Acute Oral Toxicity of Pinnatoxin G in Mice

 , , , ,
, , , ,  and
and 
Abstract
1. Introduction
2. Results
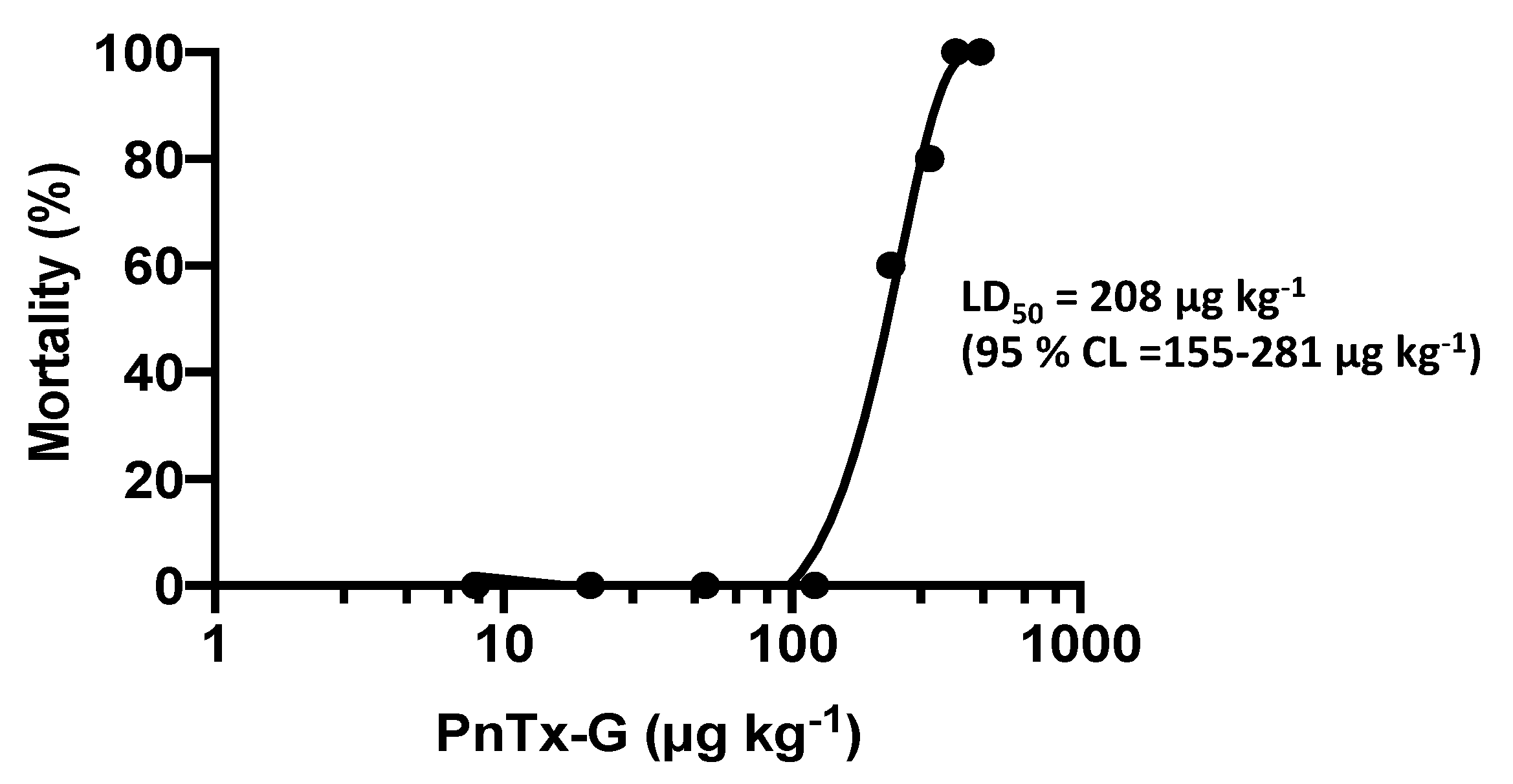
2.1. Lethality and Clinical Signs
2.2. Blood Chemistry
2.3. Necropsy
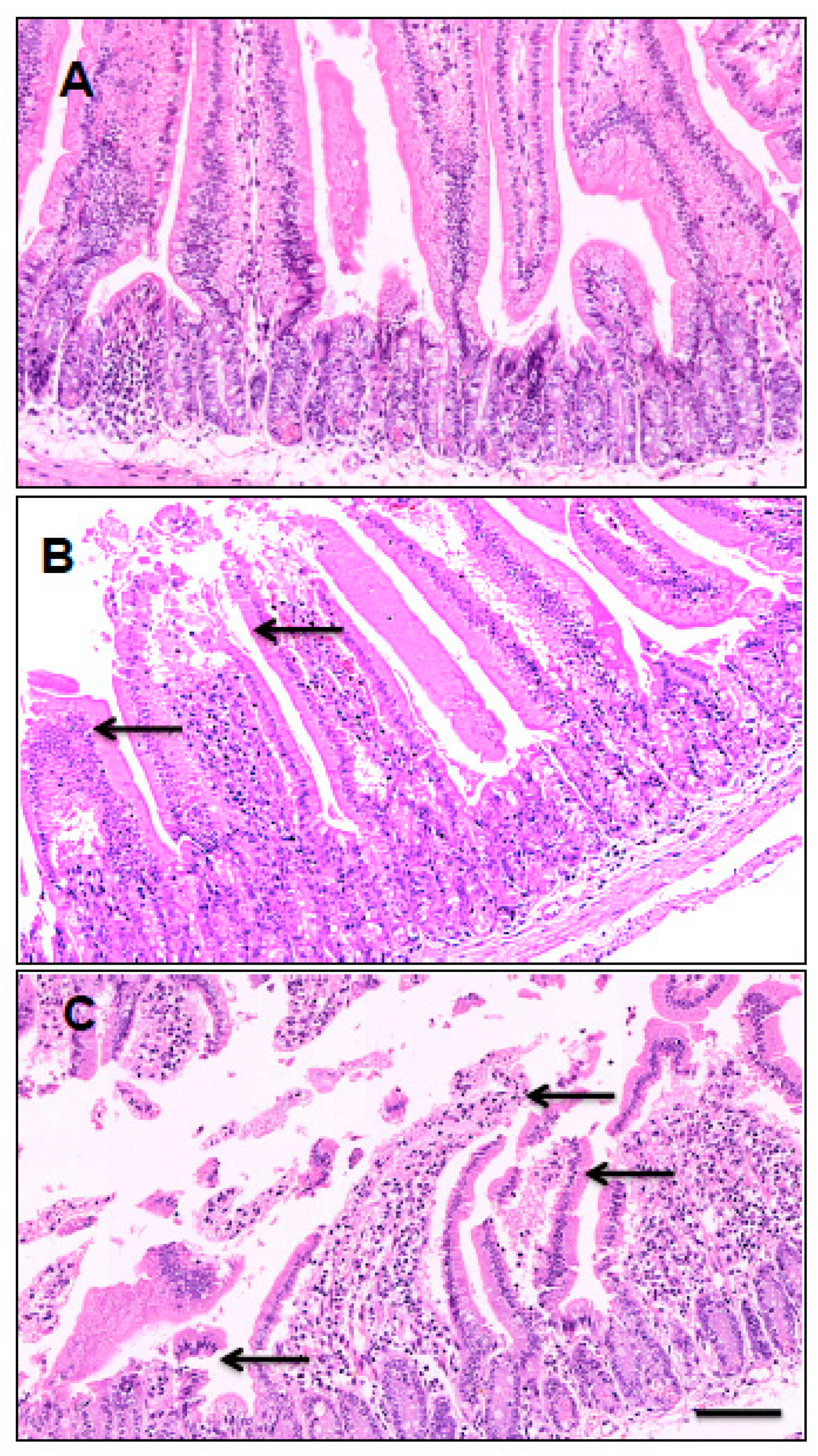
2.4. Histological Analysis
3. Discussion
4. Materials and Methods
4.1. Toxin, Reagents, and Other Chemicals
4.2. Animals and Experimental Conditions
4.3. Experimental Design
4.4. Blood Chemistry
4.5. Histological Analysis
4.6. Statistical Analysis and Determination of LD50
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zheng, S.; Huang, F.; Chen, S.; Tan, X.; Zuo, J.; Peng, J.; Xie, R. The isolation and bioactivities of pinnatoxin. Chin. J. Mar. Drugs 1990, 9, 33–35. [Google Scholar]
- Uemura, D.; Chou, T.; Haino, T.; Nagatsu, A.; Fukuzawa, S.; Zheng, S.Z.; Chen, H.S. Pinnatoxin-A a toxic amphoteric macrocycle from the okinawan bivalve Pinna muricata. J. Am. Chem. Soc. 1995, 117, 1155–1156. [Google Scholar] [CrossRef]
- Chou, T.; Haino, T.; Kuramoto, M.; Uemura, D. Isolation and structure of pinnatoxin D, a new shellfish poison from the okinawan bivalve Pinna muricata. Tetrahedron Lett. 1996, 37, 4027–4030. [Google Scholar] [CrossRef]
- Takada, N.; Umemura, N.; Suenaga, K.; Chou, T.; Nagatsu, A.; Haino, T.; Yamada, K.; Uemura, D. Pinnatoxins B and C, the most toxic components in the pinnatoxin series from the okinawan bivalve Pinna muricata. Tetrahedron Lett. 2001, 42, 3491–3494. [Google Scholar] [CrossRef]
- Selwood, A.I.; Miles, C.O.; Wilkins, A.L.; van Ginkel, R.; Munday, R.; Rise, F.; McNabb, P. Isolation, structural determination and acute toxicity of pinnatoxins E, F and G. J. Agric. Food Chem. 2010, 58, 6532–6542. [Google Scholar] [CrossRef]
- Selwood, A.I.; Wilkins, A.L.; Munday, R.; Gu, H.; Smith, K.F.; Rhodes, L.L.; Rise, F. Pinnatoxin H: A new pinnatoxin analogue from a South China Sea Vulcanodinium rugosum isolate. Tetrahedron Lett. 2014, 55, 5508–5510. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Selwood, A.; McNabb, P.; van Ginkel, R.; Holland, P.; Munday, R. Production of pinnatoxins by a peridinoid dinoflagellate isolated from Northland, New Zealand. Harmful Algae 2010, 9, 384–389. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Selwood, A.; McNabb, P.; Molenaar, S.; Munday, R.; Wilkinson, C.; Hallegraeff, G. Production of pinnatoxins E, F and G by scrippsielloid dinoflagellates isolated from Franklin Harbour, South Australia. N. Z. J. Mar. Freshw. Res. 2011, 45, 703–709. [Google Scholar] [CrossRef]
- Smith, K.F.; Rhodes, L.L.; Suda, S.; Selwood, A.I. A dinoflagellate producer of pinnatoxin G, isolated from sub-tropical Japanese waters. Harmful Algae 2011, 10, 702–705. [Google Scholar] [CrossRef]
- Rundberget, T.; Aasen, J.A.; Selwood, A.I.; Miles, C.O. Pinnatoxins and spirolides in Norwegian blue mussels and seawater. Toxicon 2011, 58, 700–711. [Google Scholar] [CrossRef] [PubMed]
- McCarron, P.; Rourke, W.A.; Hardstaff, W.; Pooley, B.; Quilliam, M.A. Identification of pinnatoxins and discovery of their fatty acid ester metabolites in mussels (Mytilus edulis) from eastern Canada. J. Agric. Food Chem. 2012, 60, 1437–1446. [Google Scholar] [CrossRef] [PubMed]
- Geiger, M.; Desanglois, G.; Hogeveen, K.; Fessard, V.; Leprêtre, T.; Mondeguer, F.; Guitton, Y.; Hervé, F.; Séchet, V.; Grovel, O.; et al. Cytotoxicity, fractionation and dereplication of extracts of the dinoflagellate Vulcanodinium rugosum, a producer of pinnatoxin G. Mar. Drugs 2013, 11, 3350–3371. [Google Scholar] [CrossRef] [PubMed]
- Hess, P.; Abadie, E.; Hervé, F.; Berteaux, T.; Séchet, V.; Aráoz, R.; Molgó, J.; Zakarian, A.; Sibat, M.; Rundberget, T. Pinnatoxin G is responsible for atypical toxicity in mussels Mytilus galloprovincialis and clams Venerupis decussata from Ingril, a French Mediterranean lagoon. Toxicon 2013, 75, 16–26. [Google Scholar] [CrossRef] [PubMed]
- García-Altares, M.; Casanova, A.; Bane, V.; Diogène, J.; Furey, A.; de la Iglesia, P. Confirmation of pinnatoxins and spirolides in shellfish and passive samplers from Catalonia (Spain) by liquid chromatography coupled with triple quadrupole and high-resolution hybrid tandem mass spectrometry. Mar. Drugs 2014, 12, 3706–3732. [Google Scholar] [CrossRef]
- McCarthy, M.; Bane, V.; García-Altares, M.; van Pelt, F.N.; Furey, A.; O’Halloran, J. Assessment of emerging biotoxins (pinnatoxin G and spirolides) at Europe’s first marine reserve: Lough Hyne. Toxicon 2015, 108, 202–209. [Google Scholar] [CrossRef]
- Al Muftah, A.; Selwood, A.I.; Foss, A.J.; Al-Jabri, H.M.; Potts, M.; Yilmaz, M. Algal toxins and producers in the marine waters of Qatar, Arabian Gulf. Toxicon 2016, 122, 54–66. [Google Scholar] [CrossRef]
- Rambla-Alegre, M.; Miles, C.O.; de la Iglesia, P.; Fernandez-Tejedor, M.; Jacobs, S.; Sioen, I.; Verbeke, W.; Samdal, I.A.; Sandvik, M.; Barbosa, V.; et al. Occurrence of cyclic imines in European commercial seafood and consumers risk assessment. Environ. Res. 2018, 161, 392–398. [Google Scholar] [CrossRef]
- Otero, P.; Miguéns, N.; Rodríguez, I.; Botana, L.M. LC-MS/MS analysis of the emerging toxin pinnatoxin-G and high levels of esterified OA group toxins in Galician commercial mussels. Toxins 2019, 11, 394. [Google Scholar] [CrossRef]
- Lamas, J.P.; Arévalo, F.; Moroño, Á.; Correa, J.; Muñíz, S.; Blanco, J. Detection and spatio-temporal distribution of pinnatoxins in shellfish from the Atlantic and Cantabrian coasts of Spain. Toxins 2019, 11, 340. [Google Scholar] [CrossRef]
- Nézan, E.; Chomerat, N. Vulcanodinium rugosum gen. Nov., sp. Nov. (dinophyceae): A new marine dinoflagellate from the French mediterranean coast. Cryptogam. Algol. 2011, 32, 3–18. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Selwood, A.; McNabb, P.; Munday, R.; Suda, S.; Molenaar, S.; Hallegraeff, G. Dinoflagellate Vulcanodinium rugosum identified as the causative organism of pinnatoxins in Australia, New Zealand and Japan. Phycologia 2011, 50, 624–628. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); Panel on contaminants in the food chain (CONTAM). Scientific opinion on marine biotoxins in shellfish-cyclic imines (spirolides, gymnodimines, pinnatoxins and pteriatoxins). EFSA J. 2010, 8, 1628–1667. [Google Scholar] [CrossRef]
- Araoz, R.; Servent, D.; Molgó, J.; Iorga, B.I.; Fruchart-Gaillard, C.; Benoit, E.; Gu, Z.; Stivala, C.; Zakarian, A. Total synthesis of pinnatoxins A and G and revision of the mode of action of pinnatoxin A. J. Am. Chem. Soc. 2011, 133, 10499–10511. [Google Scholar] [CrossRef]
- Hellyer, S.D.; Selwood, A.I.; Rhodes, L.; Kerr, D.S. Marine algal pinnatoxins E and F cause neuromuscular block in an in vitro hemidiaphragm prepration. Toxicon 2011, 58, 693–699. [Google Scholar] [CrossRef]
- Hellyer, S.D.; Selwood, A.I.; Rhodes, L.; Kerr, D.S. Neuromuscular blocking activity of pinnatoxins E, F and G. Toxicon 2013, 76, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Hellyer, S.D.; Indurthi, D.; Balle, T.; Runder-Varga, V.; Selwood, A.I.; Tyndall, J.D.; Chebib, M.; Rhodes, L.; Kerr, D.S. Pinnatoxins E, F and G target multiple nicotinic receptor subtypes. J. Neurochem. 2015, 135, 479–491. [Google Scholar] [CrossRef]
- Bourne, Y.; Sulzenbacher, G.; Radić, Z.; Aráoz, R.; Reynaud, M.; Benoit, E.; Zakarian, A.; Servent, D.; Molgó, J.; Taylor, P.; et al. Marine macrocyclic imines, pinnatoxins A and G: Structural determinants and functional properties to distinguish neuronal α7 from muscle α1(2)βγδ nAChRs. Structure 2015, 23, 1106–1115. [Google Scholar] [CrossRef]
- Molgó, J.; Marchot, P.; Aráoz, R.; Benoit, E.; Iorga, B.I.; Zakarian, A.; Taylor, P.; Bourne, Y.; Servent, D. Cyclic imine toxins from dinoflagellates: A growing family of potent antagonists of the nicotinic acetylcholine receptors. J. Neurochem. 2017, 142 (Suppl. 2), 41–51. [Google Scholar] [CrossRef]
- Delcourt, N.; Lagrange, E.; Abadie, E.; Fessard, V.; Frémy, J.-M.; Vernoux, J.-P.; Peyrat, M.-B.; Maignien, T.; Arnich, N.; Molgó, J.; et al. Pinnatoxins’ deleterious effects on cholinergic networks: From experimental models to human health. Mar. Drugs 2019, 17, 425. [Google Scholar] [CrossRef]
- Benoit, E.; Couesnon, A.; Lindovsky, J.; Iorga, B.I.; Aráoz, R.; Servent, D.; Zakarian, A.; Molgó, J. Synthetic pinnatoxins A and G reversibly block mouse skeletal neuromuscular transmission in vivo and in vitro. Mar. Drugs 2019, 17, 306. [Google Scholar] [CrossRef]
- Munday, R.; Selwood, A.I.; Rhodes, L. Acute toxicity of pinnatoxins E, F and G to mice. Toxicon 2012, 60, 995–999. [Google Scholar] [CrossRef] [PubMed]
- Molgó, J.; Benoit, E.; Aráoz, R.; Zakarian, A.; Iorga, B.I. Spirolides and Cyclic Imines: Toxicological Profile. Marine and Freshwater Toxins 2016, Volume 3, 193–217. [Google Scholar] [CrossRef]
- Organisation for Economic Cooperation and Development (OECD). Guidelines for Testing of Chemicals. 425. Acute Oral Toxicity—Up-and-Down-Procedure (UDP). Available online: https://ntp.niehs.nih.gov/iccvam/suppdocs/feddocs/oecd/oecdtg425.pdf (accessed on 13 December 2019).
- Assessment and Management of Biotoxin Risks in Bivalve Molluscs. Available online: http://www.fao.org/3/i2356e/i2356e.pdf (accessed on 13 December 2019).
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971; pp. 1–333. Available online: https://dspace.gipe.ac.in/xmlui/bitstream/handle/10973/36028/GIPE-025784.pdf?sequence=3 (accessed on 28 January 2020).



{kind=link}
{kind=link}
| Group of Treatment | Dose (µg kg−1) | Lethality | Survival Times (min) | Signs of Toxicity |
|---|---|---|---|---|
| Controls | - | 0/8 | - | - |
| PnTx-G | 8 | 0/3 | - | - |
| 20 | 0/3 | - | - | |
| 50 | 0/3 | - | - | |
| 120 | 0/3 | - | - | |
| 220 | 3/5 | 20-22-22 | Prostration, tremors, jumping, abdominal breathing, paralysis of the hind limbs, cyanosis | |
| 300 | 4/5 | 12-13-17-23 | Prostration, tremors, jumping, abdominal breathing, paralysis of the hind limbs, cyanosis | |
| 370 | 5/5 | 13-15-16-17-18 | Prostration, tremors, jumping, abdominal breathing, paralysis of the hind limbs, cyanosis | |
| 450 | 5/5 | 12-12-15-16-29 | Prostration, tremors, jumping, abdominal breathing, paralysis of the hind limbs, cyanosis |
| Parameter | Controls (n = 8) | PnTx-G 8 μg kg−1 (n = 3) | PnTx-G 20 μg kg−1 (n = 3) | PnTx-G 50 μg kg−1 (n = 3) | PnTx-G 120 μg kg−1 (n = 3) | PnTx-G 220 μg kg−1 (n = 3) | PnTx-G 300 μg kg−1 (n = 5) | PnTx-G 370 μg kg−1 (n = 3) | PnTx-G 450 μg kg−1 (n = 4) |
|---|---|---|---|---|---|---|---|---|---|
| ALT (IU/L) | 58.1 ± 11.0 | 34.7 ± 9.6 (−40%) | 37.7 ± 5.4 (−35%) | 34.0 ± 6.1 (−41%) | 29.3 ± 4.7 (−50%) | 65.5 ± 14.9 (13%) | 63.3 ± 14.0 (9%) | 43.7 ± 4.7 (−25%) | 63.3 ± 13.6 (9%) |
| AST (IU/L) | 77.8 ± 8.4 | 80.7 ± 29.2 (4%) | 78.7 ± 11.7 (1%) | 88.0 ± 11.8 (13%) | 93.3 ± 28.8 (20%) | 95.3 ± 16.0 (22%) | 97.3 ± 14.6 (25%) | 96.3 ± 8.6 (24%) | 104.3 ± 23.4 (34%) |
| GLDH (IU/L) | 46.8 ± 12.8 | 40.5 ± 3.8 (−13%) | 33.4 ± 2.7 (−27%) | 31.1 ± 0.3 (−34%) | 35.0 ± 3.0 (−25%) | 52.8 ± 11.8 (13%) | 25.2 ± 6.0 (−46%) | 35.2 ± 3.0 (−25%) | 48.9 ± 14.1 (5%) |
| CPK (IU/L) | 1158.2 ± 212.8 | 597.0 ± 199.8 (−48%) | 1365.3 ± 639.1 (18%) | 670.7 ± 114.2 (−42%) | 1381.0 ± 54.0 (19%) | 1126.3 ± 290.6 (−3%) | 1245.0 ± 330.6 (7%) | 1176.0 ± 100.7 (2%) | 1131.8 ± 138.8 (−2%) |
| Creatinine (mg/dL) | 28.1 ± 1.3 | 27.7 ± 0.9 (−1%) | 26.0 ± 1.5 (−7%) | 25.7 ± 1.9 (−9%) | 25.7 ± 1.8 (−9%) | 40.6 ± 12.9 (47%) | 24.7 ± 1.2 (−12%) | 27.7 ± 2.0 (1%) | 33.3 ± 3.4 (19%) |
| Ca2+ (mM) | 2.9 ± 0.1 | 2.5 ± 0.1 (−14%) | 2.6 ± 0.2 (−10%) | 2.6 ± 0.1 (−10%) | 2.5 ± 0.1 (−14%) | 3.1 ± 0.2 (7%) | 2.8 ± 0.2 (−3%) | 3.2 ± 0.3 (10%) | 3.2 ± 0.3 (10%) |
| Na+ (mM) | 136.3 ± 1.5 | 137.5 ± 0.8 (1%) | 133.8 ± 1.5 (−2%) | 134.2 ± 1.6 (−2%) | 136.3 ± 2.3 (0%) | 136.3 ± 5.9 (0%) | 124.1 ± 9.4 (−9%) | 138.2 ± 0.6 (1%) | 133.9 ± 3.9 (−3%) |
| K+ (mM) | 9.3 ± 0.8 | 6.6 ± 0.3 (−29%) | 8.4 ± 0.9 (−10%) | 7.6 ± 0.2 (−18%) | 7.3 ± 0.8 (−47%) | 10.4 ± 1.8 (12%) | 7.2 ± 1.3 (−23%) | 7.1 ± 3.7 (8%) | 10.3 ± 0.4 (−11%) |
| Cl− (mM) | 103.2 ± 1.0 | 103.6 ± 1.8 (1%) | 101.9 ± 1.0 (−2%) | 101.0 ± 2.2 (−2%) | 103.7 ± 2.3 (1%) | 104.8 ± 2.6 (2%) | 102.6 ± 4.3 (−1%) | 107.4 ± 1.1 (4%) | 102.9 ± 2.8 (−1%) |
| Pi (mM) | 3.9 ± 0.2 | 3.2 ± 0.2 (−18%) | 3.6 ± 0.2 (−8%) | 3.4 ± 0.2 (−13%) | 3.2 ± 0.3 (−18%) | 4.0 ± 0.4 (3%) | 4.3 ± 0.3 (10%) | 4.5 ± 0.4 (15%) | 4.5 ± 0.8 (15%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sosa, S.; Pelin, M.; Cavion, F.; Hervé, F.; Hess, P.; Tubaro, A. Acute Oral Toxicity of Pinnatoxin G in Mice. Toxins 2020, 12, 87. https://doi.org/10.3390/toxins12020087
Sosa S, Pelin M, Cavion F, Hervé F, Hess P, Tubaro A. Acute Oral Toxicity of Pinnatoxin G in Mice. Toxins. 2020; 12(2):87. https://doi.org/10.3390/toxins12020087
Chicago/Turabian StyleSosa, Silvio, Marco Pelin, Federica Cavion, Fabienne Hervé, Philipp Hess, and Aurelia Tubaro. 2020. "Acute Oral Toxicity of Pinnatoxin G in Mice" Toxins 12, no. 2: 87. https://doi.org/10.3390/toxins12020087
APA StyleSosa, S., Pelin, M., Cavion, F., Hervé, F., Hess, P., & Tubaro, A. (2020). Acute Oral Toxicity of Pinnatoxin G in Mice. Toxins, 12(2), 87. https://doi.org/10.3390/toxins12020087