Study on the Growth and Enterotoxin Production by Staphylococcus aureus in Canned Meat before Retorting

 , , and
, , and
Abstract
1. Introduction
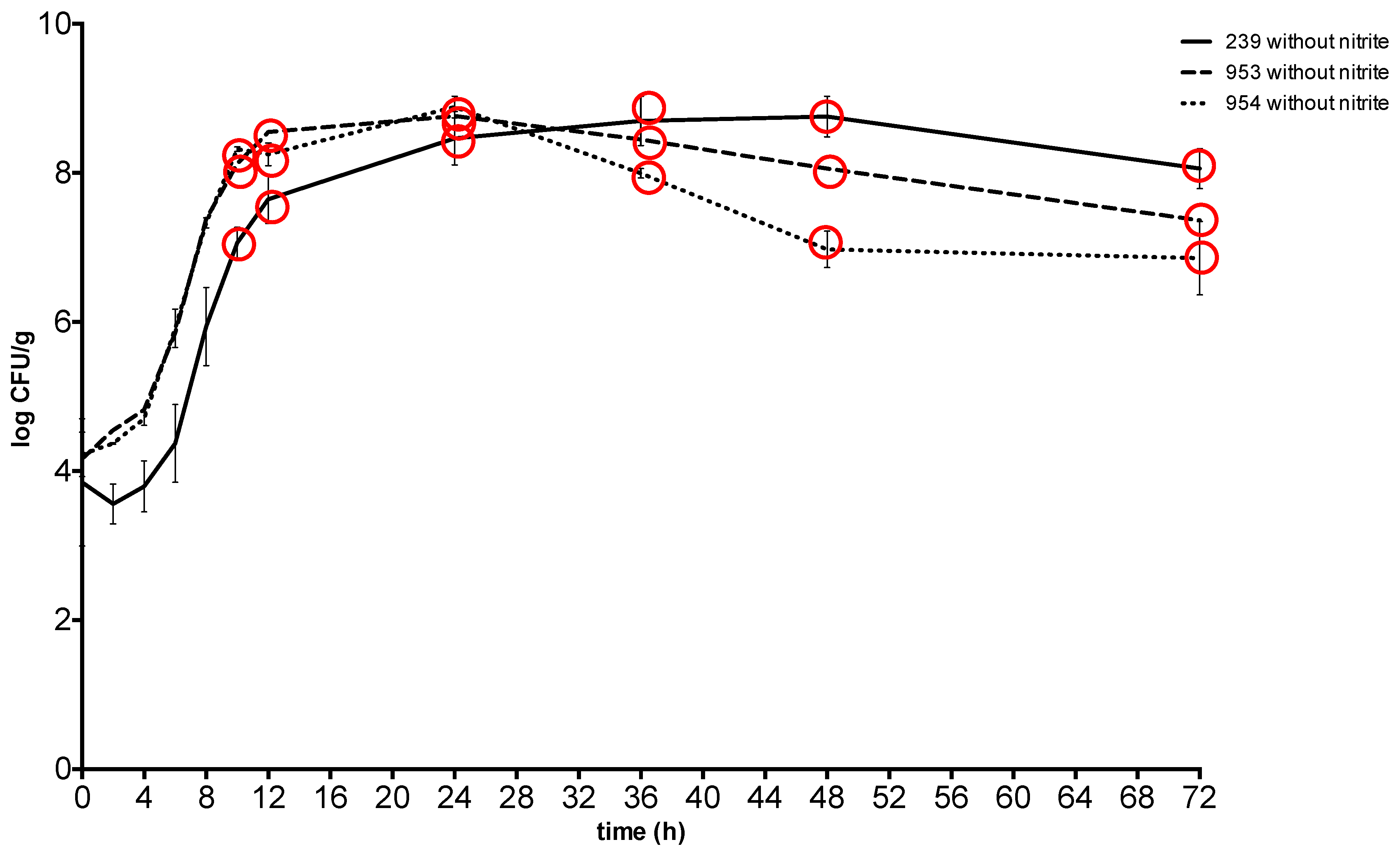
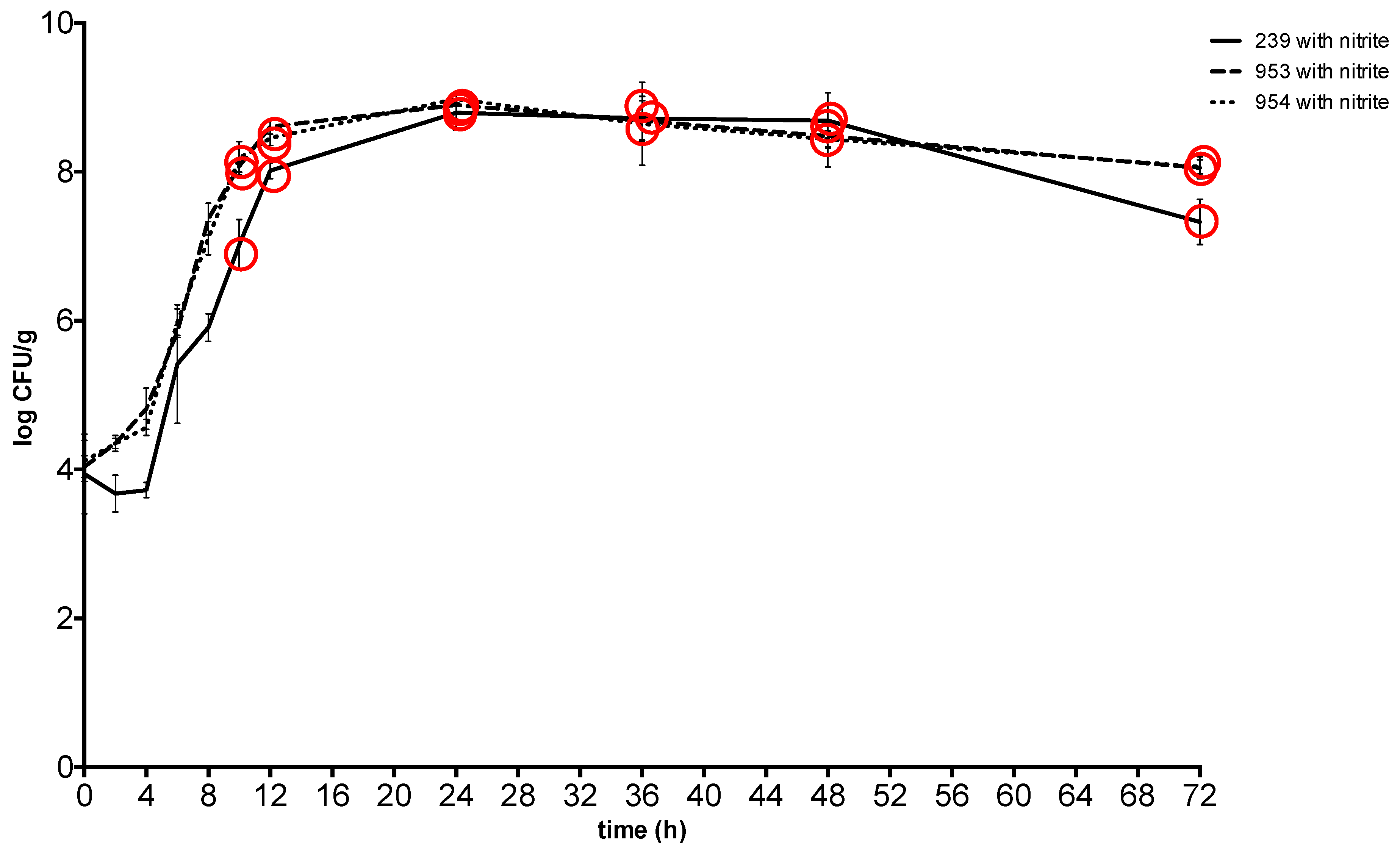
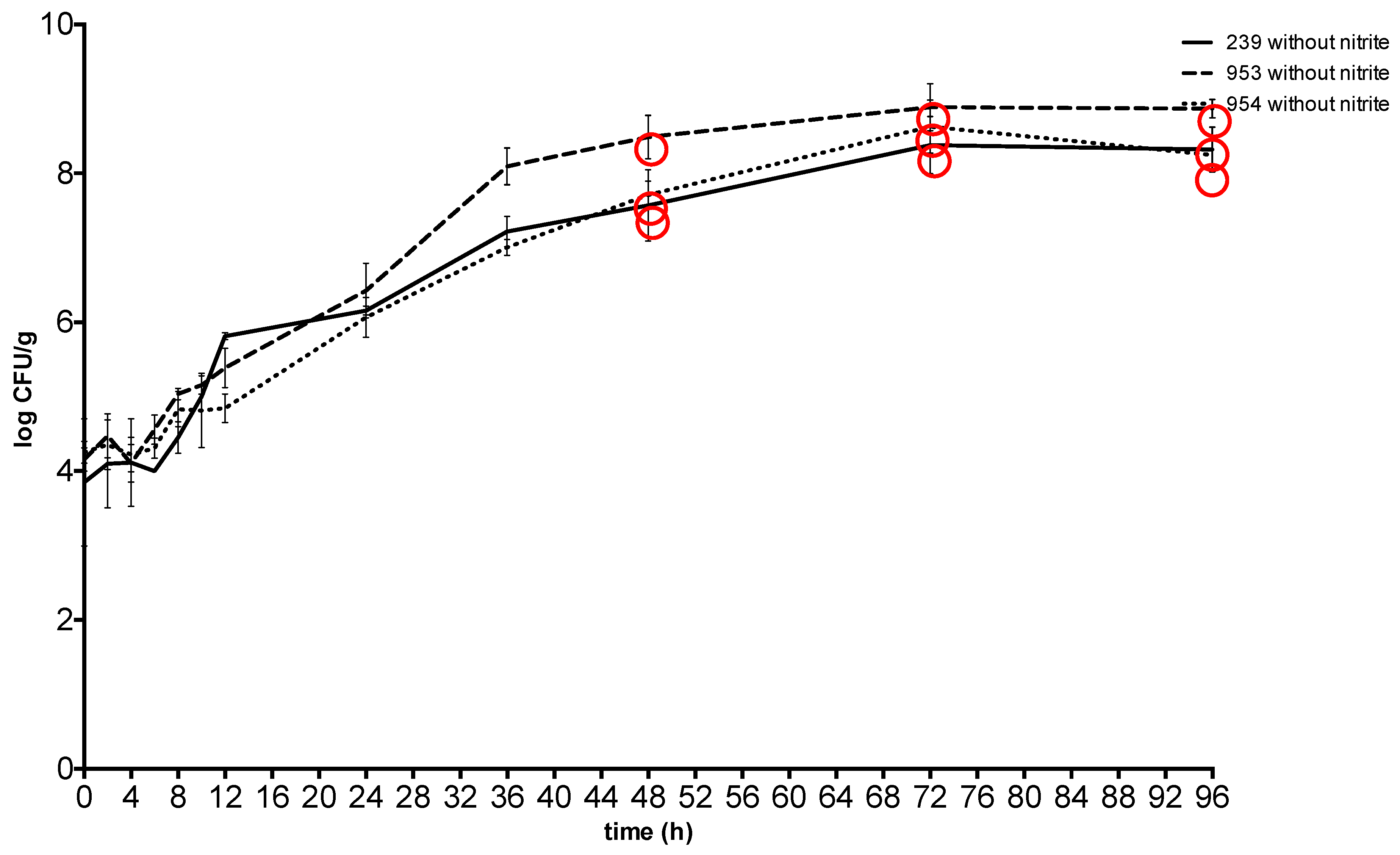
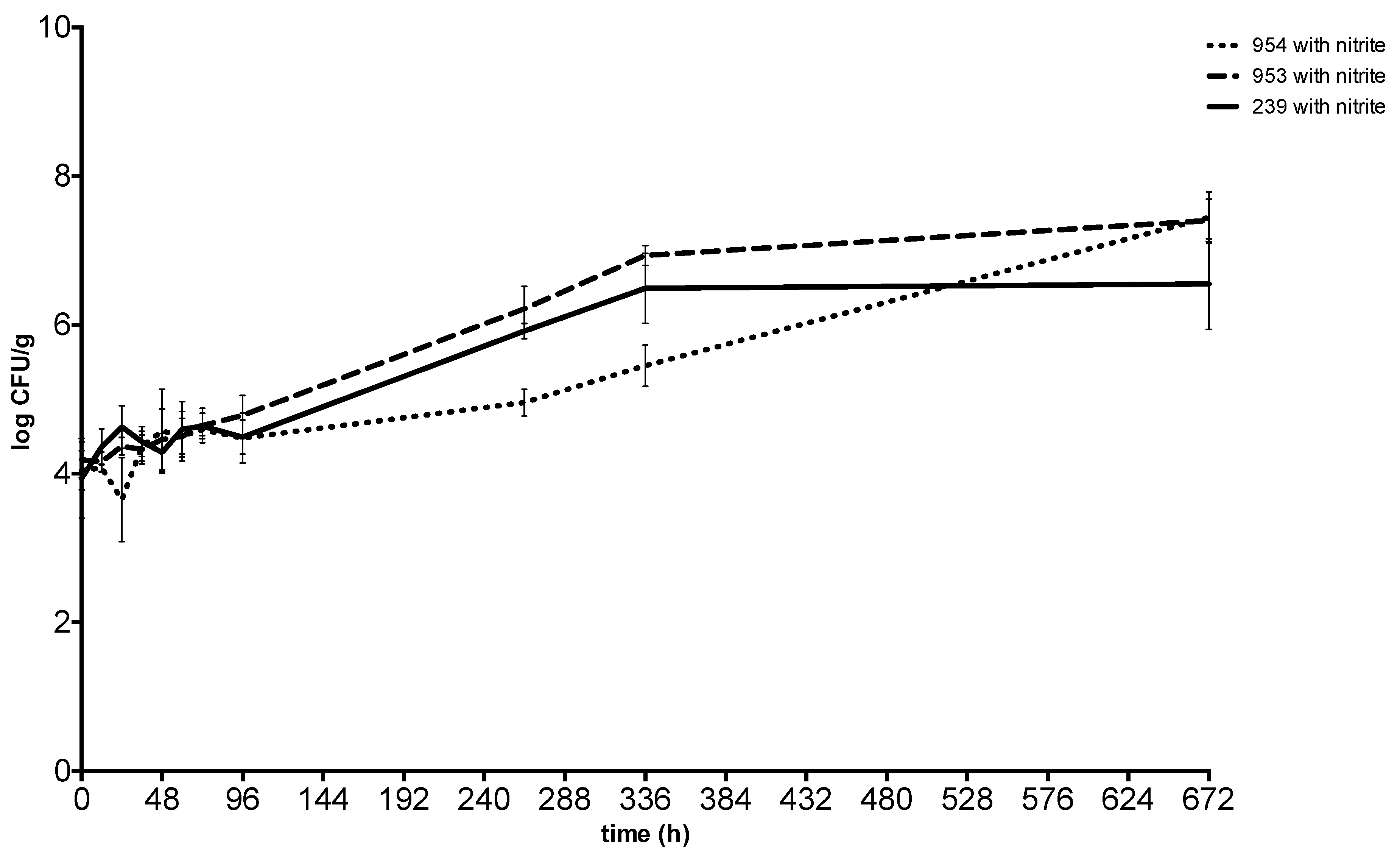
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Microbiological Analysis of Samples from the Factory
5.2. Canned Meat Spiking
5.3. Study of the Growth Rate
5.4. Enterotoxin Detection
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wallin-Carlquist, N.; Márta, D.; Borch, E.; Rådström, P. Prolonged expression and production of Staphylococcus aureus enterotoxin A in processed pork meat. Int. J. Food Microbiol. 2010, 141, S69–S74. [Google Scholar] [PubMed]
- Nasser, L.A. Molecular identification of isolated fungi, microbial and heavy metal contamination of canned meat products sold in Riyadh, Saudi Arabia. Saudi J. Biol. Sci. 2015, 22, 513–520. [Google Scholar] [CrossRef][Green Version]
- Blake, P.A.; Horwitz, M.A.; Hopkins, L.; Lombard, G.L.; McCroan, J.E.; Prucha, J.C.; Merson, M.H. Type A botulism from commercially canned beef stew. South. Med. J. 1977, 70, 5–7. [Google Scholar]
- Cragg, J.; Andrews, A.V. Observations on the microbiological flora of canned Parma ham. J. Hyg. 1973, 71, 417–422. [Google Scholar] [PubMed]
- Mitrica, L.; Granum, P.E. The amylase-producing microflora of semi-preserved canned sausages: Identification of the bacteria and characterization of their amylases. Z. Fur Lebensm.-Unters. Forsch. 1979, 169, 4–8. [Google Scholar] [CrossRef]
- Koreňová, J.; Rešková, Z.; Véghová, A.; Kuchta, T. Tracing Staphylococcus aureus in small and medium-sized food-processing factories on the basis of molecular sub-species typing. Int. J. Environ. Health Res. 2015, 25, 384–392. [Google Scholar] [CrossRef]
- Cenci-Goga, B.T.; Karama, M.; Rossitto, P.V.; Morgante, R.A.; Cullor, J.S. Enterotoxin Production by Staphylococcus aureus Isolated from Mastitic Cows. J. Food Prot. 2003, 66, 1693–1696. [Google Scholar] [CrossRef] [PubMed]
- Grispoldi, L.; Massetti, L.; Sechi, P.; Iulietto, M.F.; Ceccarelli, M.; Karama, M.; Popescu, P.A.; Pandolfi, F.; Cenci-Goga, B.T. Characterization of enterotoxin producing Staphylococcus aureus isolated from mastitic cows. J. Dairy Sci. 2019, 102, 1059–1065. [Google Scholar] [CrossRef] [PubMed]
- Hennekinne, J.A.; De Buyser, M.L.; Dragacci, S. Staphylococcus aureus and its food poisoning toxins: Characterization and outbreak investigation. FEMS Microbiol. Rev. 2012, 36, 815–836. [Google Scholar] [CrossRef]
- Asao, T.; Kumeda, Y.; Kawai, T.; Shibata, T.; Oda, H.; Haruki, K.; Nakazawa, H.; Kozaki, S. An extensive outbreak of staphylococcal food poisoning due to low-fat milk in Japan: Estimation of enterotoxin A in the incriminated milk and powdered skim milk. Epidemiol. Infect. 2003, 130, 33–40. [Google Scholar]
- Notermans, S.; Dormans, J.A.M.A.; Mead, G.C. Contribution of surface attachment to the establishment of micro-organisms in food processing plants: A review. Biofouling 1991, 5, 21–36. [Google Scholar] [CrossRef]
- Denayer, S.; Delbrassinne, L.; Nia, Y.; Botteldoorn, N. Food-Borne Outbreak Investigation and Molecular Typing: High Diversity of Staphylococcus aureus Strains and Importance of Toxin Detection. Toxins 2017, 9, 407. [Google Scholar] [CrossRef]
- Cha, J.O.; Lee, J.K.; Jung, Y.H.; Yoo, J.I.; Park, Y.K.; Kim, B.S.; Lee, Y.S. Molecular analysis of Staphylococcus aureus isolates associated with staphylococcal food poisoning in South Korea. J. Appl. Microbiol. 2006, 101, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Kerouanton, A.; Hennekinne, J.A.; Letertre, C.; Petit, L.; Chesneau, O.; Brisabois, A.; De Buyser, M.L. Characterization of Staphylococcus aureus strains associated with food poisoning outbreaks in France. Int. J. Food Microbiol. 2007, 115, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, R.J.; Kolvin, J.L.; Roberts, D. Canned foods - the problems of food poisoning and spoilage. Health Hyg. 1982, 4, 41–47. [Google Scholar]
- Mansfield, J.M.; Farkas, G.; Wieneke, A.A.; Gilbert, R.J. Studies on the growth and survival of Staphylococcus aureus in corned beef. J. Hyg. 1983, 91, 467–478. [Google Scholar] [CrossRef]
- Stersky, A.; Todd, E.; Pivnick, H. Food Poisoning Associated with Post-Process Leakage (PPL) in Canned Foods. J. Food Protect. 1980, 43, 465–476. [Google Scholar] [CrossRef]
- Beenken, K.E.; Dunman, P.M.; McAleese, F.; Macapagal, D.; Murphy, E.; Projan, S.J.; Blevins, J.S.; Smeltzer, M.S. Global Gene Expression in Staphylococcus aureus biofilms. J. Bacteriol. 2004, 186, 4665–4684. [Google Scholar] [CrossRef]
- Resch, A.; Rosenstein, R.; Nerz, C.; Götz, F. Differential Gene Expression Profiling of Staphylococcus aureus Cultivated under Biofilm and Planktonic Conditions. Appl. Environ. Microbiol. 2005, 71, 2663–2676. [Google Scholar] [CrossRef]
- Tsutsuura, S.; Shimamura, Y.; Murata, M. Temperature dependence of the production of staphylococcal enterotoxin a by Staphylococcus aureus. Biosci. Biotech. Biochem. 2013, 77, 30–37. [Google Scholar] [CrossRef]
- Valero, A.; Pérez-Rodríguez, F.; Carrasco, E.; Fuentes-Alventosa, J.M.; García-Gimeno, R.M.; Zurera, G. Modelling the growth boundaries of Staphylococcus aureus: Effect of temperature, pH and water activity. Int. J. Food Microbiol. 2009, 133, 186–194. [Google Scholar] [CrossRef]
- Betley, M.J.; Mekalanos, J.J. Staphylococcal enterotoxin A is encoded by phage. Science 1985, 229, 185–187. [Google Scholar] [CrossRef]
- Schelin, J.; Susilo, Y.B.; Johler, S. Expression of staphylococcal enterotoxins under stress encountered during food production and preservation. Toxins 2017, 9, 401. [Google Scholar] [CrossRef]
- Scott, W.J. Factors in canned ham controlling Cl. botulinum and Staph. aureus. Annales de l’Institut Pasteur de Lille 1955, 7, 68–74. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Points | Number of Samples | Mesophilic (Mean ± SD) | Staphylococcus spp. (Mean ± SD) | Micrococcus spp. (Mean ± SD) | Staphylococcus aureus (Number of Isolated Strains) |
|---|---|---|---|---|---|
| Frozen cooked beef at reception | 15 | 2.94 ± 1.57 | 1.92 ± 1.39 | 1.94 ± 1.76 | 6 |
| Sliced beef at the slicer | 12 | 2.06 ± 1.45 | 0.47 ± 0.85 | 1.24 ± 1.36 | 0 |
| Belt swabs | 9 | 1.51 ± 0.88 | 0.49 ± 0.78 | 0.34 ± 0.68 | 1 |
| Piston swabs | 9 | 1.52 ± 1.24 | 0.26 ± 0.51 | 0.19 ± 0.57 | 0 |
| Frozen cooked beef at thawing | 36 | 2.86 ± 0.83 | 1.49 ± 1.39 | 0.63 ± 1.12 | 4 |
| Defrost water | 6 | 2.02 ± 1.47 | 0.91 ± 0.74 | 0.00 ± 0.00 | 1 |
| Cooked beef after thawing | 6 | 3.12 ± 0.39 | 0.41 ± 1.01 | 0.33 ± 0.82 | 0 |
| Meat on the belt after metal detector | 12 | 3.91 ± 1.79 | 2.52 ± 1.62 | 2.84 ± 1.72 | 1 |
| Aspic at the doser | 3 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grispoldi, L.; Popescu, P.A.; Karama, M.; Gullo, V.; Poerio, G.; Borgogni, E.; Torlai, P.; Chianese, G.; Fermani, A.G.; Sechi, P.; et al. Study on the Growth and Enterotoxin Production by Staphylococcus aureus in Canned Meat before Retorting. Toxins 2019, 11, 291. https://doi.org/10.3390/toxins11050291
Grispoldi L, Popescu PA, Karama M, Gullo V, Poerio G, Borgogni E, Torlai P, Chianese G, Fermani AG, Sechi P, et al. Study on the Growth and Enterotoxin Production by Staphylococcus aureus in Canned Meat before Retorting. Toxins. 2019; 11(5):291. https://doi.org/10.3390/toxins11050291
Chicago/Turabian StyleGrispoldi, Luca, Paul Alexanderu Popescu, Musafiri Karama, Vito Gullo, Giusi Poerio, Elena Borgogni, Paolo Torlai, Giuseppina Chianese, Anna Giovanna Fermani, Paola Sechi, and et al. 2019. "Study on the Growth and Enterotoxin Production by Staphylococcus aureus in Canned Meat before Retorting" Toxins 11, no. 5: 291. https://doi.org/10.3390/toxins11050291
APA StyleGrispoldi, L., Popescu, P. A., Karama, M., Gullo, V., Poerio, G., Borgogni, E., Torlai, P., Chianese, G., Fermani, A. G., Sechi, P., & Cenci-Goga, B. (2019). Study on the Growth and Enterotoxin Production by Staphylococcus aureus in Canned Meat before Retorting. Toxins, 11(5), 291. https://doi.org/10.3390/toxins11050291