Impact of Five Succinate Dehydrogenase Inhibitors on DON Biosynthesis of Fusarium asiaticum, Causing Fusarium Head Blight in Wheat

 and
and
Abstract
1. Introduction
2. Results
2.1. Sensitivity of FGSC to Five Succinate Dehydrogenase Inhibitors
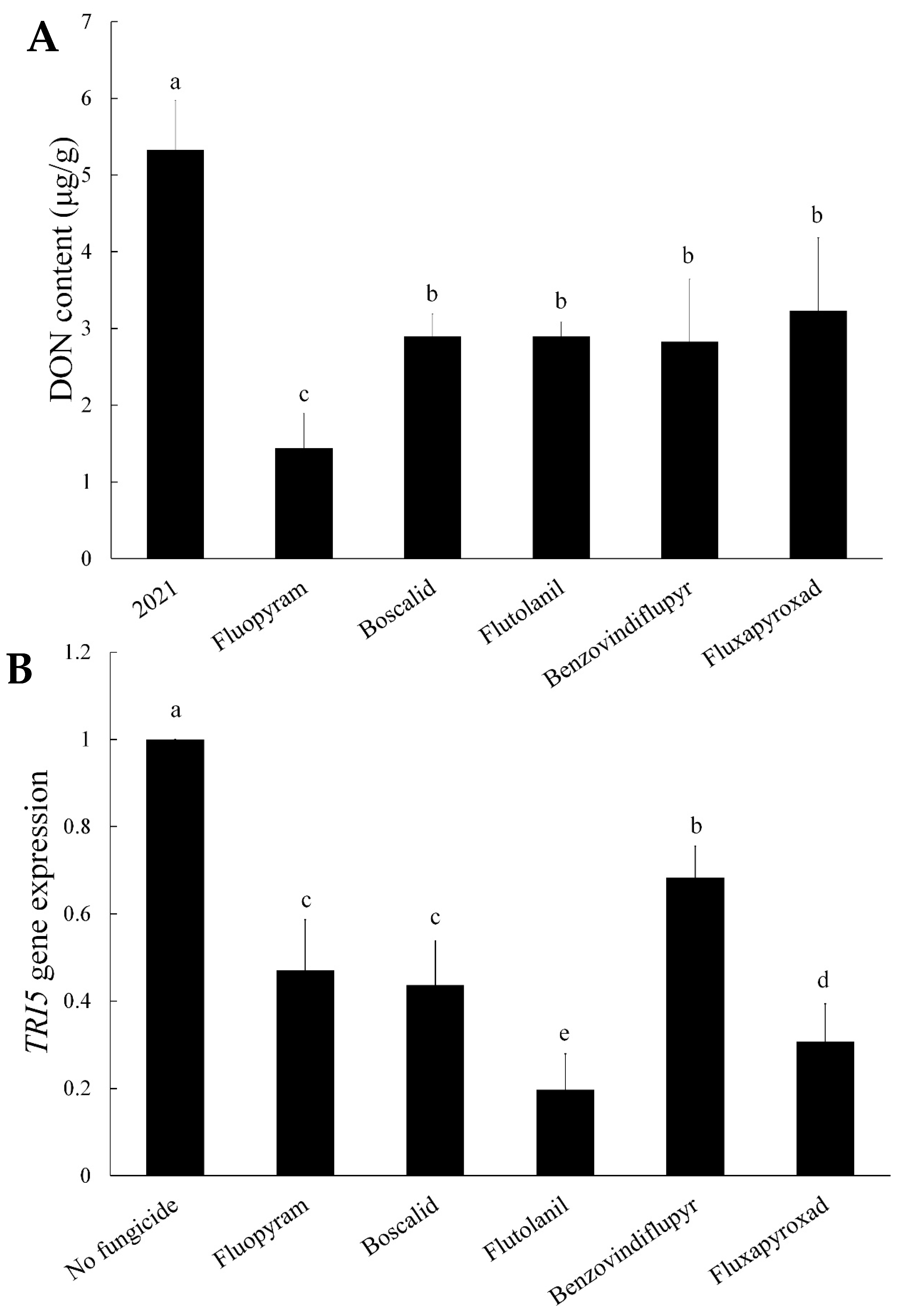
2.2. DON Production and TRI5 Gene Expression
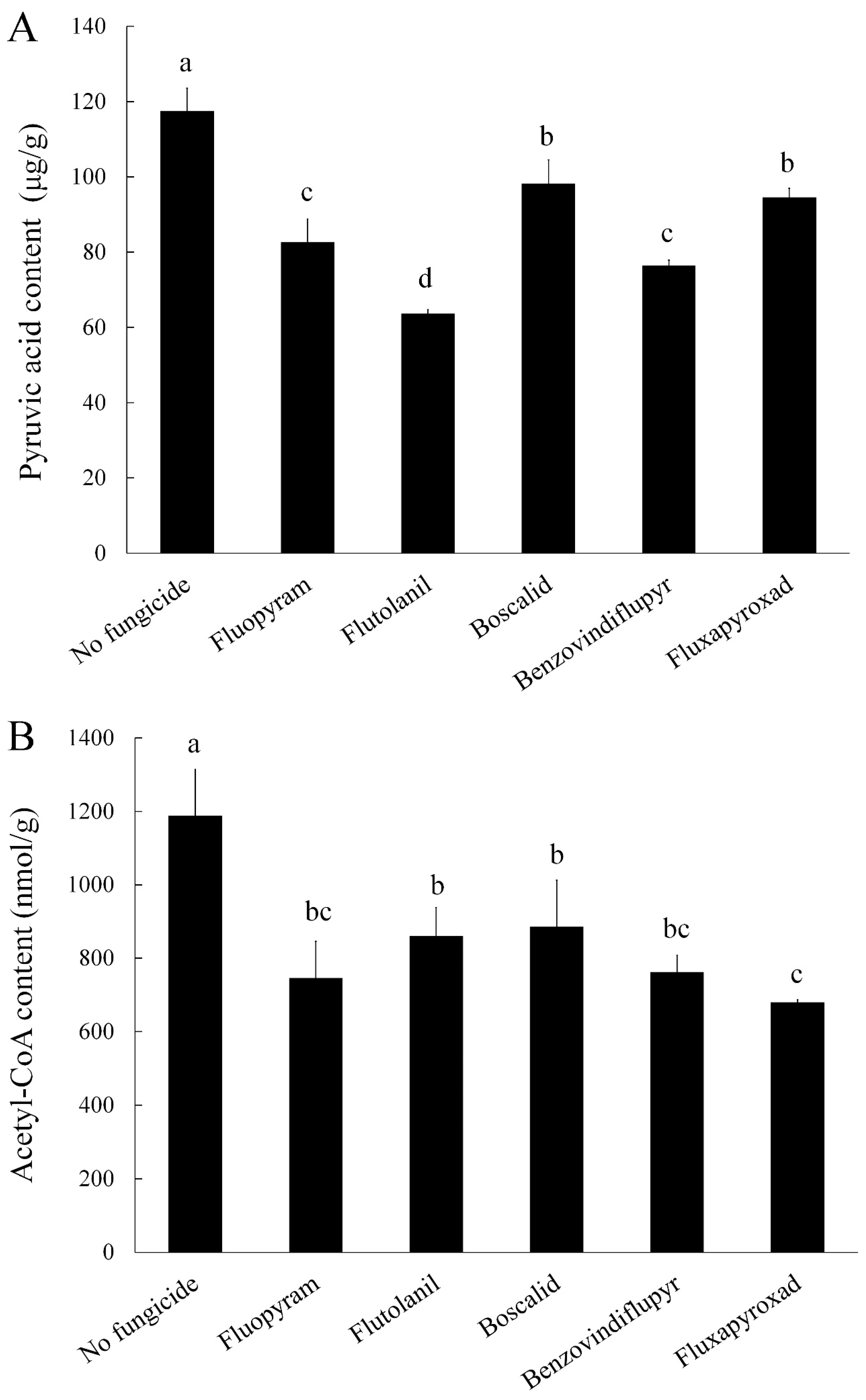
2.3. Pyruvic Acid and Acetyl-CoA Content
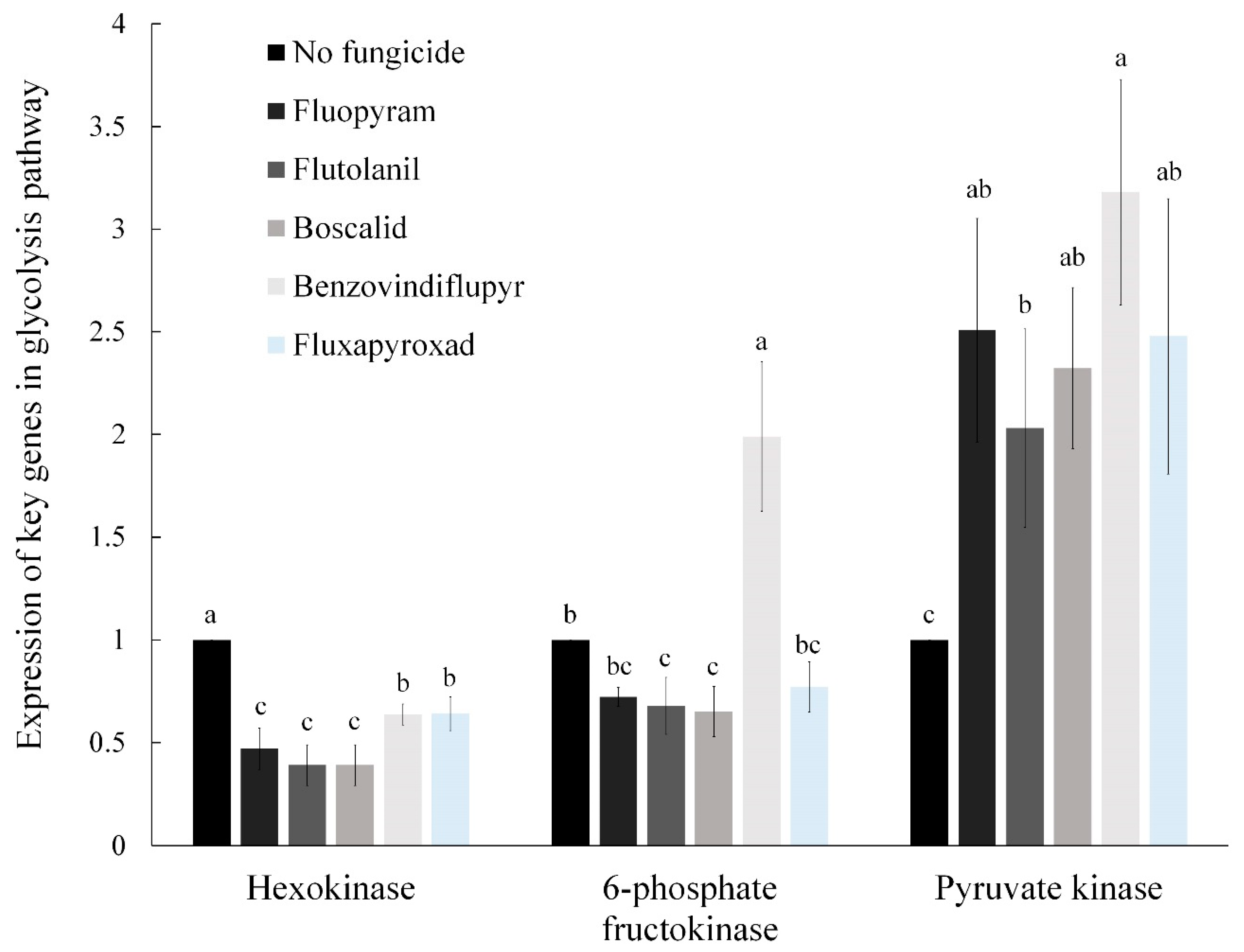
2.4. The Relative Expression of Key Genes in the Glycolysis Pathway
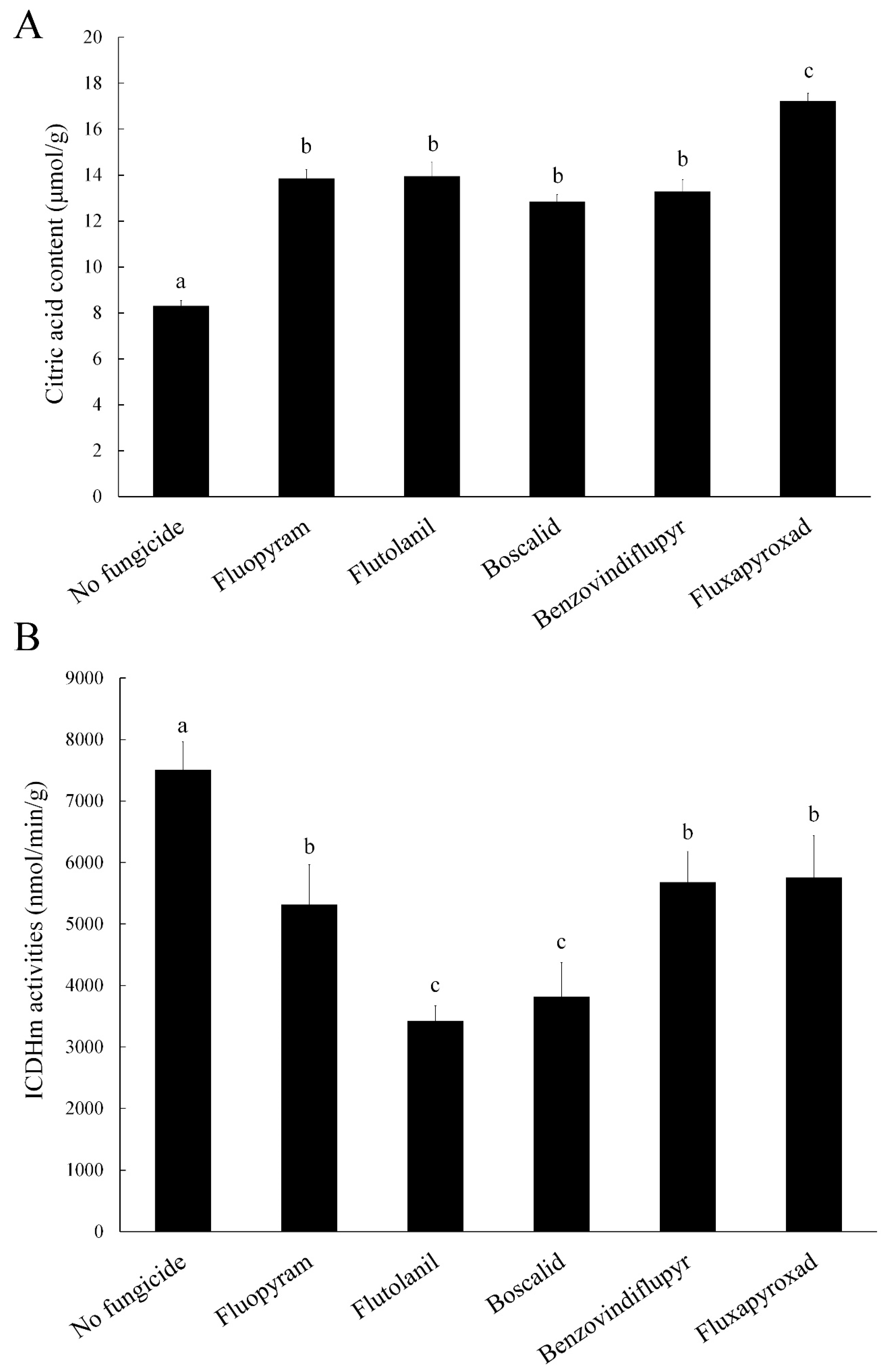
2.5. Citric Acid Content
2.6. Isocitrate Dehydrogenase Mitochondrial (ICDHm) Activity
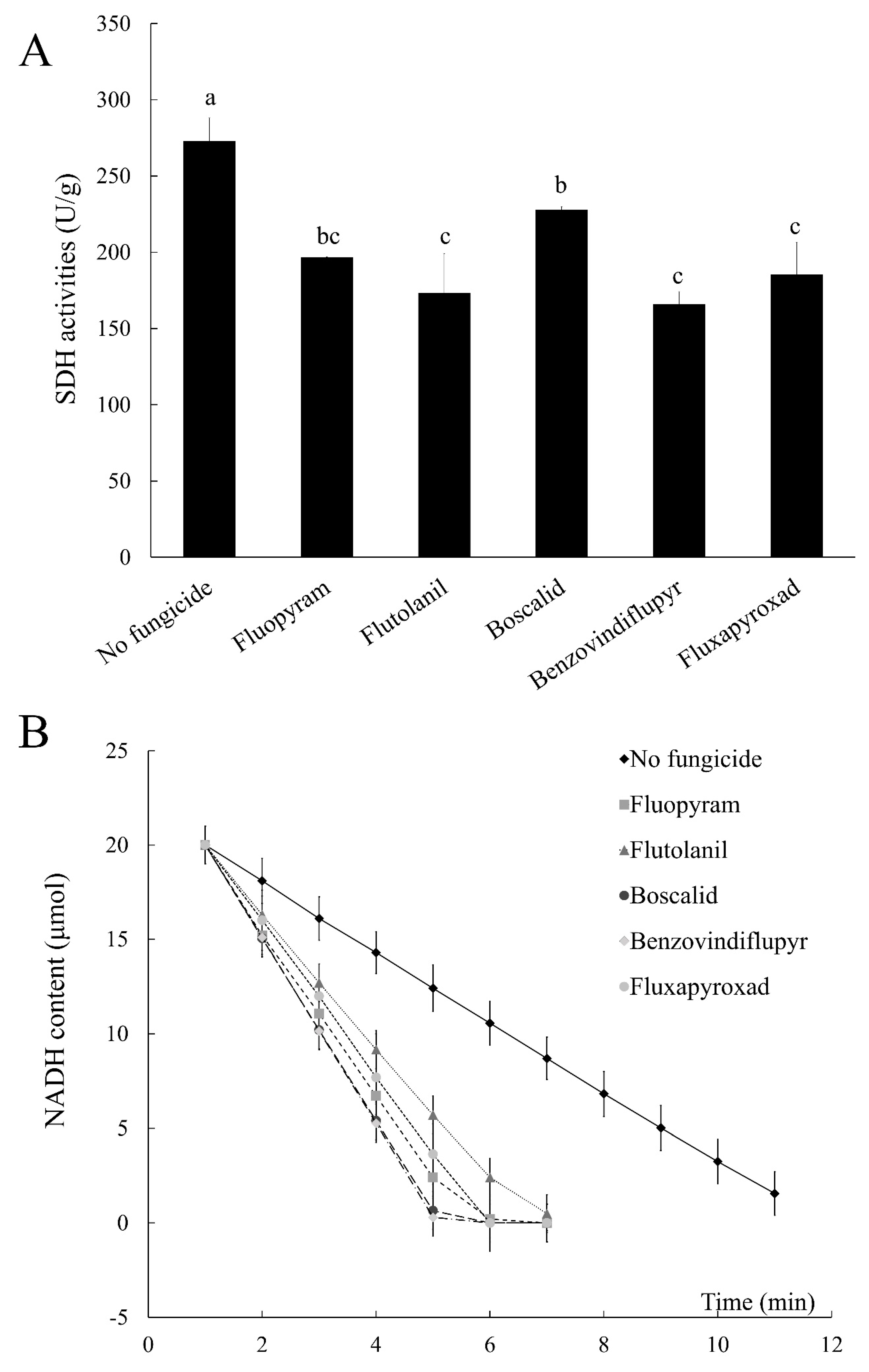
2.7. Succinate Dehydrogenase (SDH) and NADH Dehydrogenase Activities
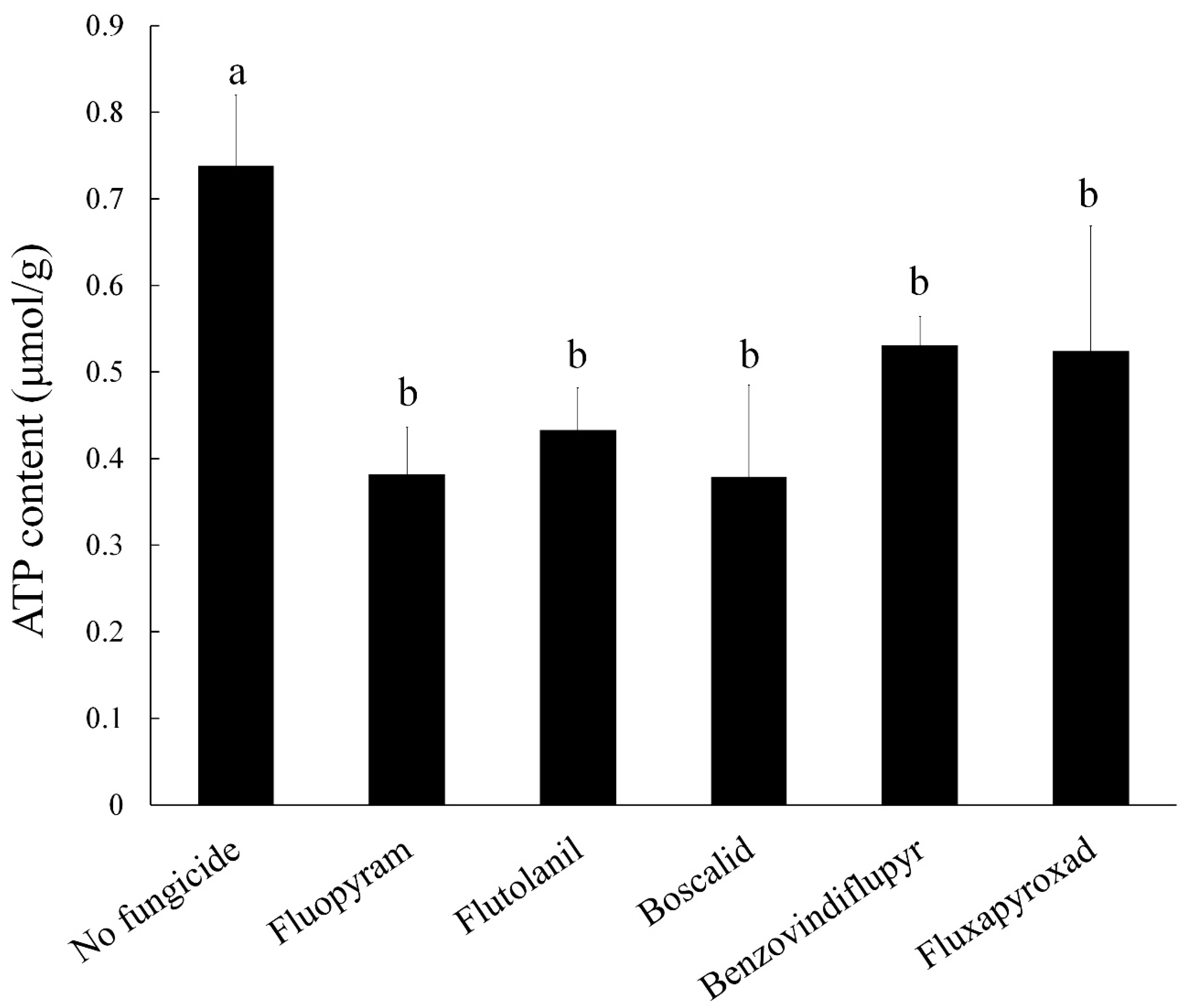
2.8. ATP Content
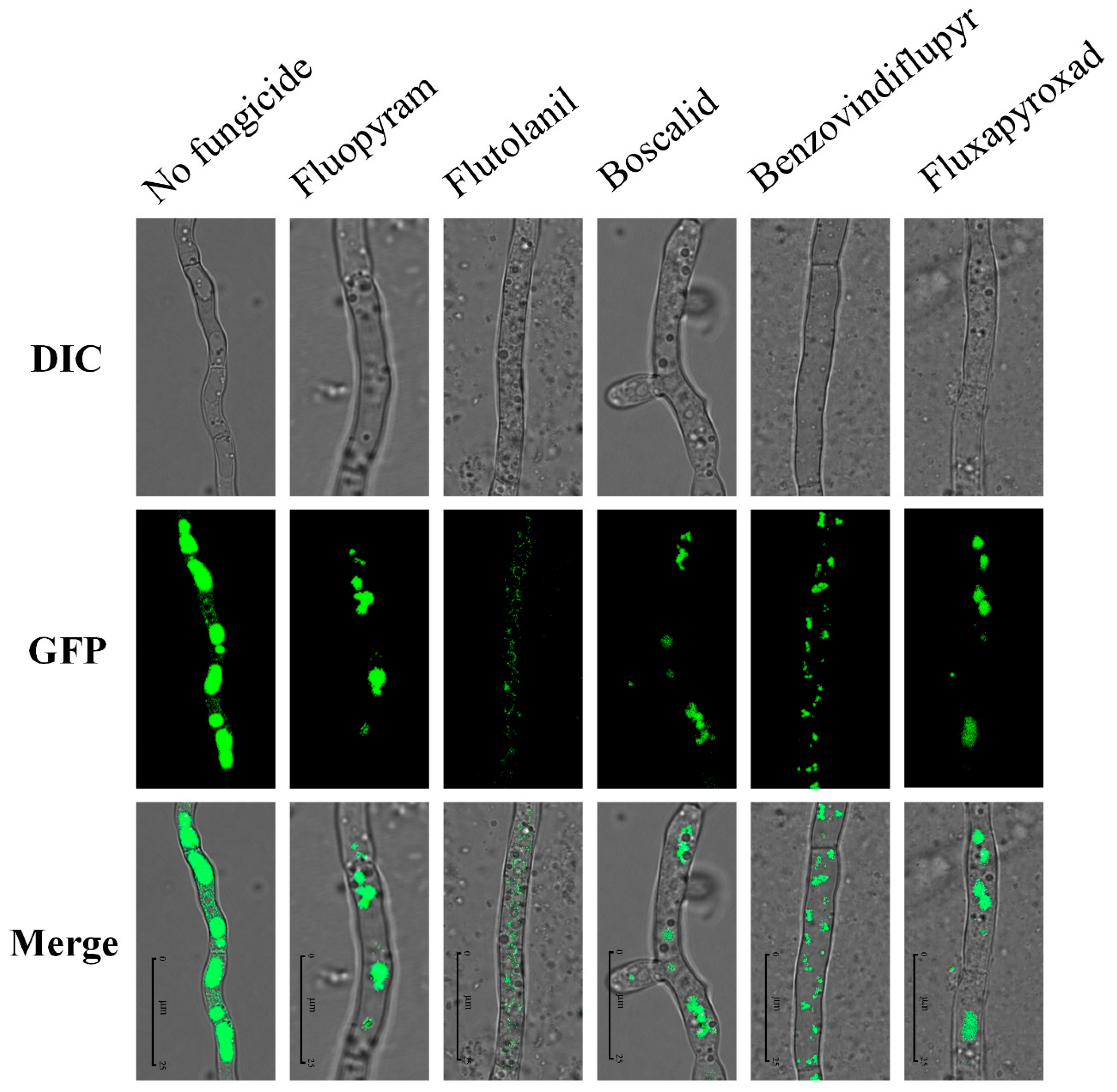
2.9. Impact on Toxisomes
3. Discussion
4. Materials and Methods
4.1. Fungicides, Fungal Strains and Culture Conditions
4.2. Fungicide Sensitivity Tests Based on Mycelial Growth and Spore Germination
4.3. RNA Extraction and Reverse Transcription PCR
4.4. DON Production and TRI5 Gene Expression
4.5. Expression of Key Genes in Glycolysis Pathway
4.6. Determination of Pyruvic acid, Acetyl-CoA, ATP and Citric Acid
4.7. SDH, ICDHm and NADH Dehydrogenase Activities
4.8. Microscopic Examinations
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Donnell, K.; Ward, T.J.; Aberra, D.; Kistler, H.C.; Aoki, T.; Orwig, N.; Kimura, M.; Bjornstad, S.; Klemsdal, S.S. Multilocus genotyping and molecular phylogenetics resolve a novel head blight pathogen within the Fusarium graminearum species complex from Ethiopia. Fungal Genet. Biol. 2008, 45, 1514–1522. [Google Scholar] [CrossRef] [PubMed]
- Yli-Mattila, T.; Gagkaeva, T.; Ward, T.J.; Aoki, T.; Kistler, H.C.; O’Donnell, K. A novel Asian clade within the Fusarium graminearum species complex includes a newly discovered cereal head blight pathogen from the Russian Far East. Mycologia 2009, 101, 841–852. [Google Scholar] [CrossRef]
- Zhang, H.; Theo, V.D.L.; Cees, W.; Chen, W.; Xu, J.; Xu, J.; Zhang, Y.; Feng, J. Population analysis of the Fusarium graminearum species complex from wheat in China show a shift to more aggressive isolates. PLoS ONE 2012, 7, e31722. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jia, X.; Chen, C.; Zhou, M. Characterization of carbendazim sensitivity and trichothecene chemotypes of Fusarium graminearum in Jiangsu province of China. Physiol. Mol. Plant Pathol. 2013, 84, 53–60. [Google Scholar] [CrossRef]
- Tanaka, T.; Hasegawa, A.; Yamamoto, S.; Lee, U.S.; Sugiura, Y.; Ueno, Y. Worldwide contamination of cereals by the Fusarium mycotoxins nivalenol, deoxynivalenol, and zearalenone. 1. Survey of 19 countries. J. Agric. Food Chem. 1988, 36, 979–983. [Google Scholar] [CrossRef]
- Goswami, R.S.; Kistler, H.C. Pathogenicity and in planta mycotoxin accumulation among members of the Fusarium graminearum species complex on wheat and rice. Phytopathology 2005, 95, 1397–1404. [Google Scholar] [CrossRef]
- Li, J.; Duan, Y.; Bian, C.; Pan, X.; Yao, C.; Wang, J.; Zhou, M. Effects of validamycin in controlling Fusarium head blight caused by Fusarium graminearum: Inhibition of DON biosynthesis and induction of host resistance. Pestic. Biochem. Physiol. 2019, 153, 152–160. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Zhou, M.; Chen, C.; Yuan, S.K. Vegetative compatibility of Fusarium graminearum isolates and genetic study on their carbendazim-resistance recombination in China. Phytopathology 2007, 97, 1584. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, X.; Ge, C.; Wang, Y.; Cao, J.; Jia, X.; Wang, J.; Zhou, M. Development and application of loop-mediated isothermal amplification for detection of the F167Y mutation of carbendazim-resistant isolates in Fusarium graminearum. Sci. Rep. 2014, 4, 7094. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, W.; Zhang, A.; Gu, C.; Zhou, M. Activity of the fungicide JS399-19 against Fusarium head blight of wheat and the risk of resistance. J. Integr. Agric. 2011, 10, 1906–1913. [Google Scholar] [CrossRef]
- Hgerhall, C. Succinate: Quinone oxidoreductases. Variations on a conserved theme. Biochim. Biophys. Acta-Bioenerget. 1997, 1320, 107–141. [Google Scholar] [CrossRef]
- Kuhn, P.J. Mode of action of carboximides. Symp. Ser. Br. Mycol. Soc. 1984, 9, 155–183. [Google Scholar]
- Keon, J.P.; White, G.A.; Hargreaves, J.A. Isolation, characterization and sequence of a gene conferring resistance to the systemic fungicide carboxin from the maize smut pathogen, Ustilago maydis. Curr. Genet. 1991, 19, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Ackrell, B.A. Progress in understanding structure–function relationships in respiratory chain complex II. FEBS Lett. 2000, 466, 1–5. [Google Scholar] [CrossRef]
- Bardas, G.A.; Veloukas, T.; Koutita, O.; Karaoglanidis, G.S. Multiple resistance of Botrytis cinerea from kiwifruit to SDHIs, Qols and fungicides of other chemical groups. Pest Manag. Sci. 2010, 66, 967–973. [Google Scholar] [CrossRef]
- Hervé, F.A.; Themis, J.M. Progress in understanding molecular mechanisms and evolution of resistance to succinate dehydrogenase inhibiting (SDHI) fungicides in phytopathogenic fungi. Crop Prot. 2010, 29, 643–651. [Google Scholar]
- Zhou, M.; Ye, Z.; Liu, J. Progress of fungicide resistance. Nanjing Agric. Univ. 1994, 17, 33–41. [Google Scholar]
- Duan, Y.; Xiu, Q.; Li, H.; Li, T.; Wang, J.; Zhou, M. Pharmacological characteristics and control efficacy of a novel SDHI fungicide pydiflumetofen against Sclerotinia sclerotiorum. Plant Dis. 2019, 103, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Alexander, N.J.; Proctor, R.H.; Mccormick, S.P. Genes, gene clusters, and biosynthesis of trichothecenes and fumonisins in fusarium. Journal of Toxicology. Toxin Rev. 2009, 28, 198–215. [Google Scholar] [CrossRef]
- Zhang, L.; Li, B.; Zhang, Y.; Jia, X.; Zhou, M. Hexokinase plays a critical role in deoxynivalenol (DON) production and fungal development in Fusarium graminearum. Mol. Plant Pathol. 2015, 17, 16–28. [Google Scholar] [CrossRef]
- Kimura, M.; Tokai, T.; Takahashi-Ando, N.; Ohsato, S.; Fujimura, M. Molecular and genetic studies of Fusarium trichothecene biosynthesis: Pathways, genes, and evolution. Biosci. Biotechnol. Biochem. 2007, 71, 2105–2123. [Google Scholar] [CrossRef] [PubMed]
- Barnett, J.A.; Kornberg, H.L. The utilization by yeasts of acids of the tricarboxylic acid cycle. J. Gen. Microbiol. 1960, 23, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Halabe, B.A. The biological significance of cancer: Umitochondria as a cause of cancer and the inhibition of glycolytic with citrate as a cancer treatment. Med. Hypotheses 2007, 69, 826–828. [Google Scholar] [CrossRef] [PubMed]
- Laporte, D.C.; Stueland, C.S.; Ikeda, T.P. Isocitrate dehydrogenase kinase/phosphatase. Biochimie 1989, 71, 1051–1057. [Google Scholar] [CrossRef]
- Garnak, M.; Reeves, H.C. Phosphorylation of isocitrate dehydrogenase of Escherichia coli. Science 1979, 203, 1111–1112. [Google Scholar] [CrossRef]
- Boenisch, M.J.; Broz, K.L.; Purvine, S.O.; Chrisler, W.B.; Nicora, C.D.; Connolly, L.R.; Freitag, M.; Baker, S.E.; Kistler, H.C. Structural reorganization of the fungal endoplasmic reticulum upon induction of mycotoxin biosynthesis. Sci. Rep. 2017, 7, 44296. [Google Scholar] [CrossRef] [PubMed]
- Mcmullen, M.; Jones, R.; Gallenberg, D. Scab of wheat and barley: A re-emerging disease of devastating impact. Plant Dis. 1997, 81, 1340–1348. [Google Scholar] [CrossRef]
- Parry, D.W.; Jenkinson, P.; Mcleod, L. Fusarium ear blight (scab) in small grain creals—A review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, J.; Zhang, Y.; Zhang, X.; Cheng, C.; Wang, J.; Hollomon, D.W.; Fan, P.S.; Zhou, M. Effect of carbendazim resistance on trichothecene production and aggressiveness of Fusarium graminearum. Mol. Plant Microbe Interact. 2009, 22, 1143–1150. [Google Scholar] [CrossRef]
- Liu, S.; Duan, Y.; Ge, C.; Chen, C.; Zhou, M. Functional analysis of the β2-tubulin gene of Fusarium graminearum and the β-tubulin gene of Botrytis cinerea by homologous replacement. Pest Manag. Sci. 2013, 69, 582–588. [Google Scholar] [CrossRef]
- Yuan, S.; Zhou, M. A major gene for resistance to carbendazim, in field isolates of Gibberella zeae. Can. J. Plant Pathol. 2005, 27, 58–63. [Google Scholar] [CrossRef]
- Duan, Y.; Xiao, X.; Li, T.; Chen, W.; Wang, J.; Fraaije, B.A.; Zhou, M. Impact of epoxiconazole on Fusarium head blight control, grain yield and deoxynivalenol accumulation in wheat. Pestic. Biochem. Phys. 2018, 152, 138–147. [Google Scholar] [CrossRef]
- Duan, Y.; Yang, Y.; Li, T.; Zhao, D.; Cao, J.; Shi, Y.; Wang, J.; Zhou, M. Development of a rapid and high-throughput molecular method for detecting the F200Y mutant genotype in benzimidazole-resistant isolates of Fusarium asiaticum. Pest Manag. Sci. 2016, 72, 2128–2135. [Google Scholar] [CrossRef]
- Liu, S.; Che, Z.; Chen, G. Multiple-fungicide resistance to carbendazim, diethofencarb, procymidone, and pyrimethanil in field isolates of Botrytis cinerea from tomato in Henan Province, China. Crop Prot. 2016, 84, 56–61. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, Z.; Chen, C.; Zhou, M.; Wang, H. Effects of fungicides JS399-19, azoxystrobin, tebuconazloe, and carbendazim on the physiological and biochemical indices and grain yield of winter wheat. Pestic. Biochem. Phys. 2010, 98, 151–157. [Google Scholar] [CrossRef]
- Chen, C.; Wang, J.; Luo, Q.; Yuan, S.; Zhou, M. Characterization and fitness of carbendazim-resistant strains of Fusarium graminearum (wheat scab). Pest Manag. Sci. 2007, 63, 1201–1207. [Google Scholar] [CrossRef]
- Bai, G.; Plattner, R.; Desjardins, A.; Kolb, F.; Mcintosh, R.A. Resistance to Fusarium head blight and deoxynivalenol accumulation in wheat. Plant Breed. 2001, 120, 1–6. [Google Scholar] [CrossRef]
- Haidukowski, M.; Pascale, M.; Perrone, G.; Pancaldi, D.; Campagna, C.; Visconti, A. Effect of fungicides on the development of Fusarium head blight, yield and deoxynivalenol accumulation in wheat inoculated under field conditions with Fusarium graminearum and Fusarium culmorum. J. Sci. Food Agric. 2005, 85, 191–198. [Google Scholar] [CrossRef]
- Duan, Y.; Tao, X.; Zhao, H.; Xiao, X.; Li, M.; Wang, J.; Zhou, M. Activity of demethylation inhibitor fungicide metconazole on Chinese Fusarium graminearum species complex and its application in carbendazim-resistance management of Fusarium head blight in wheat. Plant Dis. 2019. [Google Scholar] [CrossRef]
- Masiello, M.; Somma, S.; Ghionna, V.; Logrieco, A.F.; Moretti, A. In vitro and in field response of different fungicides against Aspergillus flavus and Fusarium species causing ear rot disease of maize. Toxins 2019, 11, 11. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Hohn, T.M.; McCormick, S.P. Trichothecene biosynthesis in Fusarium species-chemistry, genetics, and significance. Microbiol. Rev. 1993, 57, 595–604. [Google Scholar]
- Kimura, M.; Anzai, H.; Yamaguchi, I. Microbial toxins in plant–pathogen interactions: Biosynthesis, resistance mechanisms, and significance. J. Gen. Appl. Microbiol. 2001, 47, 149–160. [Google Scholar] [CrossRef]
- Cody, G.D.; Boctor, N.Z.; Filley, T.R.; Hazen, R.M.; Scott, J.H.; Sharma, A.; Yoder, H.S. Primordial carbonylated iron–sulfur compounds and the synthesis of pyruvate. Science 2000, 289, 1337–1340. [Google Scholar] [CrossRef]
- Suga, H.; Karugia, G.W.; Ward, T.; Gale, L.R.; Tomimura, K.; Nakajima, T.; Miyasaka, A.; Koizumi, S.; Kageyama, K.; Hyakumachi, M. Molecular characterization of the Fusarium graminearum species complex in Japan. Phytopathology 2008, 98, 159–166. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, N.; Yin, Y.; Chen, Y.; Jiang, J.; Ma, Z. Histone H3K4 methylation regulates hyphal growth, secondary metabolism and multiple stress responses in Fusarium graminearum. Environ. Microbiol. 2015, 17, 4615–4630. [Google Scholar] [CrossRef]
- Cappelli, R.A.; Peterson, J.L. Macroconidium formation in submerged cultures by a non-sporulating strain of Gibberella Zeae. Mycologia 1965, 57, 962. [Google Scholar]
- Bai, G.; Shaner, G. Variation in Fusarium graminearum and cultivar resistance to wheat scab. Plant Dis. 1996, 80, 975. [Google Scholar] [CrossRef]
- Qiu, J.; Xu, J.; Yu, J.; Bi, C.; Chen, C.; Zhou, M. Localisation of the benzimidazole fungicide binding site of Gibberella zeae β2-tubulin studied by site-directed mutagenesis. Pest Manag. Sci. 2011, 67, 191–198. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhu, Y.; Li, Y.; Duan, Y.; Zhang, R.; Zhou, M. β-1 tubulin rather than b2 tubulin is the preferred binding target for carbendazim in Fusarium graminearum. Phytopathology 2016, 106, 978–985. [Google Scholar] [CrossRef]
- Zheng, Z.; Gao, T.; Hou, Y.; Zhou, M. Involvement of the anucleate primary sterigmata protein FgApsB in vegetative differentiation, asexual development, nuclear migration, and virulence in Fusarium graminearum. FEMS Microbiol. Lett. 2013, 349, 88–98. [Google Scholar] [CrossRef]
- Ji, F.; Xu, J.; Liu, X.; Yin, X.; Shi, J. Natural occurrence of deoxynivalenol and zearalenone in wheat from Jiangsu province, China. Food Chem. 2014, 157, 393–397. [Google Scholar] [CrossRef]
- Okegbe, C.; Sakhtah, H.; Sekedat, M.D.; Price-Whelan, A.; Dietrich, L.E. Redox eustress: Roles for redox-active metabolites in bacterial signaling and behavior. Antioxid. Redox. Signal. 2012, 16, 658–667. [Google Scholar] [CrossRef]
- Park, H.J.; Reiser, C.O.; Kondruweit, S.; Erdmann, H.; Schmid, R.D.; Sprinzl, M. Purification and characterization of a NADH oxidase from the thermophile thermus thermophilus HB8. Eur. J. Biochem. 1992, 205, 881–885. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungicide | EC50 (μg/mL) a | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2021 | BM-1 | BM-4 | BM-13 | BM-14 | BM-17 | BM-20 | BM-2 | BM-3 | BM-5 | BM-7 | BM-9 | BM-10 | |
| Fluopyram | 10.00 ± 1.22 | 9.29 ± 0.32 | 8.83 ± 0.35 | 1.65 ± 0.17 | 9.68 ± 0.40 | 7.79 ± 0.41 | 2.29 ± 0.26 | 6.65 ± 0.38 | 2.25 ± 0.31 | 9.65 ± 0.02 | 5.19 ± 0.23 | 6.32 ± 0.30 | 7.54 ± 0.20 |
| Flutolanil | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 |
| Boscalid | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 |
| Benzovindiflupyr | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 |
| Fluxapyroxad | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 | >100 |
| Fungicide | EC50 (μg/mL) a | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2021 | BM-1 | BM-4 | BM-13 | BM-14 | BM-17 | BM-20 | BM-2 | BM-3 | BM-5 | BM-7 | BM-9 | BM-10 | |
| Fluopyram | 0.54 ± 0.11 | 0.74 ± 0.08 | 0.66 ± 0.08 | 0.39 ± 0.05 | 0.67 ± 0.06 | 0.56 ± 0.04 | 0.56 ± 0.06 | 0.54 ± 0.03 | 0.52 ± 0.05 | 0.68 ± 0.08 | 0.54 ± 0.05 | 0.69 ± 0.14 | 0.61 ± 0.03 |
| Flutolanil | 2.96 ± 0.21 | 3.23 ± 0.06 | 2.68 ± 0.07 | 2.47 ± 0.04 | 3.52 ± 0.10 | 3.01 ± 0.04 | 2.33 ± 0.03 | 2.84 ± 0.01 | 2.78 ± 0.01 | 4.24 ± 0.05 | 2.50 ± 0.07 | 2.99 ± 0.07 | 2.32 ± 0.04 |
| Boscalid | 1.52 ± 0.11 | 1.78 ± 0.08 | 2.58 ± 0.07 | 1.19 ± 0.05 | 2.11 ± 0.04 | 2.09 ± 0.03 | 1.57 ± 0.07 | 2.07 ± 0.03 | 1.28 ± 0.09 | 2.41 ± 0.10 | 3.06 ± 0.06 | 2.37 ± 0.06 | 2.40 ± 0.05 |
| Benzovindiflupyr | 1.79 ± 0.08 | 2.86 ± 0.09 | 2.43 ± 0.06 | 1.96 ± 0.04 | 2.02 ± 0.07 | 2.39 ± 0.05 | 2.20 ± 0.03 | 2.59 ± 0.04 | 2.03 ± 0.03 | 2.98 ± 0.08 | 2.15 ± 0.03 | 2.92 ± 0.06 | 2.74 ± 0.06 |
| Fluxapyroxad | 3.16 ± 0.05 | 2.95 ± 0.05 | 3.00 ± 0.04 | 2.30 ± 0.05 | 3.72 ± 0.06 | 3.99 ± 0.04 | 2.08 ± 0.04 | 3.19 ± 0.05 | 3.35 ± 0.06 | 3.78 ± 0.06 | 3.36 ± 0.06 | 3.15 ± 0.02 | 3.27 ± 0.05 |
| Primer | Sequence (5′-3′) | Use |
|---|---|---|
| FGSG_TRI5-qF | CACTTGTCAAGGAGCACTTTC | qRT-PCR for determining the expression of TRI5 (FGSG_03537) |
| FGSG_TRI5-qR | TGCTCAATCCAACATCCCTC | |
| Actin-qF | ATCCACGTCACCACTTTCAA | qRT-PCR for inter-reference actin |
| Actin-qR | TGCTTGGAGATCCACATTTG | |
| FGSG_00500-qF | CGACCTCCACGACAACA | qRT-PCR for determining the expression of hexokinase (FGSG_00500) |
| FGSG_00500-qR | GATAGCAGCAACGCCACA | |
| FGSG_09456-qF | GATAGATTGGAGAGCCGAGAGA | qRT-PCR for determining the expression of 6-phosphate fructokinase (FGSG_09456) |
| FGSG_09456-qR | GAGGTGCTGGATACACTTGATG | |
| FGSG_07528-qF | GAGATCCGAACTGGTAAGACTC | qRT-PCR for determining the expression of pyruvate kinase (FGSG_07528) |
| FGSG_07528-qR | CGTCAGAAGCGGTAGCATAA | |
| TRI1-GFP-F | ACTCACTATAGGGCGAATTGGGTACTCAAATTGGTTTTGTGAGTAGGCCTCATA | A pair of PCR primers to amplify TRI1(FGSG_00071) fragments used for construction of the TRI1-GFP, vector under its own promoter |
| TRI1-GFP-R | CACCACCCCGGTGAACAGCTCCTCGCCCTTGCTCACGTCATCCTGTACCAATTCCAATCG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Li, M.; Zhou, Z.; Li, J.; Chen, D.; Duan, Y.; Zhou, M. Impact of Five Succinate Dehydrogenase Inhibitors on DON Biosynthesis of Fusarium asiaticum, Causing Fusarium Head Blight in Wheat. Toxins 2019, 11, 272. https://doi.org/10.3390/toxins11050272
Xu C, Li M, Zhou Z, Li J, Chen D, Duan Y, Zhou M. Impact of Five Succinate Dehydrogenase Inhibitors on DON Biosynthesis of Fusarium asiaticum, Causing Fusarium Head Blight in Wheat. Toxins. 2019; 11(5):272. https://doi.org/10.3390/toxins11050272
Chicago/Turabian StyleXu, Chao, Meixia Li, Zehua Zhou, Jiaosheng Li, Dongming Chen, Yabing Duan, and Mingguo Zhou. 2019. "Impact of Five Succinate Dehydrogenase Inhibitors on DON Biosynthesis of Fusarium asiaticum, Causing Fusarium Head Blight in Wheat" Toxins 11, no. 5: 272. https://doi.org/10.3390/toxins11050272
APA StyleXu, C., Li, M., Zhou, Z., Li, J., Chen, D., Duan, Y., & Zhou, M. (2019). Impact of Five Succinate Dehydrogenase Inhibitors on DON Biosynthesis of Fusarium asiaticum, Causing Fusarium Head Blight in Wheat. Toxins, 11(5), 272. https://doi.org/10.3390/toxins11050272