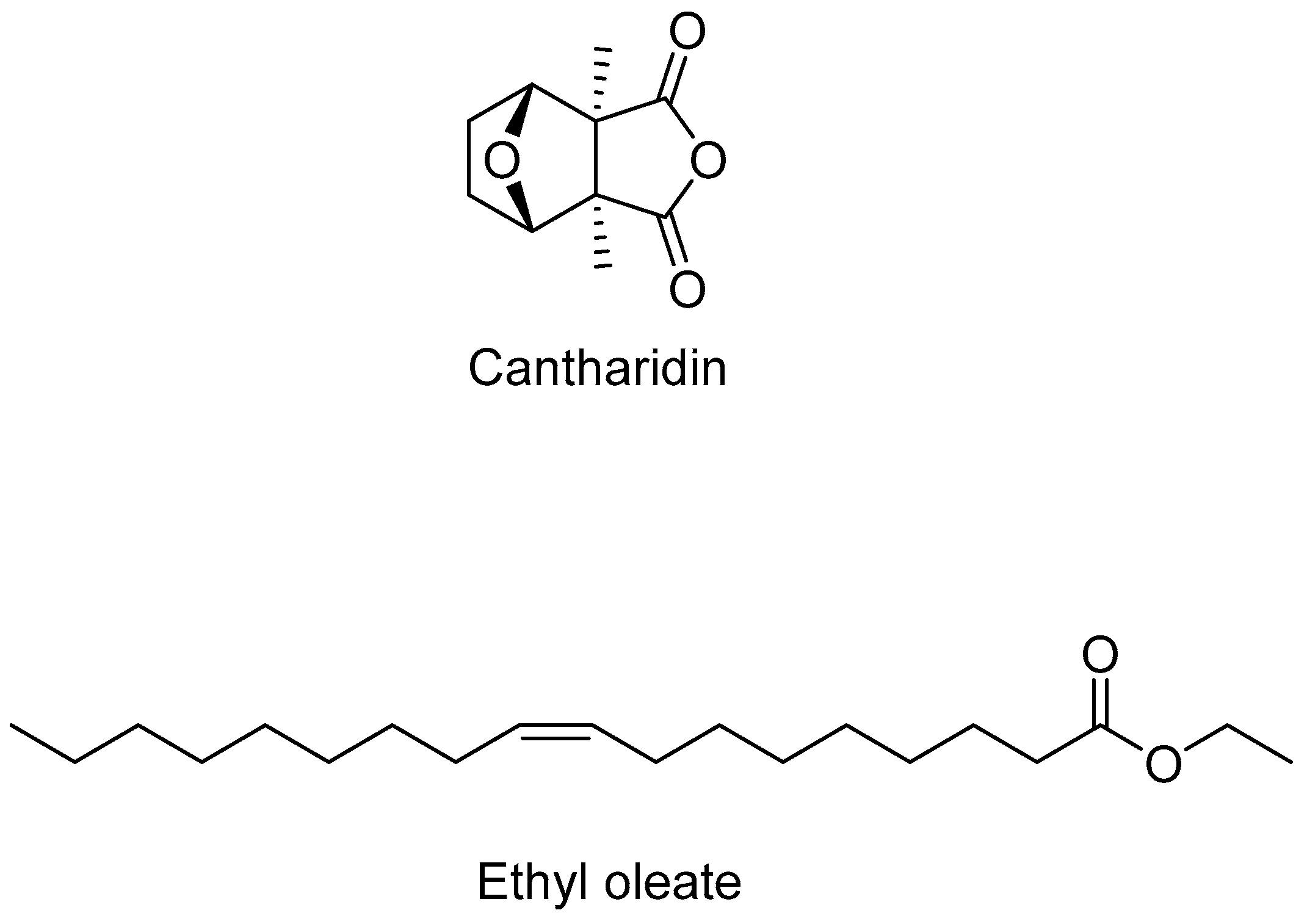
| 5.20 | 2.74 | - | - | - | Cantharidin |
| 7.41 | 1.18 | 1.72 | 0.21 | - | Myristic acid |
| 9.81 | - | 1.57 | 0.41 | - | Muskolactone |
| 10.07 | 9.45 | 10.32 | 7.49 | 7.79 | Hexadecanoic acid |
| 12.37 | 6.29 | 6.34 | 3.43 | 2.83 | Methyl linoleate |
| 12.45 | 21.76 | 23.22 | 15.58 | 10.58 | Muskolactone related |
| 12.71 | 5.03 | 5.10 | 2.95 | 1.91 | Octadecanoic acid |
| 12.81 | - | 1.55 | - | - | Ethyl oleate |
| 15.26 | 2.07 | 2.29 | 4.34 | 6.53 | Oleoamide |
| 16.87–35.60 | 35.58 | 41.79 | 61.47 | 50.76 | Alkanes and derivatives |
| Treatment | Dose (µg/µL) | Growth Inhibition % a | Effective Concentrations b | |
|---|---|---|---|---|
| EC50 µg/µL | EC90 µg/µL | |||
| EO | 0.400 | 66.23 ± 1.57 | 0.3090 (0.3090–0.3100) | 0.5471 (0.5466–0.5476) |
| 0.200 | 34.91 ± 1.11 | |||
| 0.100 | 6.85 ± 0.17 | |||
| CTD | 0.025 | 97.76 ± 7.12 | 0.0085 (0.0084–0.0085) | 0.0287 (0.0286–0.0287) |
| 0.010 | 75.18 ± 5.33 | |||
| 0.005 | 45.63 ± 7.40 | |||
| 0.001 | 0 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Navarro, M.; Bolívar, P.; Andrés, M.F.; Gómez-Muñoz, M.T.; Martínez-Díaz, R.A.; Valcárcel, F.; García-París, M.; Bautista, L.M.; González-Coloma, A. Antiparasitic Effects of Potentially Toxic Beetles (Tenebrionidae and Meloidae) from Steppe Zones. Toxins 2021, 13, 489. https://doi.org/10.3390/toxins13070489
Díaz-Navarro M, Bolívar P, Andrés MF, Gómez-Muñoz MT, Martínez-Díaz RA, Valcárcel F, García-París M, Bautista LM, González-Coloma A. Antiparasitic Effects of Potentially Toxic Beetles (Tenebrionidae and Meloidae) from Steppe Zones. Toxins. 2021; 13(7):489. https://doi.org/10.3390/toxins13070489
Chicago/Turabian StyleDíaz-Navarro, Marta, Paula Bolívar, María Fe Andrés, María Teresa Gómez-Muñoz, Rafael A. Martínez-Díaz, Félix Valcárcel, Mario García-París, Luis M. Bautista, and Azucena González-Coloma. 2021. "Antiparasitic Effects of Potentially Toxic Beetles (Tenebrionidae and Meloidae) from Steppe Zones" Toxins 13, no. 7: 489. https://doi.org/10.3390/toxins13070489
APA StyleDíaz-Navarro, M., Bolívar, P., Andrés, M. F., Gómez-Muñoz, M. T., Martínez-Díaz, R. A., Valcárcel, F., García-París, M., Bautista, L. M., & González-Coloma, A. (2021). Antiparasitic Effects of Potentially Toxic Beetles (Tenebrionidae and Meloidae) from Steppe Zones. Toxins, 13(7), 489. https://doi.org/10.3390/toxins13070489