Domoic Acid and Pseudo-nitzschia spp. Connected to Coastal Upwelling along Coastal Inhambane Province, Mozambique: A New Area of Concern

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
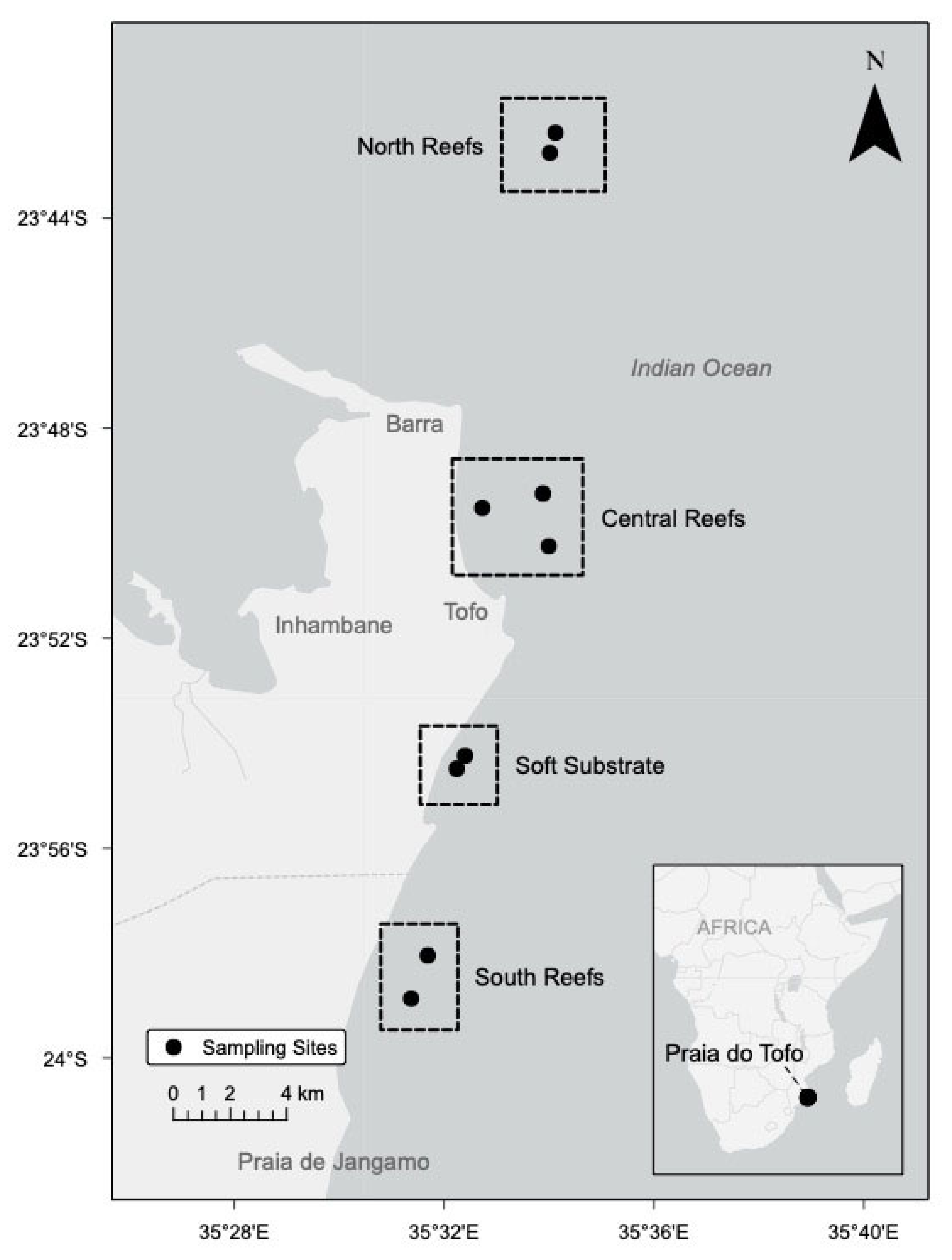
2.1. Sites
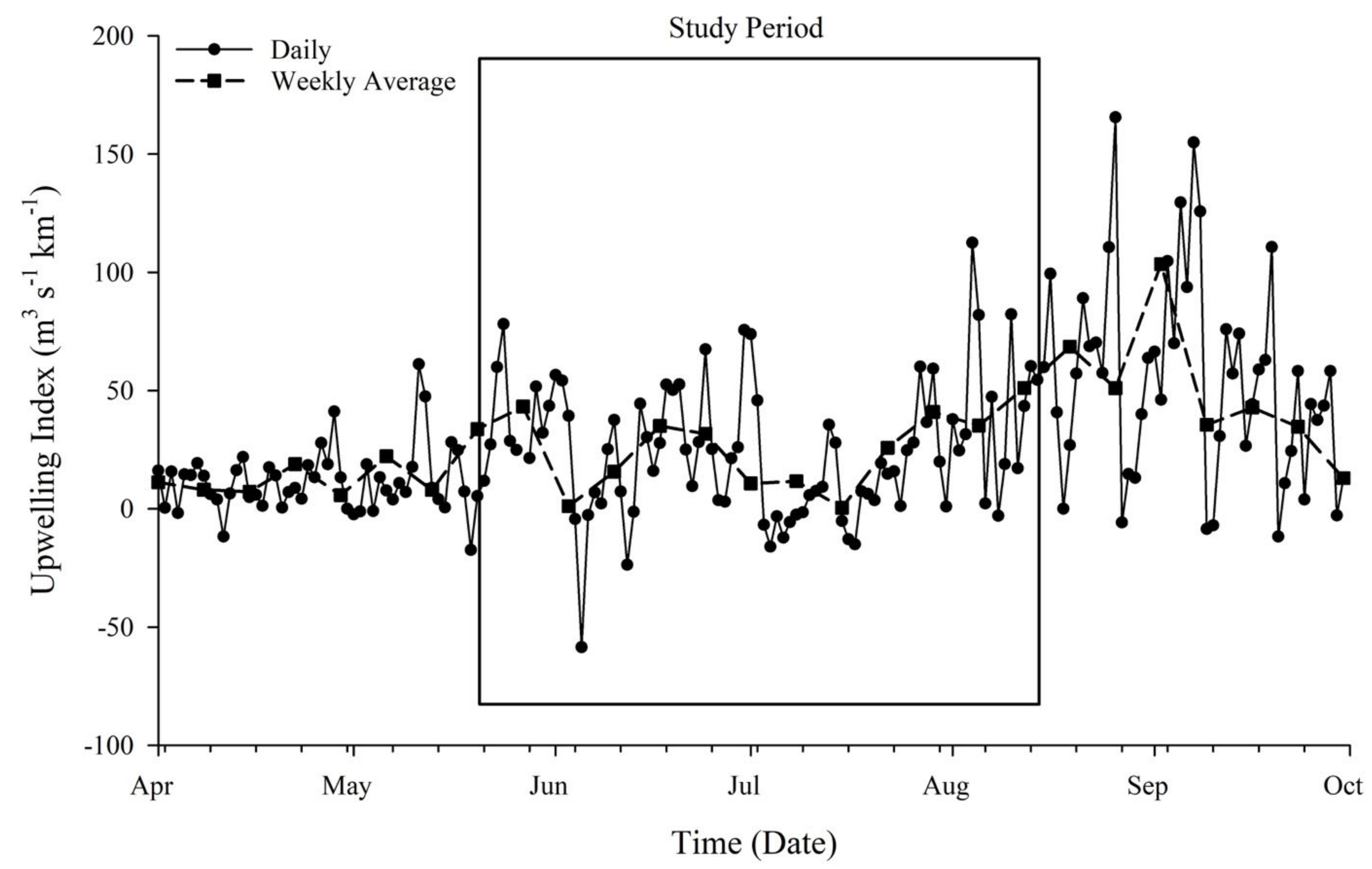
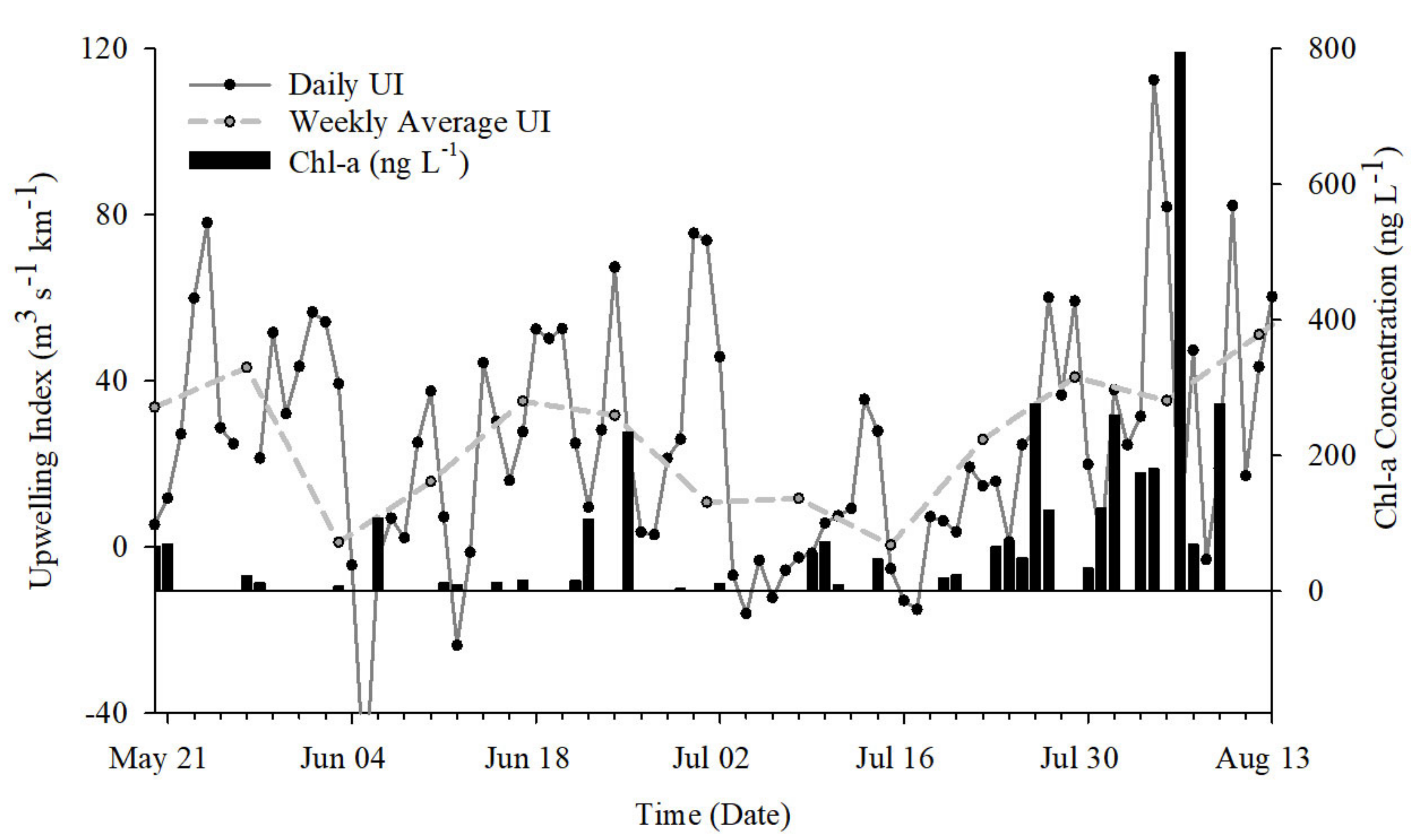
2.2. Upwelling Index
2.3. Environmental Conditions
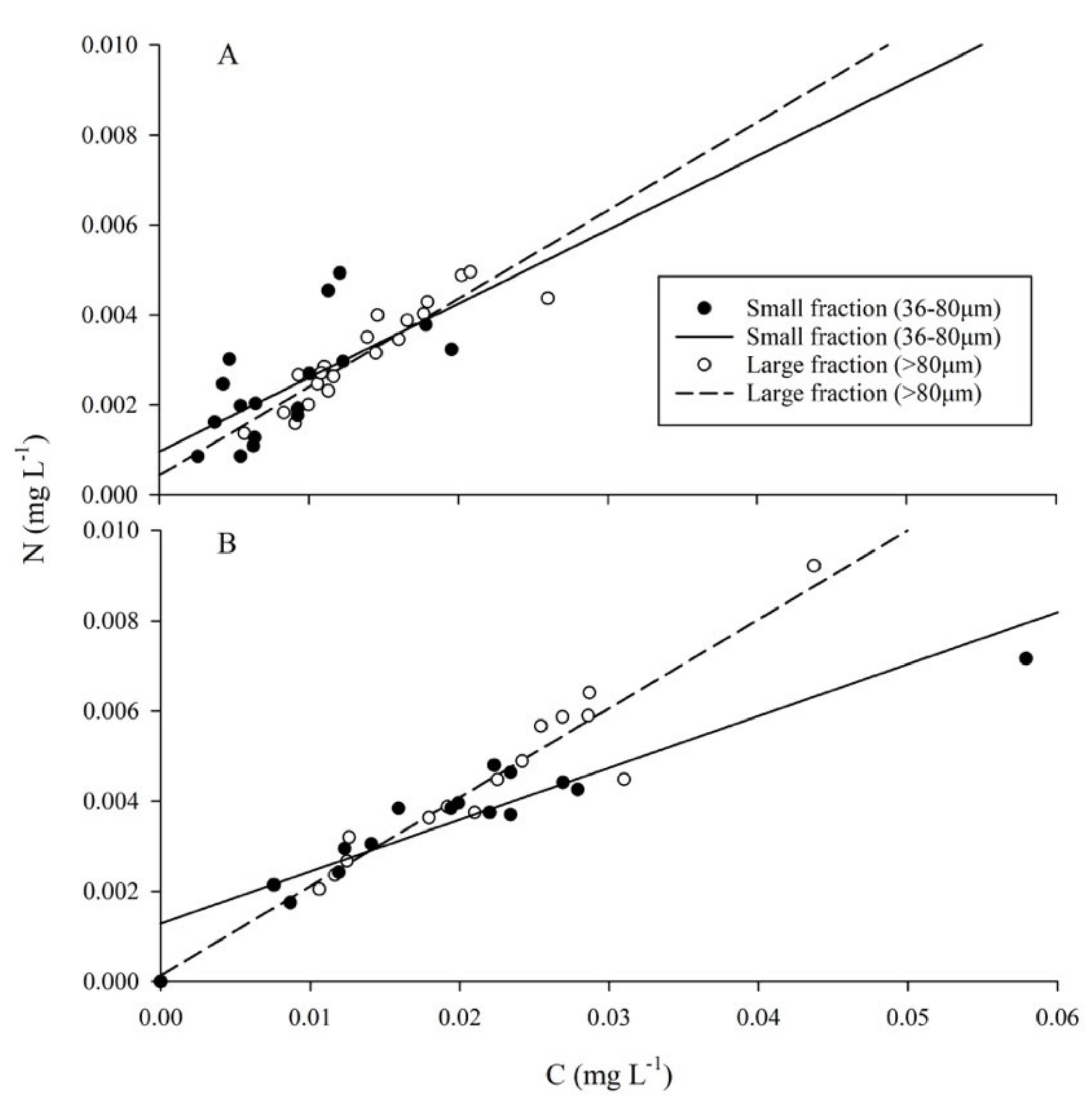
2.4. Particulate Organic Matter
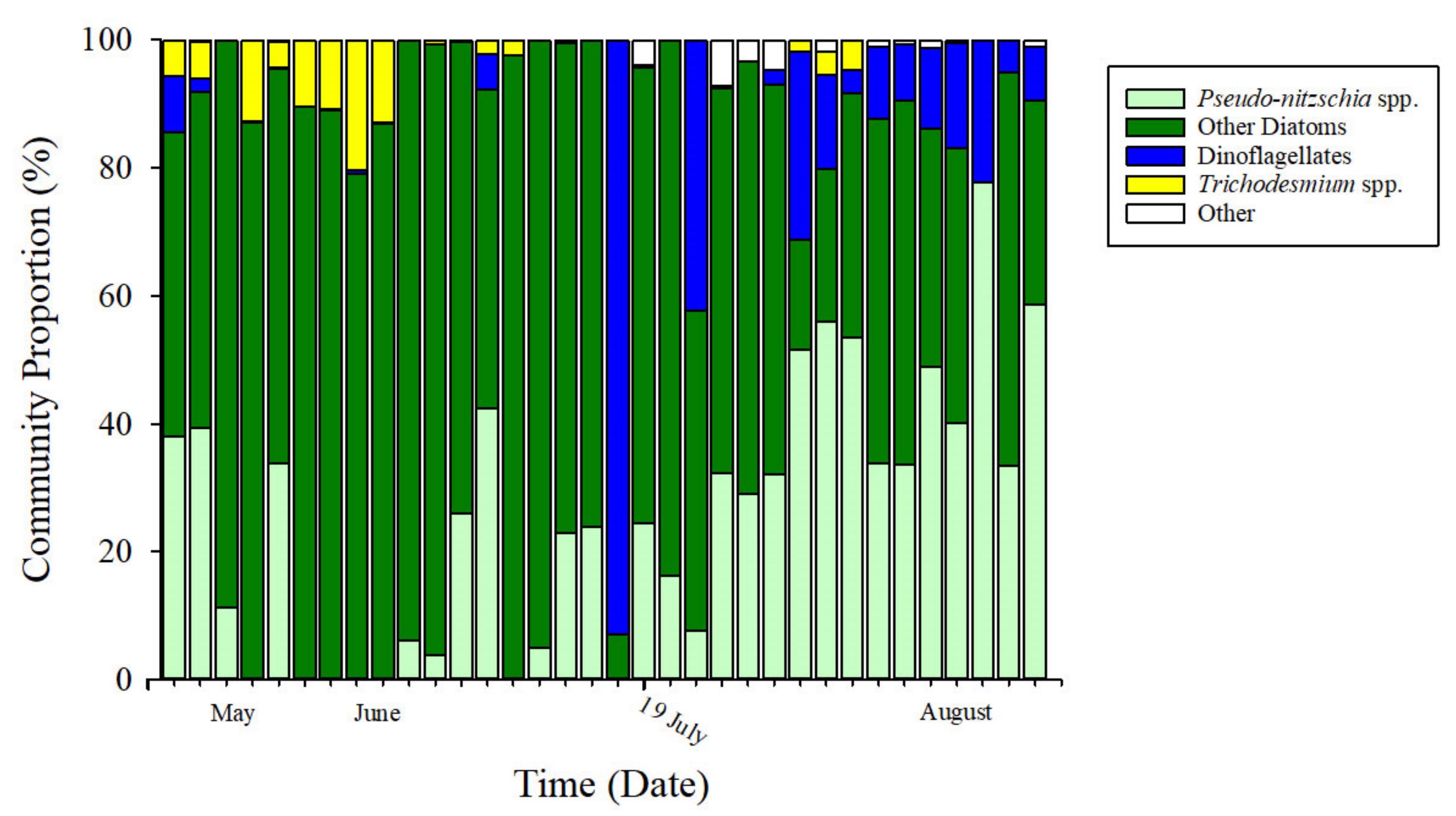
2.5. Phytoplankton Composition
2.6. Particulate DA
3. Discussion
4. Methods
4.1. Study Site
4.2. Satellite Imagery and Analysis
4.3. Upwelling Index Calculations
4.4. Field Site Monitoring and Sample Collection
4.5. Dissolved Nutrient Analysis
4.6. Pigment Analysis
4.7. Particulate Domoic Acid Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bakun, A.; Nelson, C.S. The seasonal cycle of wind-stress curl in subtropical Eastern Boundary Current regions. J. Phys. Oceanogr. 1991, 21, 1815–1834. [Google Scholar] [CrossRef]
- Carr, M.-E.; Kearns, E.J. Production regimes in four Eastern Boundary Current systems. Deep Sea Res. Pt. II Top. Stud. Oceanogr. 2003, 50, 3199–3221. [Google Scholar] [CrossRef]
- Doblin, M.A.; Petrou, K.; Sinutok, S.; Seymour, J.R.; Messer, L.F.; Brown, M.V.; Norman, L.; Everett, J.D.; McInnes, A.S.; Ralph, P.J.; et al. Nutrient uplift in a cyclonic eddy increases diversity, primary productivity and iron demand of microbial communities relative to a western boundary current. Peer J. 2016, 4, e1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everett, J.D.; Doblin, M.A. Characterising primary productivity measurements across a dynamic western boundary current region. Deep Sea Res. Pt. I Oceanogr. Res. Pap. 2015, 100, 105–116. [Google Scholar] [CrossRef]
- Kimura, S.; Kasai, A.; Nakata, H.; Sugimoto, T.; Simpson, J.H.; Cheok, J.V.S. Biological productivity of meso-scale eddies caused by frontal disturbances in the Kuroshio. ICES J. Mar. Sci. 1997, 54, 179–192. [Google Scholar] [CrossRef] [Green Version]
- Behagle, N.; du Buisson, L.; Josse, E.; Lebourges-Dhaussy, A.; Roudaut, G.; Menard, F. Mesoscale features and micronekton in the Mozambique Channel: An acoustic approach. Deep Sea Res. Pt. Ii 2014, 100, 164–173. [Google Scholar] [CrossRef]
- Wells, R.J.D.; Rooker, J.R.; Quigg, A.; Wissel, B. Influence of mesoscale oceanographic features on pelagic food webs in the Gulf of Mexico. Mar. Biol. 2017, 164, 92. [Google Scholar] [CrossRef]
- Davidson, K.; Gowen, R.J.; Harrison, P.J.; Fleming, L.E.; Hoagland, P.; Moschonas, G. Anthropogenic nutrients and harmful algae in coastal waters. J. Environ. Manag. 2014, 146, 206–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graneli, E.; Turner, J.T. Ecology of Harmful Algae; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef]
- Smayda, T.J. Novel and nuisance phytoplankton blooms in the sea: Evidence for a global epidemic. In Toxic Marine Phytoplankton; Graneli, E., Sundstorm, B., Edler, I., Anderson, D.M., Eds.; Elsevier: New York, NY, USA, 1990; pp. 29–40. [Google Scholar]
- Van Dolah, F.M. Marine algal toxins: Origins, health effects, and their increased occurrence. Environ. Health Persp. 2000, 108, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, D.M.; Hoagland, P.; Kaoru, Y.; White, A.W. Estimated Annual Economic Impacts from Harmful Algal Blooms (HABs) in the United States; Woods Hole Oceanographic Institution: Falmouth, MA, USA, 2000. [Google Scholar]
- Grattan, L.M.; Holobaugh, S.; Morris, J.G. Harmful algal blooms and public health. Harmful Algae 2016, 57, 2–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, C.; Papathanasopoulou, E.; Russel, D.; Artioli, Y. Harmful algal blooms: The impacts on cultural ecosystem services and human well-being in a case study setting, Cornwall, UK. Mar. Policy 2018, 97, 232–238. [Google Scholar] [CrossRef]
- Bates, S.S.; Bird, C.J.; Defreitas, A.S.W.; Foxall, R.; Gilgan, M.; Hanic, L.A.; Johnson, G.R.; McCulloch, A.W.; Odense, P.; Pocklington, R.; et al. Pennate diatom Nitzschia-pungens as the primary source of domoic acid, a toxin in shellfish from eastern Prince Edward Island, Canada. Can. J. Fish. Aquat. Sci. 1989, 46, 1203–1215. [Google Scholar] [CrossRef]
- Bargu, S.; Smith, E.; Ozhan, K. Toxic diatom Pseudo-nitzschia and its primary consumers (vectors). In The Diatom World; Cellular Origin, Life in Extreme Habitats and Astrobiology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 491–512. [Google Scholar]
- D’Agostino, V.C.; Degrati, M.; Sastre, V.; Santinelli, N.; Krock, B.; Krohn, T.; Dans, S.L.; Hoffmeyer, M.S. Domoic acid in a marine pelagic food web: Exposure of southern right whales Eubalaena australis to domoic acid on the Peninsula Valdes calving ground, Argentina. Harmful Algae 2017, 68, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, K.A.; Bargu, S.; Kieckhefer, T.; Silver, M.W. From sanddabs to blue whales: The pervasiveness of domoic acid. Toxicon 2002, 40, 971–977. [Google Scholar] [CrossRef]
- Work, T.M.; Barr, B.; Beale, A.M.; Fritz, L.; Quilliam, M.A.; Wright, J.L.C. Epidemiology of domoic acid poisoning in brown pelicans (Pelecanus occidentalis) and Brandt’s cormorants (Phalacrocorax penicillatus) in California. J. Zoo. Wildl. Med. 1993, 24, 54–62. [Google Scholar]
- Zabaglo, K.; Chrapusta, E.; Bober, B.; Kaminski, A.; Adamski, M.; Bialczyk, J. Environmental roles and biological activity of domoic acid: A review. Algal Res. 2016, 13, 94–101. [Google Scholar] [CrossRef]
- Jeffery, B.; Barlow, T.; Moizer, K.; Paul, S.; Boyle, C. Amnesic shellfish poison. Food Chem. Toxicol. 2004, 42, 545–557. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Robertson, A. Domoic acid and human exposure risks: A review. Toxicon 2010, 56, 218–230. [Google Scholar] [CrossRef]
- Bates, S.S.; Hubbard, K.A.; Lundholm, N.; Montresor, M.; Leaw, C.P. Pseudo-nitzschia, Nitzschia, and domoic acid: New research since 2011. Harmful Algae 2018, 79, 3–43. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.F.; Awan, S.A.; Ling, S.; Wang, R.; Wang, S. Domoic acid: Attributes, exposure risks, innovative detection techniques and therapeutics. Algal Res. 2017, 24, 97–110. [Google Scholar] [CrossRef]
- Trainer, V.L.; Bates, S.S.; Lundholm, N.; Thessen, A.E.; Cochlan, W.P.; Adams, N.G.; Trick, C.G. Pseudo-nitzschia physiological ecology, phylogeny, toxicity, monitoring and impacts on ecosystem health. Harmful Algae 2012, 14, 271–300. [Google Scholar] [CrossRef] [Green Version]
- Pitcher, G.; Pillar, S. Harmful algal blooms in eastern boundary upwelling systems. Prog. Oceanogr. 2010, 85, 1–4. [Google Scholar] [CrossRef]
- Trainer, V.L.; Pitcher, G.C.; Reguera, B.; Smayda, T.J. The distribution and impacts of harmful algal bloom species in eastern boundary upwelling systems. Prog. Oceanogr. 2010, 85, 33–52. [Google Scholar] [CrossRef]
- Fawcett, A.; Pitcher, G.C.; Bernard, S.; Cembella, A.D.; Kudela, R.M. Contrasting wind patterns and toxigenic phytoplankton in the southern Benguela upwelling system. Mar. Ecol. Prog. Ser. 2007, 348, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Trainer, V.L.; Adams, N.G.; Bill, B.D.; Stehr, C.M.; Wekell, J.C.; Moeller, P.; Busman, M.; Woodruff, D. Domoic acid production near California coastal upwelling zones, June 1998. Limnol. Oceanogr. 2000, 45, 1818–1833. [Google Scholar] [CrossRef] [Green Version]
- Trainer, V.L.; Hickey, B.M.; Homer, R.A. Biological and physical dynamics of domoic acid production off the Washington coast. Limnol. Oceanogr. 2002, 47, 1438–1446. [Google Scholar] [CrossRef]
- Anderson, C.R.; Sapiano, M.R.P.; Prasad, M.B.K.; Long, W.; Tango, P.J.; Brown, C.W.; Murtugudde, R. Predicting potentially toxigenic Pseudo-nitzschia blooms in the Chesapeake Bay. J. Mar. Syst. 2010, 83, 127–140. [Google Scholar] [CrossRef]
- Jiang, R.; Wang, Y.-S. Modeling the ecosystem response to summer coastal upwelling in the northern South China Sea. Oceanologia 2018, 60, 32–51. [Google Scholar] [CrossRef]
- Van Meerssche, E.; Pinckney, J.L. The influence of salinity in the domoic acid effect on estuarine phytoplankton communities. Harmful Algae 2017, 69, 65–74. [Google Scholar] [CrossRef]
- McKibben, S.M.; Peterson, W.; Wood, A.M.; Trainer, V.L.; Hunter, M.; White, A.E. Climatic regulation of the neurotoxin domoic acid. Proc. Natl. Acad. Sci. USA 2017, 114, 239–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhodes, L.; Jiang, W.; Knight, B.; Adamson, J.; Smith, K.; Langi, V.; Edgar, M. The genus Pseudo-nitzschia (Bacillariophyceae) in New Zealand: Analysis of the last decade’s monitoring data. N. Z. J. Mar. Fresh. 2013, 47, 490–503. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, L.; Scholin, C.; Garthwaite, I. Pseudo-nitzschia in New Zealand and the role of DNA probes and immunoassays in refining marine biotoxin monitoring programmes. Nat. Toxins 1998, 6, 105–111. [Google Scholar] [CrossRef]
- Almandoz, G.O.; Fabro, E.; Ferrario, M.; Tillmann, U.; Cembella, A.; Krock, B. Species occurrence of the potentially toxigenic diatom genus Pseudo-nitzschia and the associated neurotoxin domoic acid in the Argentine Sea. Harmful Algae 2017, 63, 45–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negri, R.M.; Montoya, N.G.; Carreto, J.I.; Akselman, R.; Inza, D. Pseudo-nitzschia australis, Mytilus edulis, Engraulis anchoita, and domoic acid in the Argentine Sea. In Harmful Algae; Reguera, B., Blanco, J., Fernandes, M.L., Wyatt, T., Eds.; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 2004. [Google Scholar]
- Tamele, I.J.; Silva, M.; Vasconcelos, V. The incidence of marine toxins and the associated seafood poisoning episodes in the African countries of the Indian Ocean and the Red Sea. Toxins 2019, 11, 58. [Google Scholar] [CrossRef] [Green Version]
- Boisier, P.; Ranaivoson, G.; Rasolofonirina, N.; Andriamahefazafy, B.; Roux, J.; Chanteau, S.; Satake, M.; Yasumoto, T. Fatal mass poisoning in Madagascar following ingestion of a shark (Carcharhinus leucas): Clinical and epidemiological aspects and isolation of toxins. Toxicon 1995, 33, 1359–1364. [Google Scholar] [CrossRef]
- Bouaïcha, N.; Chézeau, A.; Turquet, J.; Quod, J.P.; Puiseux-Dao, S. Morphological and toxicological variability of Prorocentrum lima clones isolated from four locations in the south-west Indian Ocean. Toxicon 2001, 39, 1195–1202. [Google Scholar] [CrossRef]
- Pitcher, G.C.; Calder, D. Harmful algal blooms of the southern Benguela Current: A review and appraisal of monitoring from 1989 to 1997. S. Afr. J. Mar. Sci. 2000, 22, 255–271. [Google Scholar] [CrossRef]
- Pitcher, G.C.; Cembella, A.D.; Krock, B.; Macey, B.M.; Mansfield, L.; Probyn, T.A. Identification of the marine diatom Pseudo-nitzschia multiseries (Bacillariophyceae) as a source of the toxin domoic acid in Algoa Bay, South Africa. Afr. J. Mar. Sci. 2014, 36, 523–528. [Google Scholar] [CrossRef] [Green Version]
- DAFF. South African Live Molluscan Shellfish Monitoring and Control Programme; Republic of South Africa Department of Agriculture, Forestry and Fisheries: Cape Town, South Africa, 2016. [Google Scholar]
- Sá, C.; Leal, M.C.; Silva, A.; Nordez, S.; André, E.; Paula, J.; Brotas, V. Variation of phytoplankton assemblages along the Mozambique coast as revealed by HPLC and microscopy. J. Sea Res. 2013, 79, 1–11. [Google Scholar] [CrossRef]
- Barlow, R.; Stuart, V.; Lutz, V.; Sessions, H.; Sathyendranath, S.; Platt, T.; Kyewalyanga, M.; Clementson, L.; Fukasawa, M.; Watanabe, S.; et al. Seasonal pigment patterns of surface phytoplankton in the subtropical southern hemisphere. Deep Sea Res. Pt. I 2007, 54, 1687–1703. [Google Scholar] [CrossRef]
- Ternon, J.F.; Bach, P.; Barlow, R.; Huggett, J.; Jaquemet, S.; Marsac, F.; Ménard, F.; Penven, P.; Potier, M.; Roberts, M.J. The Mozambique Channel: From physics to upper trophic levels. Deep Sea Res. Pt. II Top. Stud. Oceanogr. 2014, 100, 1–9. [Google Scholar] [CrossRef]
- Tew Kai, E.; Marsac, F. Influence of mesoscale eddies on spatial structuring of top predators’ communities in the Mozambique Channel. Prog. Oceanogr. 2010, 86, 214–223. [Google Scholar] [CrossRef]
- Tew-Kai, E.; Marsac, F. Patterns of variability of sea surface chlorophyll in the Mozambique Channel: A quantitative approach. J. Mar. Syst. 2009, 77, 77–88. [Google Scholar] [CrossRef]
- Fordyce, A. Reef fishes of Praia do Tofo and Praia da Barra, Inhambane, Mozambique. West. Indian O. J. Mar. Sci. 2018, 17, 71–91. [Google Scholar]
- Rohner, C.A.; Richardson, A.J.; Jaine, F.R.A.; Bennett, M.B.; Weeks, S.J.; Cliff, G.; Robinson, D.P.; Reeve-Arnold, K.E.; Pierce, S.J. Satellite tagging highlights the importance of productive Mozambican coastal waters to the ecology and conservation of whale sharks. Peer J. 2018, 6, e4161. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.A.M.; Litulo, C.; Santos, R.; Leal, M.C.; Fernandes, R.S.; Tibiriçá, Y.; Williams, J.; Atanassov, B.; Carreira, F.; Massingue, A.; et al. Mozambique Marine Ecosystems Review; Biodinâmica/CTV: Maputo, Mozambique, 2014; p. 139. [Google Scholar]
- Pierce, S.J.; White, W.T.; Marshall, A.D. New record of the smalleye stingray, Dasyatis microps (Myliobatiformes: Dasyatidae), from the western Indian Ocean. Zootaxa 2008, 1734, 65–68. [Google Scholar] [CrossRef]
- Rohner, C.A.; Pierce, S.J.; Marshall, A.D.; Weeks, S.J.; Bennett, M.B.; Richardson, A.J. Trends in sightings and environmental influences on a coastal aggregation of manta rays and whale sharks. Mar. Ecol. Prog. Ser. 2013, 482, 153–168. [Google Scholar] [CrossRef]
- Williams, J.L.; Pierce, S.J.; Rohner, C.A.; Fuentes, M.M.P.B.; Hamann, M. Spatial distribution and residency of green and loggerhead sea turtles using coastal reef habitats in Southern Mozambique. Front. Mar. Sci. 2017, 3, 228. [Google Scholar] [CrossRef] [Green Version]
- Tibiriçá, Y.; Birtles, A.; Valentine, P.; Miller, D.K. Diving tourism in Mozambique: An opportunity at risk? Tour. Mar. Environ. 2011, 7, 141–151. [Google Scholar] [CrossRef]
- FAO. The Future of Food and Agriculture—Trends and Challenges; FAO: Rome, Italy, 2017. [Google Scholar]
- World Bank. Communities Livelihoods Fisheries: Fisheries Governance and Shared Growth in Mozambique; World Bank: Washington, DC, USA, 2018. [Google Scholar]
- Barlow, R.; Lamont, T.; Gibberd, M.J.; Airs, R.; Jacobs, L.; Britz, K. Phytoplankton communities and acclimation in a cyclonic eddy in the southwest Indian Ocean. Deep Sea Res. Pt. I 2017, 124, 18–30. [Google Scholar] [CrossRef]
- Barlow, R.; Lamont, T.; Morris, T.; Sessions, H.; van den Berg, M. Adaptation of phytoplankton communities to mesoscale eddies in the Mozambique Channel. Deep Sea Res. Pt. I 2014, 100, 106–118. [Google Scholar] [CrossRef]
- Olofsson, M.; Karlberg, M.; Lage, S.; Ploug, H. Phytoplankton community composition and primary production in the tropical tidal ecosystem, Maputo Bay (the Indian Ocean). J. Sea Res. 2017, 125, 18–25. [Google Scholar] [CrossRef]
- Raj, R.P.; Peter, B.N.; Pushpadas, D. Oceanic and atmospheric influences on the variability of phytoplankton bloom in the Southwestern Indian Ocean. J. Mar. Syst. 2010, 82, 217–229. [Google Scholar] [CrossRef]
- Pinckney, J.L.; Millie, D.F.; Howe, K.E.; Paerl, H.W.; Hurley, J.P. Flow scintillation counting of 14C-labeled microalgal photosynthetic pigments. J. Plankton Res. 1996, 18, 1867–1880. [Google Scholar] [CrossRef] [Green Version]
- Mackey, M.D.; Mackey, D.J.; Higgens, H.W.; Wright, S.W. CHEMTAX—A program for estimating class abundances from chemical markers: Application to HPLC measurements of phytoplankton. Mar. Ecol. Prog. Ser. 1996, 144, 265–283. [Google Scholar] [CrossRef] [Green Version]
- Lamont, T.; Barlow, R.G.; Morris, T.; van den Berg, M.A. Characterisation of mesoscale features and phytoplankton variability in the Mozambique Channel. Deep Sea Res. Pt. II Top. Stud. Oceanogr. 2014, 100, 94–105. [Google Scholar] [CrossRef]
- Quartly, G.D.; Srokosz, M.A. Eddies in the southern Mozambique Channel. Deep-Sea Res. Pt. Ii 2004, 51, 69–83. [Google Scholar] [CrossRef] [Green Version]
- Weimerskirch, H.; Le Corre, M.; Jaquemet, S.; Potier, M.; Marsac, F. Foraging strategy of a top predator in tropical waters: Great frigatebirds in the Mozambique Channel. Mar. Ecol. Prog. Ser. 2004, 275, 297–308. [Google Scholar] [CrossRef]
- McCabe, R.M.; Hickey, B.M.; Kudela, R.; Lefebvre, K.; Adams, N.G.; Bill, B.D.; Fulland, F.M.D.; Thomson, R.E.; Cochlan, W.; Trainer, V. An unprecedented coastwide toxic algal bloom linked to anomalous ocean conditions. Geophys. Res. Lett. 2016, 43, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Trainer, V.; Hickey, B.M.; Bates, S.S. Toxic diatoms. In Oceans and Human Health; Walsh, P., Smith, S., Fleming, L.E., Solo-Gabriele, H., Gerwick, W., Eds.; Academic Press: Cambridge, MA, USA, 2008. [Google Scholar]
- Lundholm, N.; Clarke, A.; Ellegaard, M. A 100-year record of changing Pseudo-nitzschia species in a sill-fjord in Denmark related to nitrogen loading and temperature. Harmful Algae 2010, 9, 449–457. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Moisan, C. Shifts and stasis in marine HAB monitoring in New Zealand. Environ. Sci. Pollut. Res. Int. 2013, 20, 6872–6877. [Google Scholar] [CrossRef]
- Smith, J.; Connell, P.; Evans, R.H.; Gellene, A.G.; Howard, M.D.A.; Jones, B.H.; Kaveggia, S.; Palmer, L.; Schnetzer, A.; Seegers, B.N.; et al. A decade and a half of Pseudo-nitzschia spp. and domoic acid along the coast of southern California. Harmful Algae 2018, 79, 87–104. [Google Scholar] [CrossRef]
- Almandoz, G.O.; Ferrario, M.E.; Ferreyra, G.A.; Schloss, I.R.; Esteves, J.L.; Paparazzo, F.E. The genus Pseudo-nitzschia (Bacillariophyceae) in continental shelf waters of Argentina (Southwestern Atlantic Ocean, 38–55° S). Harmful Algae 2007, 6, 93–103. [Google Scholar] [CrossRef]
- Méndez, S.M.; Ferrario, M.; Cefarelli, A.O. Description of toxigenic species of the genus Pseudo-nitzschia in coastal waters of Uruguay: Morphology and distribution. Harmful Algae 2012, 19, 53–60. [Google Scholar] [CrossRef]
- Fernandes, L.F.; Brandini, F. The potentially toxic diatom Pseudo-nitzschia H. Peragallo in the Paraná and Santa Catarina States, Southern Brazil. Ineringia Ser. Bot. 2010, 65, 47–62. [Google Scholar]
- Redfield, A.C. Biological control of chemical factors in the environment. Am. Sci. 1958, 46, 204–221. [Google Scholar]
- Bristow, L.A.; Mohr, W.; Ahmerkamp, S.; Kuypers, M.M.M. Nutrients that limit growth in the ocean. Curr. Biol. 2017, 27, R474–R478. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, C.S. Ecology of Phytoplankton; Cambridge University Press: New York, NY, USA, 2006. [Google Scholar]
- Parsons, M.L.; Dortch, Q.; Turner, R.E. Sedimentological evidence of an increase in Pseudo-nitzschia (Bacillariophyceae) abundance in response to coastal eutrophication. Limnol. Oceanogr. 2002, 47, 551–558. [Google Scholar] [CrossRef]
- Martiny, A.C.; Pham, C.T.A.; Primeau, F.W.; Vrugt, J.A.; Moore, J.K.; Levin, S.A.; Lomas, M.W. Strong latitudinal patterns in the elemental ratios of marine plankton and organic matter. Nat. Geosci. 2013, 6, 279–283. [Google Scholar] [CrossRef]
- Jones, R.H.; Flynn, K.J. Nutritional status and diet composition affect the value of diatoms as copepod prey. Science 2005, 307, 1457–1459. [Google Scholar] [CrossRef] [PubMed]
- Sterner, R.W.; Schultz, K.L. Zooplankton nutrition: Recent progress and a reality check. Aquat. Ecol. 1998, 32, 261–279. [Google Scholar] [CrossRef]
- Cohen, N.R.; Gong, W.; Moran, D.M.; McIlvin, M.R.; Saito, M.A.; Marchetti, A. Transcriptomic and proteomic responses of the oceanic diatom Pseudo-nitzschia granii to iron limitation. Environ. Microbiol. 2018, 20, 3109–3126. [Google Scholar] [CrossRef]
- Pan, Y.; Bates, S.S.; Cembella, A.D. Environmental stress and domoic acid production by Pseudo-nitzschia: A physiological perspective. Nat. Toxins 1998, 6, 127–135. [Google Scholar] [CrossRef]
- Hu, Z.-X.; Xu, N.; Aifen, L.; Duan, S. Effects of different N:P ratios on the growth of Pseudo-nitzschia pungens, Prorocentrum donghaiense and Phaeocystis globosa. Acta Hydrobiol. Sin. 2008, 32, 482–487. [Google Scholar] [CrossRef]
- Quilliam, M.A.; Sim, P.G.; McCulloch, A.W.; McInnes, A.G. High-performance liquid chromatography of domoic acid, a marine neurotoxin, with application to shellfish and plankton. Int. J. Environ. Anal. Chem. 2006, 36, 139–154. [Google Scholar] [CrossRef]
- Wohlgeschaffen, G.D.; Mann, K.H.; Subba Rao, D.V.; Pocklington, R. Dynamics of the phycotoxin domoic acid: Accumulation and excretion in two commercially important bivalves. J. Appl. Phycol. 1992, 4, 297–310. [Google Scholar] [CrossRef]
- Petroff, R.; Hendrix, A.; Shum, S.; Grant, K.S.; Lefebvre, K.A.; Burbacher, T.M. Public health risks associated with chronic, low-level domoic acid exposure: A review of the evidence. Pharmacol. Ther. 2021, 227, 107865. [Google Scholar] [CrossRef]
- Ferriss, B.E.; Marcinek, D.J.; Ayres, D.; Borchert, J.; Lefebvre, K.A. Acute and chronic dietary exposure to domoic acid in recreational harvesters: A survey of shellfish consumption behavior. Environ. Int. 2017, 101, 70–79. [Google Scholar] [CrossRef]
- Hiolski, E.M.; Kendrick, P.S.; Frame, E.R.; Myers, M.S.; Bammler, T.K.; Beyer, R.P.; Farin, F.M.; Wilkerson, H.W.; Smith, D.R.; Marcinek, D.J.; et al. Chronic low-level domoic acid exposure alters gene transcription and impairs mitochondrial function in the CNS. Aquat. Toxicol. 2014, 155, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, K.A.; Kendrick, P.S.; Ladiges, W.; Hiolski, E.M.; Ferriss, B.E.; Smith, D.R.; Marcinek, D.J. Chronic low-level exposure to the common seafood toxin domoic acid causes cognitive deficits in mice. Harmful Algae 2017, 64, 20–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husson, B.; Hernandez-Farinas, T.; Le Gendre, R.; Schapira, M.; Chapelle, A. Two decades of Pseudo-nitzschia spp. blooms and king scallop (Pecten maximus) contamination by domoic acid along the French Atlantic and English Channel coasts: Seasonal dynamics, spatial heterogeneity and interannual variability. Harmful Algae 2016, 51, 26–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekula-Wood, E.; Benitez-Nelson, C.; Morton, S.; Anderson, C.; Burrell, C.; Thunell, R. Pseudo-nitzschia and domoic acid fluxes in Santa Barbara Basin (CA) from 1993 to 2008. Harmful Algae 2011, 10, 567–575. [Google Scholar] [CrossRef]
- Lugomela, C. Population dynamics of Pseudo-nitzschia species (Bacillariophyceae) in the near shore waters of Dar Es Salaam, Tanzania. Tanz. J. Sci. 2013, 39, 38–48. [Google Scholar]
- Hermes, J.C.; Masumoto, Y.; Beal, L.M.; Roxy, M.K.; Vialard, J.; Andres, M.; Annamalai, H.; Behera, S.; D’Adamo, N.; Doi, T.; et al. A sustained ocean observing system in the Indian Ocean for climate related scientific knowledge and societal needs. Front. Mar. Sci. 2019, 6, 355. [Google Scholar] [CrossRef]
- Lugomela, C.; Lyimo, T.J.; Bryceson, I.; Semesi, A.K.; Bergman, B. Trichodesmium in coastal waters of Tanzania: Diversity, seasonality, nitrogen and carbon fixation. Hydrobiologia 2002, 477, 1–13. [Google Scholar] [CrossRef]
- Backeberg, B.C.; Reason, C.J.C. A connection between the South Equatorial Current north of Madagascar and Mozambique Channel Eddies. Geophys. Res. Lett. 2010, 37, L04604. [Google Scholar] [CrossRef] [Green Version]
- Madhu, N.V.; Maheswaran, P.A.; Jyothibabu, R.; Sunil, V.; Revichandran, C.; Balasubramanian, T.; Gopalakrishnan, T.C.; Nair, K.K.C. Enhanced biological production off Chennai triggered by October 1999 super cyclone (Orissa). Curr. Sci. 2002, 82, 1472–1479. [Google Scholar]
- Uz, B.M. What causes the sporadic phytoplankton bloom southeast of Madagascar? J. Geophys. Res. 2007, 112, 1129. [Google Scholar] [CrossRef] [Green Version]
- Hamisi, M.I.; Mamboya, F.A. Nutrient and phytoplankton dynamics along the Ocean Road Sewage Discharge Channel, Dar es Salaam, Tanzania. J. Ecosys. 2014, 2014, 271456. [Google Scholar] [CrossRef]
- Saji, N.H.; Goswami, B.N.; Vinayachandran, P.N.; Yamagata, T. A dipole mode in the tropical Indian Ocean. Nature 1999, 401, 360–363. [Google Scholar] [CrossRef]
- Usman, M.T.; Reason, C.J.C. Dry spell frequencies and their variability over southern Africa. Climate Res. 2004, 26, 199–211. [Google Scholar] [CrossRef] [Green Version]
- Porter, A.; Bohl, D.; Kwasi, S.; Donnenfeld, Z.; Cilliers, J. Prospects and Challenges: Mozambique’s Growth and Human Development Outlook to 2040; Institute for Security Studies,: Pretoria, South Africa, 2017. [Google Scholar]
- FAO. National Aquaculture Sector Overview: Mozambique; FAO: Rome, Italy, 2005. [Google Scholar]
- Bryden, H.L.; Beal, L.M.; Duncan, L.M. Structure and transport of the Agulhas Current and its temporal variability. J. Oceanogr. 2005, 61, 479–492. [Google Scholar] [CrossRef]
- Lamont, T.; Roberts, M.J.; Barlow, R.G.; Morris, T.; van den Berg, M.A. Circulation patterns in the Delagoa Bight, Mozambique, and the influence of deep ocean eddies. Afr. J. Mar. Sci. 2010, 32, 553–562. [Google Scholar] [CrossRef]
- Lutjeharms, J.R.E. The ocean environment off southeastern Africa: A review. S. Afr. J. Sci. 2006, 102, 419–426. [Google Scholar]
- Swart, N.C.; Lutjeharms, J.R.E.; Ridderinkhof, H.; de Ruijter, W.P.M. Observed characteristics of Mozambique Channel eddies. J. Geophys. Res. 2010, 115, C09006. [Google Scholar] [CrossRef] [Green Version]
- Westberry, T.K.; Siegel, D.A. Spatial and temporal distribution of Trichodesmium blooms in the world’s oceans. Glob. Biogeochem. Cy. 2006, 20, GB4016. [Google Scholar] [CrossRef]
- Goreau, T.F.; Goreau, N.I.; Goreau, T.J. Corals and coral reefs. Sci. Am. 1979, 241, 124–137. [Google Scholar] [CrossRef]
- Bakun, A. Coastal upwelling indices, west coast of North America. Spec. Sci. Rep. Fish. Ser. 1973, 231, 1946–1971. [Google Scholar]
- Gonzalez-Nuevo, G.; Gago, J.; Cabanas, J.M. Upwelling index: A powerful tool for marine research in the NW Iberian upwelling system. J. Oper. Oceano. 2014, 7, 47–57. [Google Scholar] [CrossRef]
- Lavín, A.; Díaz del Río, G.; Cabanas, J.M.; Casas, G. Afloramiento en el Noroeste de la Peninsula Iberica Indices de Afloramiento Para El Punto 43.0N–11.0W Periodo 1966-1989; Instituto Español de Oceanografia: Madrid, Spain, 1991. [Google Scholar]
- Utermöhl, H. Toward the improvement of the quantitative phytoplankton method. Mitt. Int. Verieningung Fur Limnol. 1958, 9, 1–39. [Google Scholar]
- Bianchi, T.S.; Bauer, J.E. Particulate organic carbon cycling and transformation. In Treatise on Estuarine and Coastal Science; Academic Press: Cambridge, MA, USA, 2011; pp. 69–117. [Google Scholar]
- Van Heukelem, L.; Lewitus, A.J.; Kana, T.M.; Craft, N.E. Improved separations of phytoplankton pigments using temperature-controlled high performance liquid chromatography. Mar. Ecol. Prog. Ser. 1994, 114, 303–313. [Google Scholar] [CrossRef]
- Latasa, M. Improving estimations of phytoplankton class abundances using CHEMTAX. Mar. Ecol. Prog. Ser. 2007, 329, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Higgins, H.W.; Wright, S.W.; Schluter, L. Quantitative interpretation of chemotaxonomic pigment data. In Phytoplankton Pigments: Characterization, Chemotaxonomy, and Applications in Oceanography; Roy, S., Llewellyn, C.A., Eds.; Cambridge University Press: New York, NY, USA, 2011; pp. 257–313. [Google Scholar]
- Van der Molen, J.S.; Scharler, U.M.; Muir, D. Species composition, abundance and biomass of microphytoplankton in the KwaZulu-Natal Bight on the east coast of South Africa. Afr. J. Mar. Sci. 2016, 38, S139–S153. [Google Scholar] [CrossRef]
- Harris, R.P.; Wiebe, P.H.; Lenz, J.; Skjoldal, H.R.; Huntly, M. Zooplankton Methodology Manual; Academic Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Baustian, M.M.; Bargu, S.; Morrison, W.; Sexton, C.; Rabalais, N.N. The polychaete, Paraprionospio pinnata, is a likely vector of domoic acid to the benthic food web in the northern Gulf of Mexico. Harmful Algae 2018, 79, 44–49. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kelchner, H.; Reeve-Arnold, K.E.; Schreiner, K.M.; Bargu, S.; Roques, K.G.; Errera, R.M. Domoic Acid and Pseudo-nitzschia spp. Connected to Coastal Upwelling along Coastal Inhambane Province, Mozambique: A New Area of Concern. Toxins 2021, 13, 903. https://doi.org/10.3390/toxins13120903
Kelchner H, Reeve-Arnold KE, Schreiner KM, Bargu S, Roques KG, Errera RM. Domoic Acid and Pseudo-nitzschia spp. Connected to Coastal Upwelling along Coastal Inhambane Province, Mozambique: A New Area of Concern. Toxins. 2021; 13(12):903. https://doi.org/10.3390/toxins13120903
Chicago/Turabian StyleKelchner, Holly, Katie E. Reeve-Arnold, Kathryn M. Schreiner, Sibel Bargu, Kim G. Roques, and Reagan M. Errera. 2021. "Domoic Acid and Pseudo-nitzschia spp. Connected to Coastal Upwelling along Coastal Inhambane Province, Mozambique: A New Area of Concern" Toxins 13, no. 12: 903. https://doi.org/10.3390/toxins13120903
APA StyleKelchner, H., Reeve-Arnold, K. E., Schreiner, K. M., Bargu, S., Roques, K. G., & Errera, R. M. (2021). Domoic Acid and Pseudo-nitzschia spp. Connected to Coastal Upwelling along Coastal Inhambane Province, Mozambique: A New Area of Concern. Toxins, 13(12), 903. https://doi.org/10.3390/toxins13120903