Mycotoxins, Phytoestrogens and Other Secondary Metabolites in Austrian Pastures: Occurrences, Contamination Levels and Implications of Geo-Climatic Factors

 , , , and
, , , and
Abstract
1. Introduction
2. Results
2.1. Occurrence and Concentrations of the Detected Metabolites
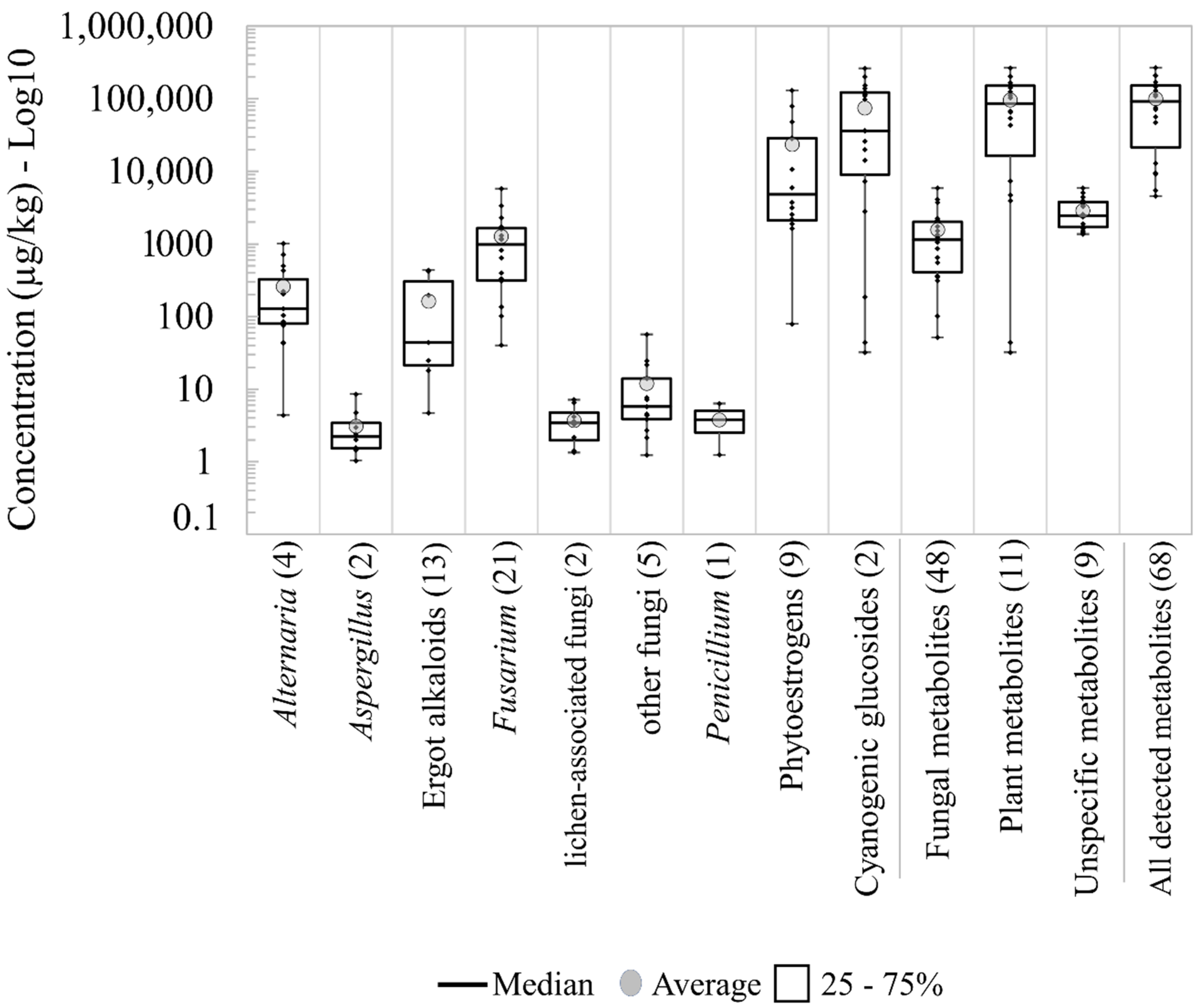
2.1.1. Groups of Metabolites
2.1.2. Regulated Mycotoxins and Related Metabolites
2.1.3. Emerging Mycotoxins
2.1.4. Other Mycotoxins and Metabolites from Fusarium, Alternaria, Aspergillus, Penicillium and Other Fungi
2.1.5. Metabolites from Lichen-Associated and Other Fungi Genera
2.1.6. Plant Compounds (Phytoestrogens and Cyanogenic Glycosides) and Unspecific Metabolites
2.2. Co-Occurrence of Mycotoxins and Other Metabolites
2.3. Effect of Season, Locations and Pasture Diversity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sampling of Pastures
5.2. Mycotoxin Analysis
5.2.1. Chemicals and Reagents
5.2.2. Sample Preparation, Extraction and Estimation of Apparent Recoveries
5.2.3. LC-MS/MS Parameters
5.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McAllister, T.A.; Ribeiro, G.; Stanford, K.; Wang, Y. Forage-Induced Animal Disorders. In Forages, 7th ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 839–860. [Google Scholar] [CrossRef]
- Fletcher, M.T.; Netzel, G. Food Safety and Natural Toxins. Toxins 2020, 12, 236. [Google Scholar] [CrossRef] [PubMed]
- Adams, N.R. Detection of the effects of phytoestrogens on sheep and cattle. J. Anim. Sci. 1995, 73, 1509–1515. [Google Scholar] [CrossRef]
- Di Menna, M.E.; Mortimer, P.H.; Smith, B.L.; Tulloch, M. The incidence of the genus Myrothecium in New Zealand pastures and its relation to animal disease. J. Gen. Microbiol. 1973, 79, 81–87. [Google Scholar] [CrossRef][Green Version]
- Riet-Correa, F.; Rivero, R.; Odriozola, E.; Adrien, M.L.; Medeiros, R.M.T.; Schild, A.L. Mycotoxicoses of ruminants and horses. J. Vet. Diagn. Investig. 2013, 25, 692–708. [Google Scholar] [CrossRef] [PubMed]
- Baholet, D.; Kolackova, I.; Kalhotka, L.; Skladanka, J.; Haninec, P. Effect of Species, Fertilization and Harvest Date on Microbial Composition and Mycotoxin Content in Forage. Agriculture 2019, 9, 102. [Google Scholar] [CrossRef]
- Rodrigues, I. A review on the effects of mycotoxins in dairy ruminants. Anim. Prod. Sci. 2014, 54, 1155–1165. [Google Scholar] [CrossRef]
- Fink-Gremmels, J. The role of mycotoxins in the health and performance of dairy cows. Vet. J. 2008, 176, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Santos Pereira, C.; Cunha, S.C.; Fernandes, J.O. Prevalent mycotoxins in animal feed: Occurrence and analytical methods. Toxins 2019, 11, 290. [Google Scholar] [CrossRef]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef]
- Cinar, A.; Onbasi, E. Mycotoxins: The hidden danger in food. Mycotoxins Food Saf. 2019. [Google Scholar] [CrossRef]
- Battilani, P.; Palumbo, R.; Giorni, P.; Dall’Asta, C.; Dellafiora, L.; Gkrillas, A.; Toscano, P.; Crisci, A.; Brera, C.; De Santis, B. Mycotoxin mixtures in food and feed: Holistic, innovative, flexible risk assessment modelling approach: MYCHIF. EFSA Support. Publ. 2020, 17, 1757E. [Google Scholar] [CrossRef]
- Reed, K.F.M.; Moore, D.D. A preliminary survey of zearalenone and other mycotoxins in Australian silage and pasture. Anim. Prod. Sci. 2009, 49, 696–703. [Google Scholar] [CrossRef]
- Burkin, A.A.; Kononenko, G.P.; Gavrilova, O.P.; Gagkaeva, T.Y. About zearalenone levels in grass fodders and toxine producing activity of Fusarium fungi. Sel’skokhozyaistvennaya Biol. 2015, 50, 255–262. [Google Scholar] [CrossRef][Green Version]
- Orina, A.; Gavrilova, O.P.; Gagkaeva, T.; Burkin, A.; Kononenko, G. The contamination of Fabaceae plants with fungi and mycotoxins. Agric. Food Sci. 2020, 29, 265–275. [Google Scholar] [CrossRef]
- Burkin, A.A.; Kononenko, G.P. Mycotoxin contamination of meadow grasses in European Russia. Sel’skokhozyaistvennaya Biol. 2015, 50, 503–512. [Google Scholar] [CrossRef]
- Gott, P.; Hendel, E.; Lea, K.; Smith, S.; Hofstetter-Schahs, U.; Robbins, K.; Murugesan, G. 423 Ergovaline and Additional Mycotoxins in Tall Fescue. J. Anim. Sci. 2018, 96, 208–209. [Google Scholar] [CrossRef]
- European Commission. Commission Decision of 12 August 2002 implementing Council Directive96/23/EC concerning the performance of analytical methods and the interpretation of results. OJEU 2002, 221, 10–32. [Google Scholar]
- European Commission. Recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding (2006/576/EC). Off. J. Eur. Union 2006, 229, 7–9. [Google Scholar]
- European Commission. Commission Recommendation 2012/154/EU of 15 March 2012 on the monitoring of the presence of ergot alkaloids in feed and food. Off. J. Eur. Union 2012, 77, 20–21. [Google Scholar]
- Rychlik, M.; Humpf, H.-U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Vaclavikova, M.; Malachova, A.; Veprikova, Z.; Dzuman, Z.; Zachariasova, M.; Hajslova, J. ‘Emerging’mycotoxins in cereals processing chains: Changes of enniatins during beer and bread making. Food Chem. 2013, 136, 750–757. [Google Scholar] [CrossRef]
- Vejdovszky, K.; Schmidt, V.; Warth, B.; Marko, D. Combinatory estrogenic effects between the isoflavone genistein and the mycotoxins zearalenone and alternariol in vitro. Mol. Nutr. Food Res. 2017, 61, 1600526. [Google Scholar] [CrossRef]
- Milićević, D.R.; Škrinjar, M.; Baltić, T. Real and perceived risks for mycotoxin contamination in foods and feeds: Challenges for food safety control. Toxins 2010, 2, 572–592. [Google Scholar] [CrossRef]
- Fletcher, M.T.; McKenzie, R.A.; Blaney, B.J.; Reichmann, K.G. Pyrrolizidine alkaloids in Crotalaria taxa from northern Australia: Risk to grazing livestock. J. Agric. Food. Chem. 2009, 57, 311–319. [Google Scholar] [CrossRef]
- Wocławek-Potocka, I.; Korzekwa, A.; Skarzyński, D.J. Can phytoestrogens pose a danger in the reproduction of cows? Med. Weter. 2008, 64, 515–519. [Google Scholar]
- Reed, K.F.M. Fertility of herbivores consuming phytoestrogen-containing Medicago and Trifolium species. Agriculture 2016, 6, 35. [Google Scholar] [CrossRef]
- McGorum, B.; Pirie, R.; Fry, S. Quantification of cyanogenic glycosides in white clover (Trifolium repens L.) from horse pastures in relation to equine grass sickness. Grass Forage Sci. 2012, 67, 274–279. [Google Scholar] [CrossRef]
- Johny, A.; Fæste, C.K.; Bogevik, A.S.; Berge, G.M.; Fernandes, J.M.; Ivanova, L. Development and validation of a liquid chromatography high-resolution mass spectrometry method for the simultaneous determination of mycotoxins and phytoestrogens in plant-based fish feed and exposed fish. Toxins 2019, 11, 222. [Google Scholar] [CrossRef]
- Socas-Rodríguez, B.; Lanková, D.; Urbancová, K.; Krtková, V.; Hernández-Borges, J.; Rodríguez-Delgado, M.Á.; Pulkrabová, J.; Hajšlová, J. Multiclass analytical method for the determination of natural/synthetic steroid hormones, phytoestrogens, and mycoestrogens in milk and yogurt. Anal. Bioanal. Chem. 2017, 409, 4467–4477. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef]
- Pavarini, D.P.; Pavarini, S.P.; Niehues, M.; Lopes, N.P. Exogenous influences on plant secondary metabolite levels. JAFST 2012, 176, 5–16. [Google Scholar] [CrossRef]
- Zhi-lin, Y.; Chuan-chao, D.; Lian-qing, C. Regulation and accumulation of secondary metabolites in plant-fungus symbiotic system. AJB 2007, 6, 1266–1271. [Google Scholar]
- Thakur, M.; Bhattacharya, S.; Khosla, P.K.; Puri, S. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J. Appl. Res. Med. Aroma. 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Szulc, J.; Okrasa, M.; Dybka-Stępień, K.; Sulyok, M.; Nowak, A.; Otlewska, A.; Szponar, B.; Majchrzycka, K. Assessment of Microbiological Indoor Air Quality in Cattle Breeding Farms. AAQR 2019, 20, 1–10. [Google Scholar]
- Hajnal, E.J.; Kos, J.; Malachová, A.; Steiner, D.; Stranska, M.; Krska, R.; Sulyok, M. Mycotoxins in maize harvested in Serbia in the period 2012–2015. Part 2: Non-regulated mycotoxins and other fungal metabolites. Food Chem. 2020, 317, 126409. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Novak, B.; Nagl, V.; Berthiller, F. Emerging mycotoxins: Beyond traditionally determined food contaminants. J. Agric. Food Chem. 2017, 65, 7052–7070. [Google Scholar] [CrossRef]
- Reisinger, N.; Schurer-Waldheim, S.; Mayer, E.; Debevere, S.; Antonissen, G.; Sulyok, M.; Nagl, V. Mycotoxin Occurrence in Maize Silage-A Neglected Risk for Bovine Gut Health? Toxins 2019, 11, 577. [Google Scholar] [CrossRef]
- Gallo, A.; Ghilardelli, F.; Atzori, A.S.; Zara, S.; Novak, B.; Faas, J.; Fancello, F. Co-Occurrence of Regulated and Emerging Mycotoxins in Corn Silage: Relationships with Fermentation Quality and Bacterial Communities. Toxins 2021, 13, 232. [Google Scholar] [CrossRef]
- EFSA. European Food Safety Authority. Scientific opinion on risks for animal and public health related to the presence of nivalenol in food and feed. EFSA J. 2013, 11, 3262. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on the risk for public and animal health related to the presence of sterigmatocystin in food and feed. EFSA J. 2013, 11, 3254. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on the risks for human and animal health related to the presence of modified forms of certain mycotoxins in food and feed. EFSA J. 2014, 12, 3916–4023. [Google Scholar]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; EFSA Panel on Contaminants in the Food Chain (CONTAM); et al. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, e04851. [Google Scholar] [PubMed]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Grasl-Kraupp, B.; EFSA Panel on Contaminants in the Food Chain (CONTAM); et al. Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. Eur. Food Saf. Auth. 2017, 15, e04718. [Google Scholar]
- Nichea, M.J.; Palacios, S.A.; Chiacchiera, S.M.; Sulyok, M.; Krska, R.; Chulze, S.N.; Torres, A.M.; Ramirez, M.L. Presence of multiple mycotoxins and other fungal metabolites in native grasses from a wetland ecosystem in Argentina intended for grazing cattle. Toxins 2015, 7, 3309–3329. [Google Scholar] [CrossRef] [PubMed]
- Nichea, M.J.; Cendoya, E.; Zachetti, V.G.L.; Chiacchiera, S.M.; Sulyok, M.; Krska, R.; Torres, A.M.; Chulze, S.N.; Ramirez, M.L. Mycotoxin profile of Fusarium armeniacum isolated from natural grasses intended for cattle feed. World Mycotoxin J. 2015, 8, 451–457. [Google Scholar] [CrossRef]
- Nesic, K.; Ivanovic, S.; Nesic, V. Fusarial toxins: Secondary metabolites of Fusarium fungi. In Reviews of Environmental Contamination and Toxicology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 228, pp. 101–120. [Google Scholar]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef]
- Kemboi, D.C.; Ochieng, P.E.; Antonissen, G.; Croubels, S.; Scippo, M.-L.; Okoth, S.; Kangethe, E.K.; Faas, J.; Doupovec, B.; Lindahl, J.F. Multi-Mycotoxin Occurrence in Dairy Cattle and Poultry Feeds and Feed Ingredients from Machakos Town, Kenya. Toxins 2020, 12, 762. [Google Scholar] [CrossRef] [PubMed]
- Di Menna, M.E.; Lauren, D.R.; Poole, P.R.; Mortimer, P.H.; Hill, R.A.; Agnew, M.P. Zearalenone in New Zealand pasture herbage and the mycotoxin-producing potential of Fusarium species from pasture. N. Z. J. Agric. Res. 1987, 30, 499–504. [Google Scholar] [CrossRef]
- Gott, P.; Stam, A.; Johns, A.; Miller, B.; Bell, B.; Jenkins, T.; Murugesan, G. 039 Mycotoxin survey of common Bermudagrass in south-central Florida. J. Anim. Sci. 2017, 95, 19–20. [Google Scholar] [CrossRef]
- EFSA, J. Opinion of the scientific panel on contaminants in the food chain on a request from the commission related to zearalenone as undesirable substance in animal feed. EFSA J. 2004, 89, 1–35. [Google Scholar]
- Smith, J.; Di Menna, M.; McGowan, L. Reproductive performance of Coopworth ewes following oral doses of zearalenone before and after mating. Reproduction 1990, 89, 99–106. [Google Scholar] [CrossRef]
- Štýbnarová, M.; Křížová, L.; Pavlok, S.; Mičová, P.; Látal, O.; Pozdíšek, J. Nutritive Value and Mycotoxin Contamination of Herbage in Mountain Locality Exposed to Renewed Cattle Grazing. Acta Univ. Agric. Silvic. Mendel. Brun. 2016, 64, 883–891. [Google Scholar] [CrossRef]
- Ryu, J.-C.; Ohtsubo, K.; Izumiyama, N.; Nakamura, K.; Tanaka, T.; Yamamura, H.; Ueno, Y. The acute and chronic toxicities of nivalenol in mice. Fundam. Appl. Toxicol. 1988, 11, 38–47. [Google Scholar] [CrossRef]
- Del Regno, M.; Adesso, S.; Popolo, A.; Quaroni, A.; Autore, G.; Severino, L.; Marzocco, S. Nivalenol induces oxidative stress and increases deoxynivalenol pro-oxidant effect in intestinal epithelial cells. Toxicol. Appl. Pharmacol. 2015, 285, 118–127. [Google Scholar] [CrossRef]
- Alassane-Kpembi, I.; Puel, O.; Pinton, P.; Cossalter, A.-M.; Chou, T.-C.; Oswald, I.P. Co-exposure to low doses of the food contaminants deoxynivalenol and nivalenol has a synergistic inflammatory effect on intestinal explants. Arch. Toxicol. 2017, 91, 2677–2687. [Google Scholar] [CrossRef]
- Alassane-Kpembi, I.; Puel, O.; Oswald, I.P. Toxicological interactions between the mycotoxins deoxynivalenol, nivalenol and their acetylated derivatives in intestinal epithelial cells. Arch. Toxicol. 2015, 89, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Vejdovszky, K.; Warth, B.; Sulyok, M.; Marko, D. Non-synergistic cytotoxic effects of Fusarium and Alternaria toxin combinations in Caco-2 cells. Toxicol. Lett. 2016, 241, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Křížová, L.; Dadáková, K.; Dvořáčková, M.; Kašparovský, T. Feedborne Mycotoxins Beauvericin and Enniatins and Livestock Animals. Toxins 2021, 13, 32. [Google Scholar] [CrossRef]
- Sy-Cordero, A.A.; Pearce, C.J.; Oberlies, N.H. Revisiting the enniatins: A review of their isolation, biosynthesis, structure determination and biological activities. J. Antibiot. 2012, 65, 541–549. [Google Scholar] [CrossRef]
- Schiff, P.L. Ergot and its alkaloids. Am. J. Pharm. Educ. 2006, 70, 1–10. [Google Scholar] [CrossRef]
- Guerre, P. Ergot alkaloids produced by endophytic fungi of the genus Epichloë. Toxins 2015, 7, 773–790. [Google Scholar] [CrossRef]
- Klotz, J.L. Activities and effects of ergot alkaloids on livestock physiology and production. Toxins 2015, 7, 2801–2821. [Google Scholar] [CrossRef]
- Canty, M.J.; Fogarty, U.; Sheridan, M.K.; Ensley, S.M.; Schrunk, D.E.; More, S.J. Ergot alkaloid intoxication in perennial ryegrass (Lolium perenne): An emerging animal health concern in Ireland? Ir. Vet. J. 2014, 67, 1–7. [Google Scholar] [CrossRef]
- Mostrom, M.S.; Jacobsen, B.J. Ruminant Mycotoxicosis. Vet. Clin. N. Am. Small. Anim. Pract. 2011, 27, 315–344. [Google Scholar] [CrossRef]
- Evans, T.J. Diminished reproductive performance and selected toxicants in forages and grains. Vet. Clin. N. Am. Small. Anim. Pract. 2011, 27, 345–371. [Google Scholar] [CrossRef] [PubMed]
- Marczuk, J.; Zietek, J.; Zwierz, K.; Winiarczyk, S.; Lutnicki, K.; Brodzki, P.; Adaszek, L. Ergovaline poisoning in a herd of dairy cows—A case report. Med. Weteryn. Vet. Med. Sci. Pract. 2019, 75, 635–639. [Google Scholar] [CrossRef]
- Botha, C.; Naude, T.; Moroe, M.; Rottinghaus, G. Gangrenous ergotism in cattle grazing fescue (Festuca elatior L.) in South Africa: Clinical communication. J. S. Afr. Vet. Assoc. 2004, 75, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.H.; Lyons, S.E.; Poole, R.K.; Poore, M.H. Ergot alkaloids induce vasoconstriction of bovine uterine and ovarian blood vessels. J. Anim. Sci. 2018, 96, 4812–4822. [Google Scholar] [CrossRef]
- Nasr, H.; Pearson, O. Inhibition of prolactin secretion by ergot alkaloids. Eur. J. Endocrinol. 1975, 80, 429–443. [Google Scholar] [CrossRef]
- Poole, R.K.; Poole, D.H. Impact of ergot alkaloids on female reproduction in domestic livestock species. Toxins 2019, 11, 364. [Google Scholar] [CrossRef] [PubMed]
- Adams, N. Clover phytoestrogens in sheep in western Australia. Pure Appl. Chem. 1998, 70, 1855–1862. [Google Scholar] [CrossRef][Green Version]
- Smith, J.F.; Jagusch, K.T.; Brunswick, L.E.C.; McGowan, L.T. The effect of lucerne feeding on the ovulation rate in ewes. Proc. N. Z. Soc. Anim. Prod. 1980, 40, 44–49. [Google Scholar]
- Coufal-Majewski, S.; Stanford, K.; McAllister, T.; Blakley, B.; McKinnon, J.; Chaves, A.V.; Wang, Y. Impacts of cereal ergot in food animal production. Front. Vet. Sci. 2016, 3, 15. [Google Scholar] [CrossRef]
- McCormick, S.P.; Bhatnagar, D.; Lee, L.S. Averufanin is an aflatoxin B1 precursor between averantin and averufin in the biosynthetic pathway. Appl. Environ. Microbiol. 1987, 53, 14–16. [Google Scholar] [CrossRef]
- Hsieh, D.; Lin, M.; Yao, R. Conversion of sterigmatocystin to aflatoxin B1 by Aspergillus parasiticus. Biochem. Biophys. Res. Commun. 1973, 52, 992–997. [Google Scholar] [CrossRef]
- Solfrizzo, M. Recent advances on Alternaria mycotoxins. Curr. Opin. Food Sci. 2017, 17, 57–61. [Google Scholar] [CrossRef]
- Vejdovszky, K.; Hahn, K.; Braun, D.; Warth, B.; Marko, D. Synergistic estrogenic effects of Fusarium and Alternaria mycotoxins in vitro. Arch. Toxicol. 2017, 91, 1447–1460. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA J. 2011, 9, 2407. [Google Scholar] [CrossRef]
- Hellwig, V.; Grothe, T.; Mayer-Bartschmid, A.; Endermann, R.; Geschke, F.-U.; Henkel, T.; Stadler, M. Altersetin, a new antibiotic from cultures of endophytic Alternaria spp. Taxonomy, fermentation, isolation, structure elucidation and biological activities. J. Antibiot. 2002, 55, 881–892. [Google Scholar] [CrossRef]
- Daou, R.; Joubrane, K.; Maroun, R.G.; Khabbaz, L.R.; Ismail, A.; El Khoury, A. Mycotoxins: Factors influencing production and control strategies. AIMS Agric. Food. 2021, 6, 416–447. [Google Scholar] [CrossRef]
- Marroquín-Cardona, A.G.; Johnson, N.M.; Phillips, T.D.; Hayes, A.W. Mycotoxins in a changing global environment—A review. Food Chem Toxicol. 2014, 69, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J. Plant age and seasonal timing determine endophyte growth and alkaloid biosynthesis. Fungal Ecol. 2017, 29, 52–58. [Google Scholar] [CrossRef]
- Magan, N.; Medina, A.; Aldred, D. Possible climate-change effects on mycotoxin contamination of food crops pre-and postharvest. Plant Pathol. 2011, 60, 150–163. [Google Scholar] [CrossRef]
- Medina, A.; Akbar, A.; Baazeem, A.; Rodriguez, A.; Magan, N. Climate change, food security and mycotoxins: Do we know enough? Fungal Biol. Rev. 2017, 31, 143–154. [Google Scholar] [CrossRef]
- Medina, Á.; Rodríguez, A.; Magan, N. Climate change and mycotoxigenic fungi: Impacts on mycotoxin production. Curr. Opin. Food Sci. 2015, 5, 99–104. [Google Scholar] [CrossRef]
- Mostrom, M.; Evans, T.J. Phytoestrogens. In Reproductive and Developmental Toxicology; Gupta, R.C., Ed.; Academic Press—Medical: Amsterdam, The Netherlands, 2011; pp. 707–722. [Google Scholar]
- Chatenet, C. Les phytoestrogènes. Actual. Pharm. 2008, 47, 10–23. [Google Scholar]
- Medlock, K.L.; Branham, W.S.; Sheehan, D.M. Effects of coumestrol and equol on the developing reproductive tract of the rat. Proc. Soc. Exp. Biol. Med. 1995, 208, 67–71. [Google Scholar] [CrossRef]
- Romero-R, C.M.; Castellanos, M.d.R.T.; Mendoza, R.M.; Reyes, R.A.; García, A.R. Oestrogenic syndrome in dairy cows by alfalfa comsuption with large amount of coumestrol. Vet. Mex 1997, 28, 25–30. [Google Scholar]
- Coop, I.E. Depression of lambing performance from mating on lucerne. Proc. N. Z. Soc. Anim. Prod. 1977, 37, 149–151. [Google Scholar]
- Shemesh, M.; Shore, L.S. Effects of Environmental Estrogens on Reproductive Parameters in Domestic Animals. Isr. J. Vet. Med. 2012, 67, 6–10. [Google Scholar]
- Sulyok, M.; Stadler, D.; Steiner, D.; Krska, R. Validation of an LC-MS/MS-based dilute-and-shoot approach for the quantification of >500 mycotoxins and other secondary metabolites in food crops: Challenges and solutions. Anal. Bioanal. Chem. 2020, 2607–2620. [Google Scholar] [CrossRef] [PubMed]
- Steiner, D.; Sulyok, M.; Malachova, A.; Mueller, A.; Krska, R. Realizing the simultaneous liquid chromatography-tandem mass spectrometry based quantification of >1200 biotoxins, pesticides and veterinary drugs in complex feed. J. Chromatogr. A 2020, 1629, 461502. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Metabolite | Positive Samples (%) 1 | Concentration (μg/kg DM) 2 | ||
|---|---|---|---|---|---|
| Average ± SD | Median | Range | |||
| Alternaria | Alternariol 3 | 61 | 6.41 ± 7.43 | 2.81 | 1.00–23.7 |
| Alternariolmethylether 3 | 56 | 7.30 ± 8.30 | 4.45 | 1.01–29.4 | |
| Altersetin | 83 | 220 ± 246 | 127 | 4.36–861 | |
| Infectopyrone | 33 | 76.5 ± 78.7 | 36.3 | 16.3–212 | |
| Total 4 | 83 | 260 ± 286 | 128 | 4.36–1010 | |
| Aspergillus | Averufin | 6 | - | - | 1.15 |
| Sterigmatocystin 3 | 44 | 2.94 ± 2.13 | 2.21 | 1.03–7.34 | |
| Total 4 | 44 | 3.08 ± 2.48 | 2.21 | 1.03–8.49 | |
| Ergot alkaloids 5 | Chanoclavine | 17 | 152 ± 245 | 17.93 | 2.35–435 |
| Ergocornine | 22 | 20.1 ± 26.1 | 7.83 | 5.57–59.2 | |
| Ergocorninine | 22 | 8.72 ± 8.83 | 4.86 | 3.27–21.9 | |
| Ergocristine | 17 | 38.0 ± 31.9 | 37.5 | 6.33–70.1 | |
| Ergocristinine | 17 | 8.21 ± 5.71 | 8.64 | 2.30–13.7 | |
| Ergocryptine | 28 | 24.8 ± 28.4 | 9.27 | 3.6–71.5 | |
| Ergocryptinine | 17 | 6.12 ± 6.30 | 3.01 | 1.97–13.4 | |
| Ergometrine | 22 | 8.76 ± 6.19 | 7.80 | 2.38–17.1 | |
| Ergometrinine | 11 | 1.92 ± 0.26 | 1.92 | 1.73–2.1 | |
| Ergosine | 22 | 15.9 ± 13.5 | 15.1 | 1.1–32.1 | |
| Ergosinine | 17 | 3.99 ± 2.39 | 3.24 | 2.06–6.66 | |
| Ergotamine | 11 | 75.7 ± 93.3 | 75.7 | 9.7–142 | |
| Ergotaminine | 11 | 11.6 ± 13.2 | 11.6 | 2.24–20.9 | |
| Total 4 | 39 | 163 ± 191 | 43.9 | 4.70–435 | |
| Fusarium | 15-Hydroxyculmorin 3 | 44 | 152 ± 243 | 39.2 | 13.0–721 |
| Antibiotic Y | 67 | 254 ± 374 | 66.5 | 45.5–1290 | |
| Apicidin 3 | 39 | 31.3 ± 31.5 | 25.9 | 5.84–97.9 | |
| Aurofusarin 3 | 83 | 196 ± 213 | 133 | 7.89–835 | |
| Beauvericin 3 | 44 | 3.99 ± 3.03 | 2.6 | 1.02–9.34 | |
| Chrysogine | 61 | 13.6 ± 15.5 | 7.42 | 4.07–58.2 | |
| Culmorin 3 | 89 | 129 ± 216 | 51.1 | 9.53–882 | |
| Deoxynivalenol 5 | 11 | 306 ± 281 | 306 | 107–505 | |
| DON-3-glucoside 6 | 6 | - | - | 102 | |
| Enniatin A 3 | 6 | - | - | 2.01 | |
| Enniatin A1 3 | 44 | 5.54 ± 6.03 | 2.92 | 1.22–19.1 | |
| Enniatin B 3 | 94 | 38.3 ± 63.9 | 11.8 | 1.30–241 | |
| Enniatin B1 3 | 89 | 15.3 ± 24.8 | 5.49 | 1.19–93.3 | |
| Enniatin B2 3 | 28 | 3.41 ± 2.74 | 2.27 | 1.19–7.90 | |
| Epiequisetin 3 | 56 | 9.27 ± 7.96 | 8.09 | 1.18–27.2 | |
| Equisetin 3 | 67 | 57.9 ± 60.4 | 37.6 | 2.72–179 | |
| HT-2 Glucoside 6 | 6 | - | - | 14.0 | |
| Moniliformin 3 | 100 | 5.70 ± 3.52 | 5.79 | 1.45–13.1 | |
| Nivalenol | 83 | 170 ± 182 | 78.6 | 38.1–574 | |
| Siccanol 3 | 61 | 716 ± 392 | 758 | 119.3–1480 | |
| Zearalenone 5 | 50 | 29.6 ± 44.3 | 9.93 | 2.61–138 | |
| Sum of enniatins | 94 | 57.4 ± 95.5 | 18.5 | 1.3–364 | |
| Sum of type B Trichothecenes | 83 | 218 ± 289 | 78.6 | 38.1–1070 | |
| Total 4 | 100 | 1280 ± 1430 | 983 | 40.2–5770 | |
| Penicillium | Pestalotin | 11 | 3.79 ± 3.60 | 3.79 | 1.24–6.33 |
| Total 4 | 11 | 3.79 ± 3.60 | 3.79 | 1.24–6.33 | |
| lichen-associated fungi | Lecanoric acid | 39 | 2.31 ± 0.86 | 2.17 | 1.34–3.60 |
| Usnic acid | 17 | 4.49 ± 0.53 | 4.19 | 4.18–5.10 | |
| Total 4 | 44 | 3.71 ± 2.18 | 3.44 | 1.34–7.13 | |
| other fungi | Ilicicolin A | 22 | 1.92 ± 0.98 | 1.83 | 1.00–3.02 |
| Ilicicolin B | 44 | 4.00 ± 3.33 | 2.85 | 1.23–11.7 | |
| Ilicicolin E | 11 | 1.44 ± 0.11 | 1.44 | 1.36–1.51 | |
| Rubellin D | 17 | 5.00 ± 5.00 | 2.7 | 1.56–10.7 | |
| Monocerin | 50 | 11.0 ± 11.8 | 2.97 | 1.32–33.4 | |
| Total 4 | 72 | 12.0 ± 15.4 | 5.73 | 1.23–56.9 | |
| Sum of fungal metabolites | 100 | 1570 ± 1580 | 1145 | 51.7–5880 | |
| Phytoestrogens | Biochanin | 89 | 7060 ± 7560 | 3240 | 62.1–20,650 |
| Coumestrol | 67 | 41.6 ± 34.4 | 32.9 | 7.88–130 | |
| Daidzein | 83 | 936 ± 1840 | 139 | 5.16–6110 | |
| Daidzin | 33 | 167 ± 200 | 88.7 | 15.8–543 | |
| Genistein | 83 | 2760 ± 4780 | 704 | 28.4–17,550 | |
| Genistin | 50 | 311 ± 513 | 139 | 14.6–1630 | |
| Glycitein | 83 | 7470 ± 10,700 | 1500 | 315–35,850 | |
| Ononin | 83 | 2230 ± 4210 | 186 | 47.1–15,130 | |
| Sissotrine | 78 | 4210 ± 9050 | 331 | 8.19–33,070 | |
| Total 4 | 89 | 23,570 ± 35,920 | 4850 | 78.8–130,530 | |
| Cyanogenic glucosides | Linamarin | 83 | 50,620 ± 44,880 | 49,790 | 2030–147,500 |
| Lotaustralin | 100 | 32,6200 ± 34,640 | 16,850 | 32.1–115,900 | |
| Total 4 | 100 | 74,800 ± 79,000 | 36,400 | 32.1–263,400 | |
| Sum of plant metabolites | 100 | 95,760 ± 81,560 | 85,700 | 32.1–265,3200 | |
| Unspecific | 3-Nitropropionic acid | 11 | 4.87 ± 1.91 | 4.87 | 3.52–6.22 |
| Brevianamid F | 100 | 18.9 ± 13.7 | 14.1 | 6.50–62.4 | |
| Citreorosein | 50 | 18.1 ± 12.4 | 16.6 | 4.52–44.9 | |
| cyclo(L-Pro-L-Tyr) | 100 | 498 ± 347 | 361 | 172–1383 | |
| cyclo(L-Pro-L-Val) | 100 | 2190 ± 1000 | 1970 | 1080–4290 | |
| Endocrocin | 11 | 17.4 ± 6.77 | 17.4 | 12.6–22.1 | |
| Iso-Rhodoptilometrin | 22 | 2.25 ± 0.95 | 1.96 | 1.49–3.60 | |
| Rugulusovine | 100 | 13.7 ± 8.60 | 11.7 | 3.75–39.0 | |
| Tryptophol | 100 | 127 ± 118 | 74.0 | 53.1–485 | |
| Sum of unspecific metabolites | 100 | 2860 ± 1380 | 2460 | 1370–5910 | |
| Sum of all detected metabolites | 100 | 100,200 ± 80,900 | 92,100 | 4560–266,700 | |
| Variable | Early | Late | SEM 1 | p-Value |
|---|---|---|---|---|
| Number metabolites/sample | ||||
| All metabolites | 24.4 | 39.6 | 3.51 | 0.008 |
| Fungal metabolites | 11.8 | 24.0 | 3.03 | 0.012 |
| Concentration (µg/kg) | ||||
| from Alternaria | 76 | 329 | 85.0 | 0.052 |
| from Aspergillus | 1.61 | 1.18 | 0.77 | 0.693 |
| Ergot Alkaloids | 5.32 | 110 | 44.6 | 0.120 |
| from Fusarium | 526 | 1890 | 431.8 | 0.041 |
| from Lichen | 1.76 | 1.56 | 0.81 | 0.865 |
| from other fungi species | 1.24 | 14.6 | 4.23 | 0.041 |
| from Penicillium | 0.00 | 0.76 | 0.50 | 0.303 |
| Fungal Metabolites | 611 | 2332 | 452 | 0.017 |
| Phytoestrogens | 7867 | 31,420 | 11,195 | 0.158 |
| Cyanogenic glycosides | 71,666 | 77,318 | 27,251 | 0.886 |
| Plant metabolites | 79,532 | 108,738 | 27,678 | 0.468 |
| Unspecific metabolites | 3144 | 4291 | 646 | 0.083 |
| Total Metabolites | 82,556 | 114,294 | 27,363 | 0.426 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penagos-Tabares, F.; Khiaosa-ard, R.; Nagl, V.; Faas, J.; Jenkins, T.; Sulyok, M.; Zebeli, Q. Mycotoxins, Phytoestrogens and Other Secondary Metabolites in Austrian Pastures: Occurrences, Contamination Levels and Implications of Geo-Climatic Factors. Toxins 2021, 13, 460. https://doi.org/10.3390/toxins13070460
Penagos-Tabares F, Khiaosa-ard R, Nagl V, Faas J, Jenkins T, Sulyok M, Zebeli Q. Mycotoxins, Phytoestrogens and Other Secondary Metabolites in Austrian Pastures: Occurrences, Contamination Levels and Implications of Geo-Climatic Factors. Toxins. 2021; 13(7):460. https://doi.org/10.3390/toxins13070460
Chicago/Turabian StylePenagos-Tabares, Felipe, Ratchaneewan Khiaosa-ard, Veronika Nagl, Johannes Faas, Timothy Jenkins, Michael Sulyok, and Qendrim Zebeli. 2021. "Mycotoxins, Phytoestrogens and Other Secondary Metabolites in Austrian Pastures: Occurrences, Contamination Levels and Implications of Geo-Climatic Factors" Toxins 13, no. 7: 460. https://doi.org/10.3390/toxins13070460
APA StylePenagos-Tabares, F., Khiaosa-ard, R., Nagl, V., Faas, J., Jenkins, T., Sulyok, M., & Zebeli, Q. (2021). Mycotoxins, Phytoestrogens and Other Secondary Metabolites in Austrian Pastures: Occurrences, Contamination Levels and Implications of Geo-Climatic Factors. Toxins, 13(7), 460. https://doi.org/10.3390/toxins13070460