Comparison of the Protective Effects of Bee Venom Extracts with Varying PLA2 Compositions in a Mouse Model of Parkinson’s Disease


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Purification PLA2 and Melittin from Crude Bee Venom
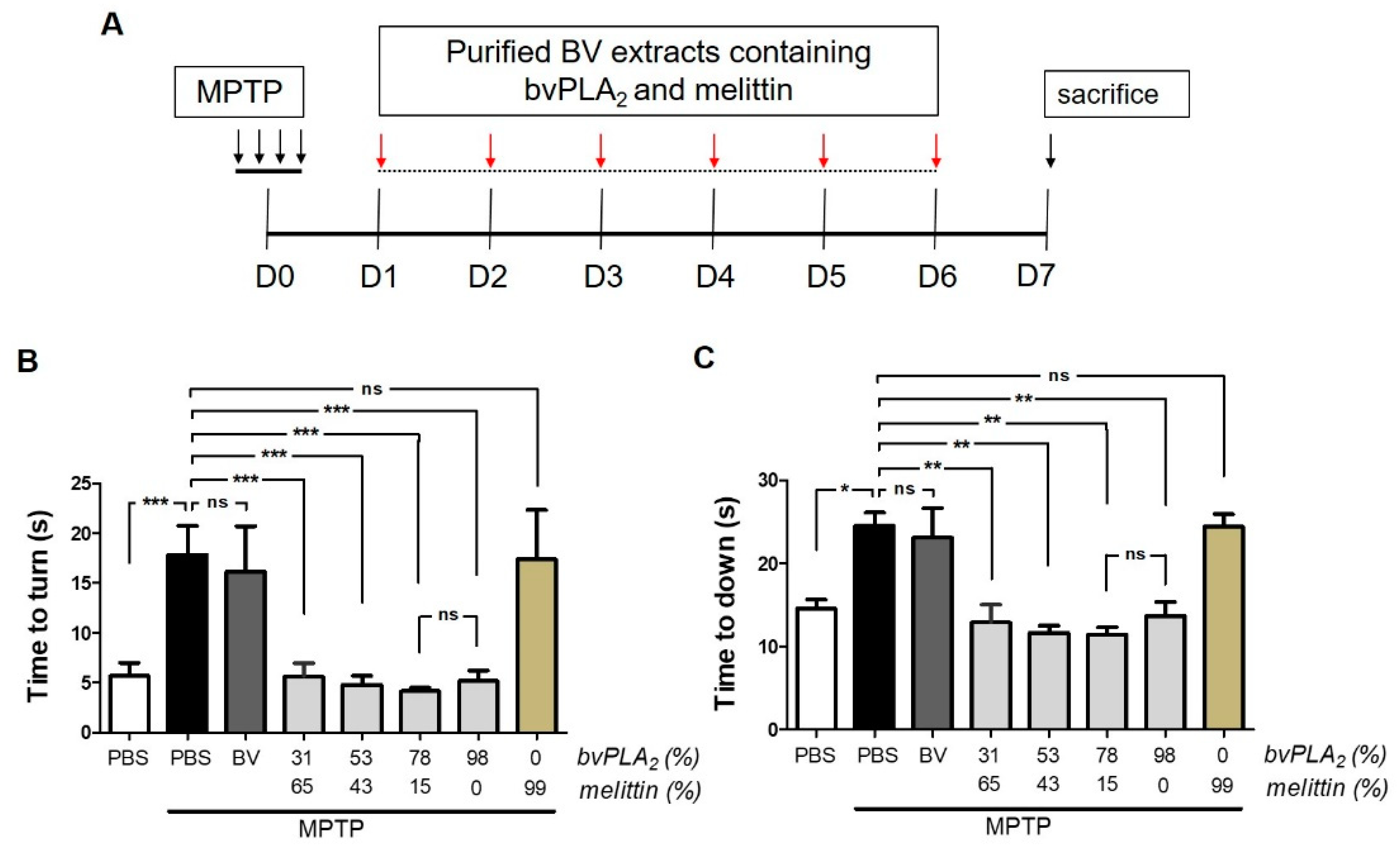
2.2. BvPLA2 with Enriched PLA2 Levels Protects against Neurotoxicity in Parkinson’s Disease in MPTP-Induced Mice
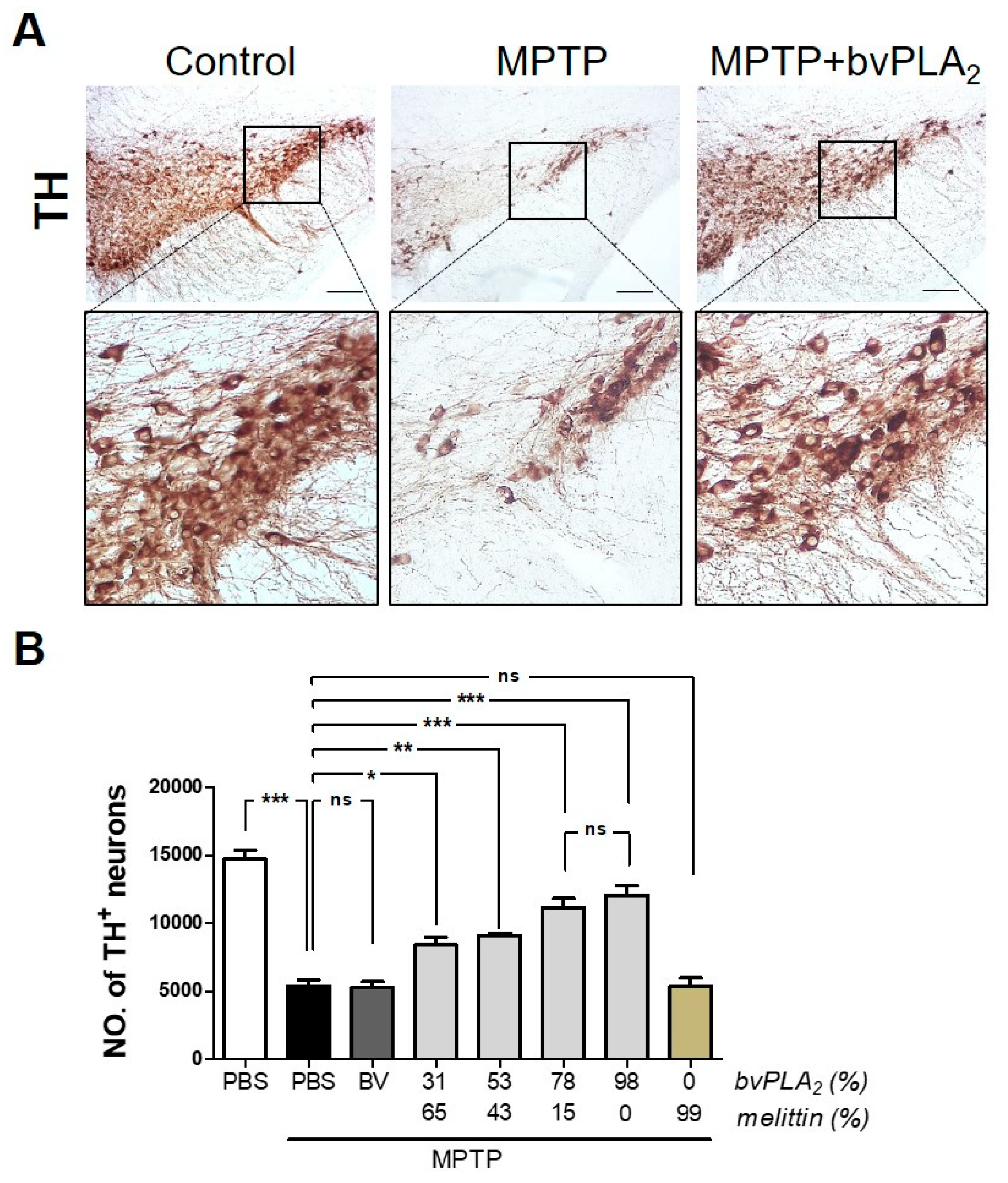
2.3. BvPLA2 with Enriched PLA2 Rescues Loss of Dopaminergic Neurons in MPTP-Treated Mice
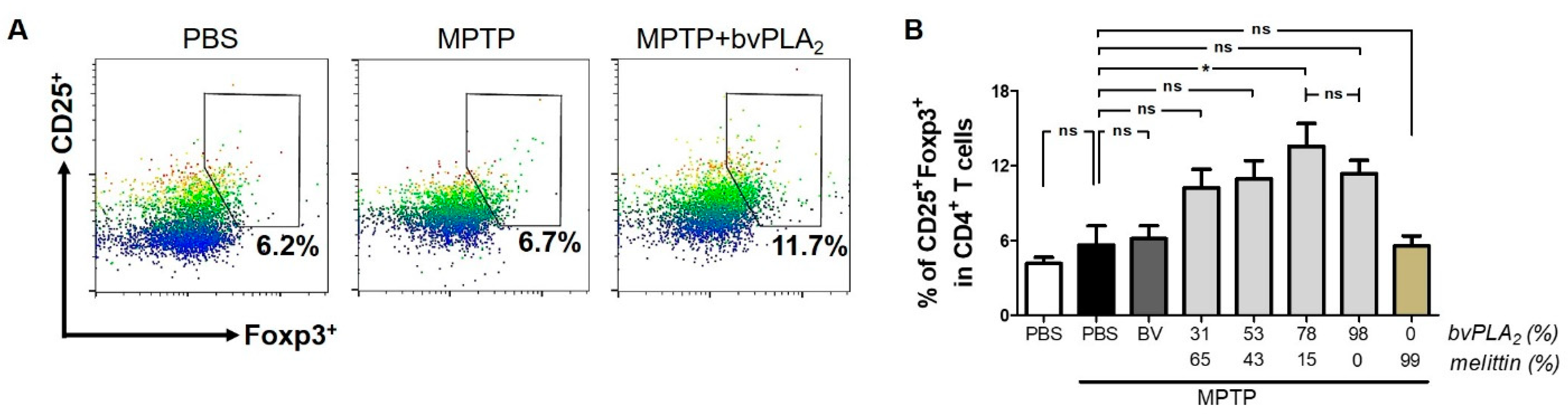
2.4. BvPLA2 with Enriched PLA2 Levels Induces Differentiation of Regulatory T Cells in MPTP-Challenged Mice
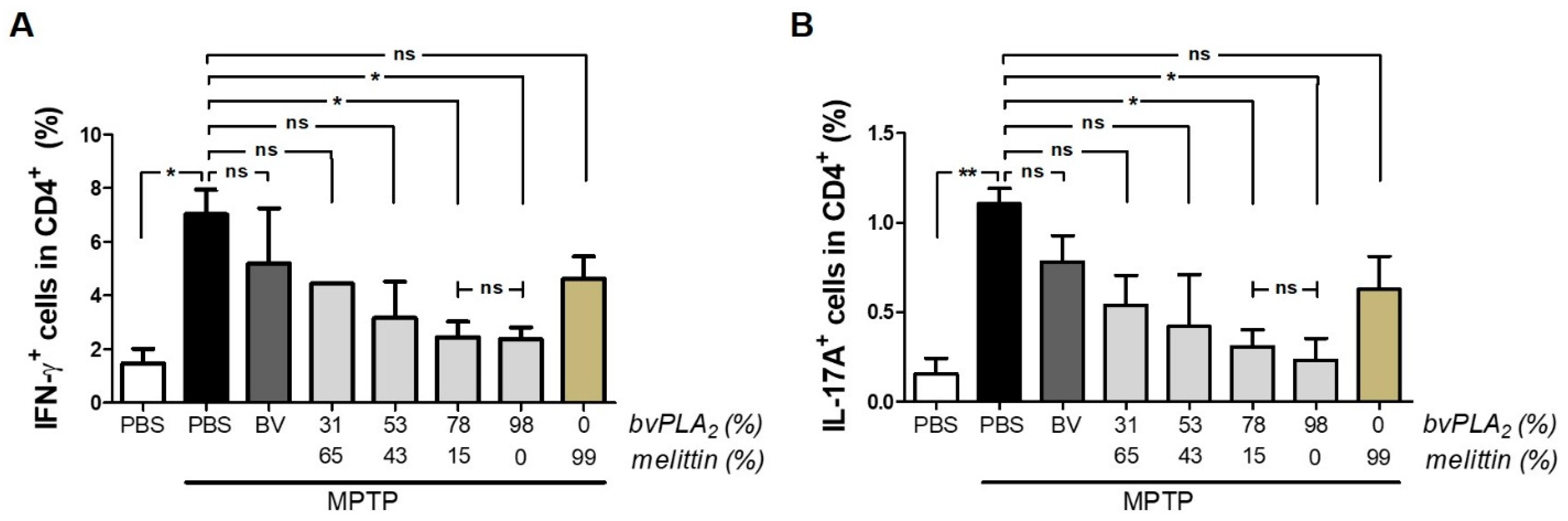
2.5. BvPLA2 with Enriched PLA2 Suppresses the Differentiation of CD4+ Effector T Cells in PD
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Preparation and Manufacturing of bvPLA2 and Melittin
5.2. Animals
5.3. MPTP-Induced Mouse Model
5.4. BvPLA2 Treatment
5.5. Immunohistochemistry
5.6. Unbiased Stereological Estimation
5.7. Flow Cytometry
5.8. Behavioral Test
5.9. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bertram, L.; Tanzi, R.E. The genetic epidemiology of neurodegenerative disease. J. Clin. Investig. 2005, 115, 1449–1457. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Fu, Q.; Cheng, L.; Zhai, M.; Wu, W.; Huang, L.; Du, G. Inflammatory response in Parkinson’s disease (review). Mol. Med. Rep. 2014, 10, 2223–2233. [Google Scholar] [CrossRef] [PubMed]
- Brochard, V.; Combadiere, B.; Prigent, A.; Laouar, Y.; Perrin, A.; Beray-Berthat, V.; Bonduelle, O.; Alvarez-Fischer, D.; Callebert, J.; Launay, J.M.; et al. Infiltration of CD4+ lymphocytes into the brain contributes to neurodegeneration in a mouse model of Parkinson disease. J. Clin. Investig. 2009, 119, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.A.; Cao, J.; Hsu, Y.H.; Magrioti, V.; Kokotos, G. Phospholipase A2 enzymes: Physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kang, S.S.; Kim, J.H.; Bae, C.S.; Choi, S.H. Inhibitory effect of whole bee venom in adjuvant-induced arthritis. In Vivo 2005, 19, 801–805. [Google Scholar] [PubMed]
- Silva, J.; Monge-Fuentes, V.; Gomes, F.; Lopes, K.; dos Anjos, L.; Campos, G.; Arenas, C.; Biolchi, A.; Goncalves, J.; Galante, P.; et al. Pharmacological alternatives for the treatment of neurodegenerative disorders: Wasp and bee venoms and their components as new neuroactive tools. Toxins 2015, 7, 3179–3209. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.S.; Lee, G.; Lee, C.; Ye, M.; Chung, H.S.; Kim, H.; Bae, S.J.; Hwang, D.S.; Bae, H. Bee venom phospholipase A2, a novel Foxp3+ regulatory T cell inducer, protects dopaminergic neurons by modulating neuroinflammatory responses in a mouse model of parkinson’s disease. J. Immunol. 2015, 195, 4853–4860. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Chung, H.S.; Lee, C.; Yoon, M.S.; Yu, A.R.; Kim, J.S.; Hwang, D.S.; Shim, I.; Bae, H. Neuroprotective effects of bee venom phospholipase A2 in the 3xTg AD mouse model of Alzheimer’s disease. J. Neuroinflamm. 2016, 13, 10. [Google Scholar] [CrossRef] [PubMed]
- Raghuraman, H.; Chattopadhyay, A. Melittin: A membrane-active peptide with diverse functions. Biosci. Rep. 2007, 27, 189–223. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Balling, R. The role of regulatory T cells in neurodegenerative diseases. Wiley interdisciplinary reviews. Syst. Biol. Med. 2013, 5, 153–180. [Google Scholar] [CrossRef]
- Chen, Y.; Qi, B.; Xu, W.; Ma, B.; Li, L.; Chen, Q.; Qian, W.; Liu, X.; Qu, H. Clinical correlation of peripheral CD4+cell subsets, their imbalance and Parkinson’s disease. Mol. Med. Rep. 2015, 12, 6105–6111. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.D.; Stone, D.K.; Hutter, J.A.; Benner, E.J.; Mosley, R.L.; Gendelman, H.E. Regulatory T cells attenuate Th17 cell-mediated nigrostriatal dopaminergic neurodegeneration in a model of Parkinson′s disease. J. Immunol. 2010, 184, 2261–2271. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Yamane, H.; Paul, W.E. Differentiation of effector CD4 T cell populations. Ann. Rev. Immunol. 2010, 28, 445–489. [Google Scholar] [CrossRef]
- Awad, K.; Abushouk, A.I.; AbdelKarim, A.H.; Mohammed, M.; Negida, A.; Shalash, A.S. Bee venom for the treatment of Parkinson′s disease: How far is it possible? Pharmacother. Biomed. Pharmacother. 2017, 91, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Habermann, E. Bee and wasp venoms. Science 1972, 177, 314–322. [Google Scholar] [CrossRef] [PubMed]
- De Souza, J.M.; Goncalves, B.D.C.; Gomez, M.V.; Vieira, L.B.; Ribeiro, F.M. Animal toxins as therapeutic tools to treat neurodegenerative diseases. Front. Pharmacol. 2018, 9, 145. [Google Scholar] [CrossRef] [PubMed]
- Stojkovska, I.; Wagner, B.M.; Morrison, B.E. Parkinson’s disease and enhanced inflammatory response. Exp. Biol. Med. 2015, 240, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Bae, H. Anti-inflammatory applications of melittin, a major component of bee venom: Detailed mechanism of action and adverse effects. Molecules 2016, 21, 616. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Guan, S.M.; Sun, W.; Fu, H. Melittin, the major pain-producing substance of bee venom. Neurosci. Bull. 2016, 32, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.Y.; Fujita, T.; Kumamoto, E. Phospholipase A2 activation by melittin enhances spontaneous glutamatergic excitatory transmission in rat substantia gelatinosa neurons. Neuroscience 2005, 135, 485–495. [Google Scholar] [CrossRef]
- Jackson-Lewis, V.; Przedborski, S. Protocol for the MPTP mouse model of Parkinson’s disease. Nat. Protoc. 2007, 2, 141–151. [Google Scholar] [CrossRef] [PubMed]
- West, M.J. New stereological methods for counting neurons. Neurobiol. Aging 1993, 14, 275–285. [Google Scholar] [CrossRef]
- Ogawa, N.; Hirose, Y.; Ohara, S.; Ono, T.; Watanabe, Y. A simple quantitative bradykinesia test in MPTP-treated mice. Res. Commun. Chem. Pathol. Pharmacol. 1985, 50, 435–441. [Google Scholar] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.H.; Kim, M.; Lee, J.; Jeon, H.N.; Kim, S.H.; Bae, H. Comparison of the Protective Effects of Bee Venom Extracts with Varying PLA2 Compositions in a Mouse Model of Parkinson’s Disease. Toxins 2019, 11, 358. https://doi.org/10.3390/toxins11060358
Kim KH, Kim M, Lee J, Jeon HN, Kim SH, Bae H. Comparison of the Protective Effects of Bee Venom Extracts with Varying PLA2 Compositions in a Mouse Model of Parkinson’s Disease. Toxins. 2019; 11(6):358. https://doi.org/10.3390/toxins11060358
Chicago/Turabian StyleKim, Kyung Hwa, Minhwan Kim, Jaehwan Lee, Hat Nim Jeon, Se Hyun Kim, and Hyunsu Bae. 2019. "Comparison of the Protective Effects of Bee Venom Extracts with Varying PLA2 Compositions in a Mouse Model of Parkinson’s Disease" Toxins 11, no. 6: 358. https://doi.org/10.3390/toxins11060358
APA StyleKim, K. H., Kim, M., Lee, J., Jeon, H. N., Kim, S. H., & Bae, H. (2019). Comparison of the Protective Effects of Bee Venom Extracts with Varying PLA2 Compositions in a Mouse Model of Parkinson’s Disease. Toxins, 11(6), 358. https://doi.org/10.3390/toxins11060358