The Ecological Importance of Toxicity: Sea Anemones Maintain Toxic Defence When Bleached


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
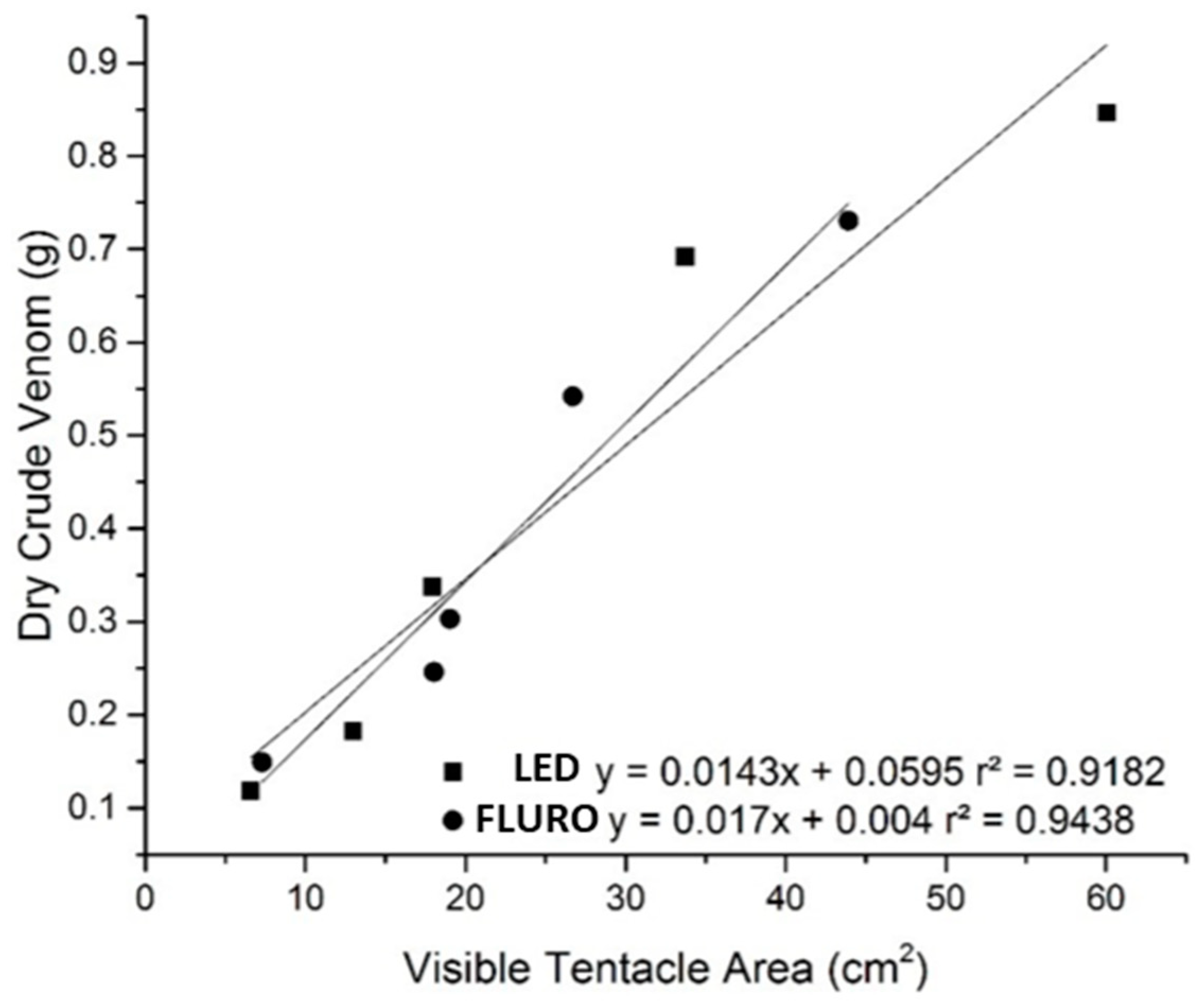
2.1. Anemone Bleaching
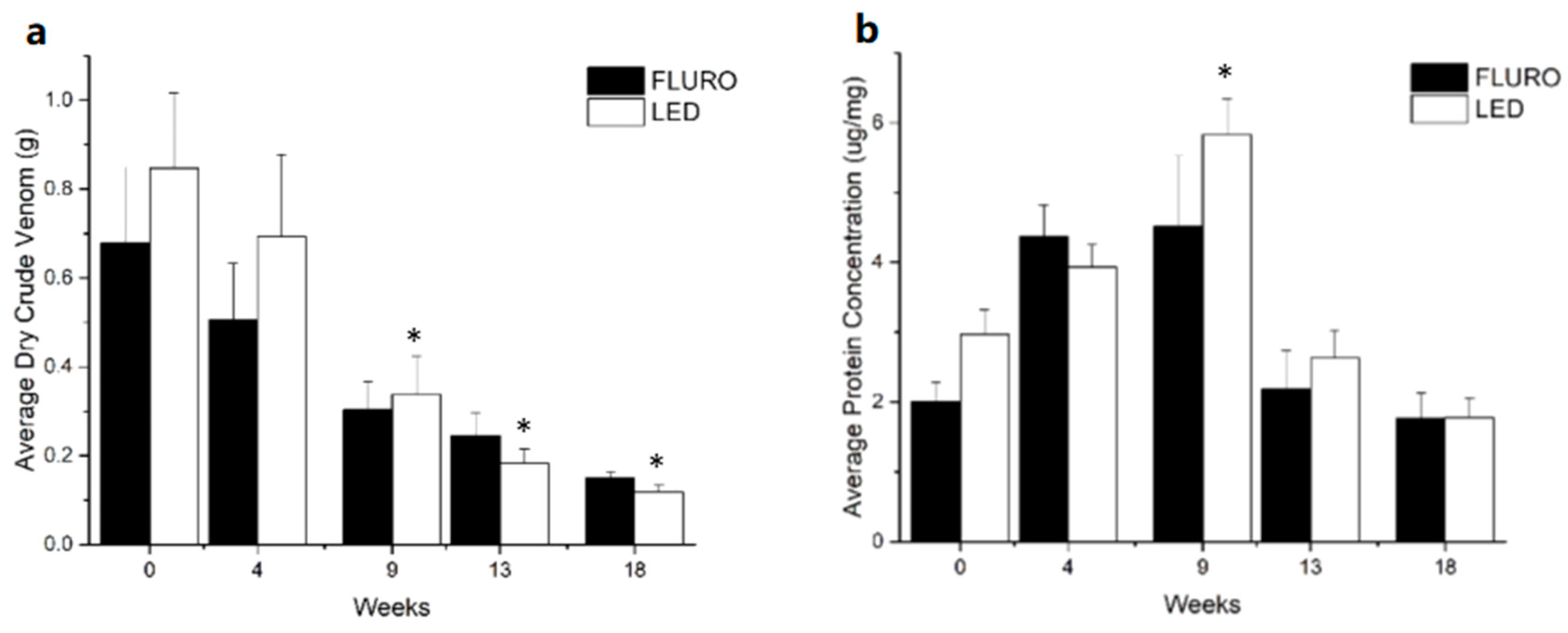
2.2. Venom Characterisation
2.3. Protein Composition of Venom
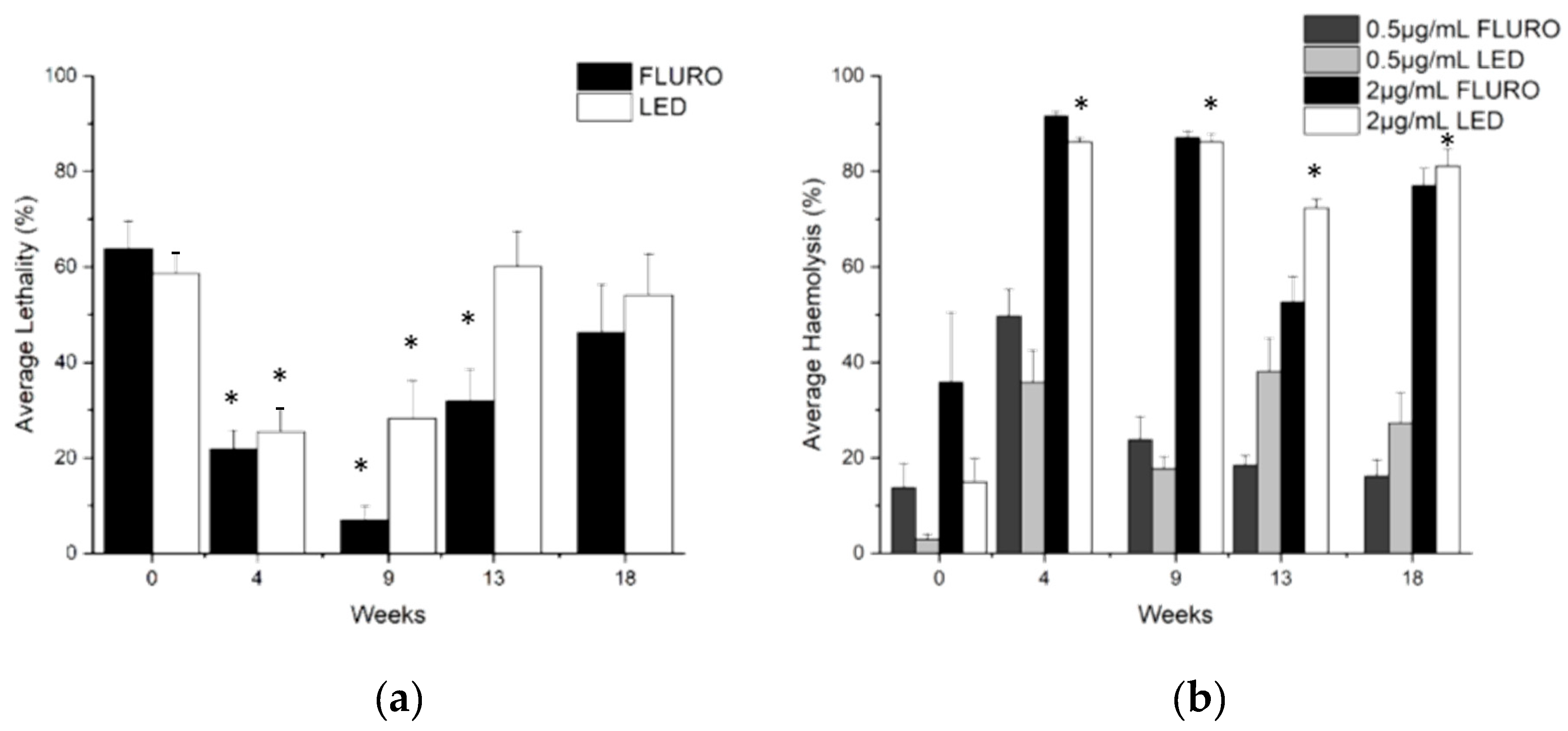
2.4. Lethality and Haemolysis
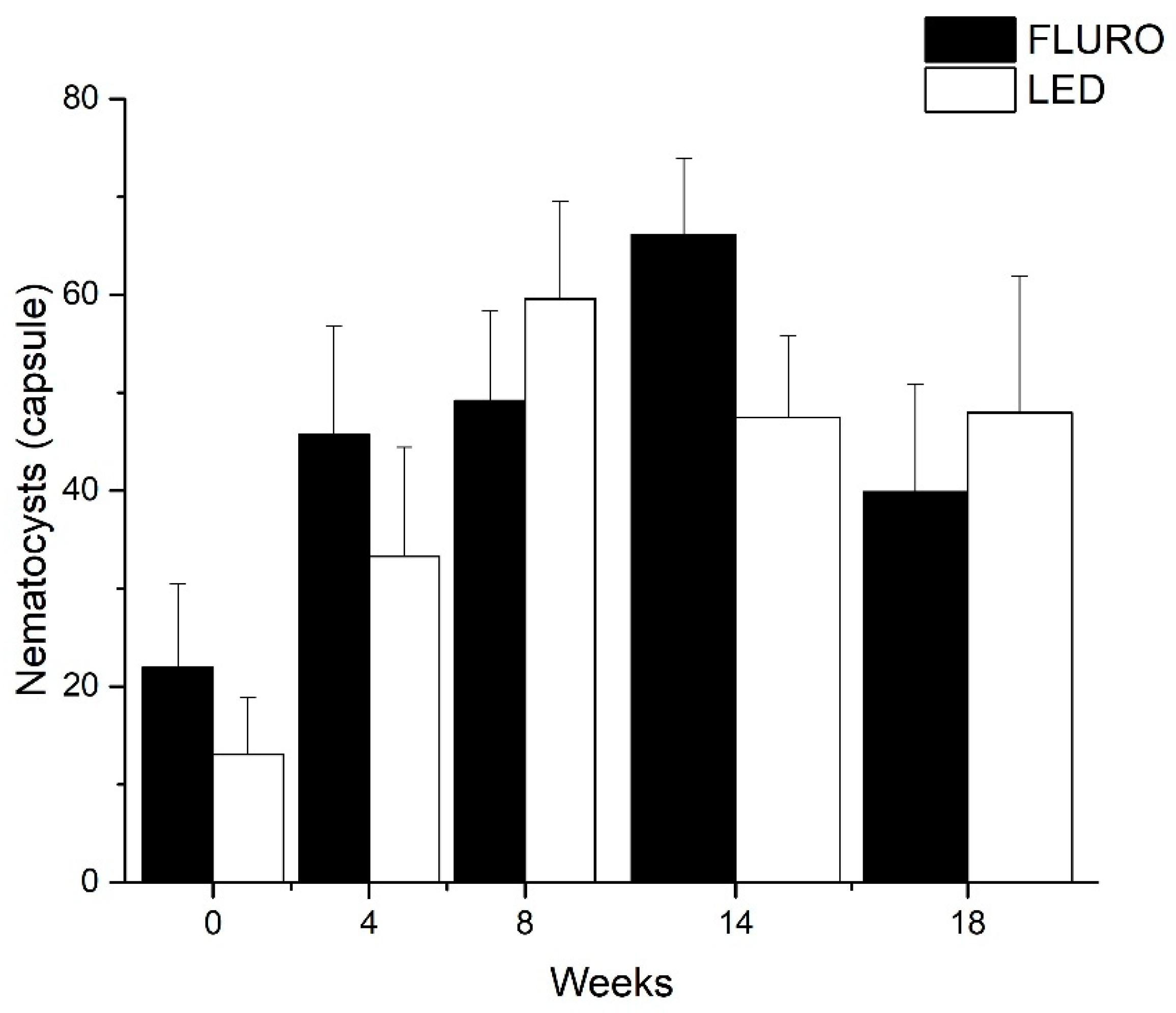
2.5. Nematocysts
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Bleaching of Anemones
5.2. Venom Collection
5.3. Venom Characterisation
5.4. Electrophoresis
5.5. Artemia Lethality Assay
5.6. Haemolysis Assay
5.7. Nematocyst Sampling and Quantification
5.8. Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hutton, D.; Smith, V.J. Antibacterial properties of isolated amoebocytes from the sea anemone Actinia equina. Biol. Bull. 1996, 191, 441–451. [Google Scholar] [CrossRef]
- Bailey, P.M.; Bakker, A.J.; Seymour, J.E.; Wilce, J.A. A functional comparison of the venom of three Australian jellyfish-Chironex fleckeri, Chiropsalmus sp., and Carybdea xaymacana- on cytosolic Ca2+, haemolysis and Artemia sp. lethality. Toxicon 2005, 45, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Frazao, B.; Vasconcelos, V.; Antunes, A. Sea anemone (Cnidaria, Anthozoa, Actiniaria) toxins: An overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef] [PubMed]
- Anderluh, G.; Maček, P. Cytolytic peptide and protein toxins from sea anemones (Anthozoa: Actiniaria). Toxicon 2002, 40, 111–124. [Google Scholar] [CrossRef]
- Ramezanpour, M.; da Silva, K.B.; Sanderson, B.J. The effect of sea anemone (H. magnifica) venom on two human breast cancer lines: Death by apoptosis. Cytotechnology 2014, 66, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Tytgat, J. Sea anemone toxins: From lab bench discovery to commercial drugs? Toxicon 2014, 91, 170. [Google Scholar] [CrossRef]
- Norton, R.S. Structures of sea anemone toxins. Toxicon 2009, 54, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Nedosyko, A.M.; Young, J.E.; Edwards, J.W.; Burke da Silva, K. Searching for a toxic key to unlock the mystery of anemonefish and anemone symbiosis. PLoS ONE 2014, 9, e98449. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, A.; Ozbek, S. The nematocyst: A molecular map of the cnidarian stinging organelle. Int. J. Dev. Biol. 2012, 56, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Mebs, D. Chemical biology of the mutualistic relationships of sea anemones with fish and crustaceans. Toxicon 2009, 54, 1071–1074. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Gordon, D.; Gurevitz, M. Sea anemone toxins affecting voltage-gated sodium channels--molecular and evolutionary features. Toxicon 2009, 54, 1089–1101. [Google Scholar] [CrossRef] [PubMed]
- Dixon, A.K.; Needham, D.; Al-Horani, F.A.; Chadwick, N.E. Microhabitat use and photoacclimation in the clownfish sea anemone Entacmaea quadricolor. J. Mar. Biol. Assoc. U. K. 2013, 94, 473–480. [Google Scholar] [CrossRef]
- Jones, A.M.; Gardner, S.; Sinclair, W. Losing ‘Nemo’: Bleaching and collection appear to reduce inshore populations of anemonefishes. J. Fish Biol. 2008, 73, 753–761. [Google Scholar] [CrossRef]
- Baker, A.C.; Glynn, P.W.; Riegl, B. Climate change and coral reef bleaching: An ecological assessment of long-term impacts, recovery trends and future outlook. Estuar. Coast. Shelf Sci. 2008, 80, 435–471. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O. Climate Change, coral bleaching and the furture of the world’s coral reefs. Mar. Fresh. Res. 1999, 50, 839–866. [Google Scholar] [CrossRef]
- Brown, B.E. Coral Bleaching: Causes and consequences. Coral Reefs 1997, 16, 129–138. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef]
- Weis, V.M. Cellular mechanisms of cnidarian bleaching: Stress causes the collapse of symbiosis. J. Exp. Biol. 2008, 211, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Pontasch, S.; Scott, A.; Hill, R.; Bridge, T.; Fisher, P.L.; Davy, S.K. Symbiodinium diversity in the sea anemone Entacmaea quadricolor on the east Australian coast. Coral Reefs 2013, 33, 537–542. [Google Scholar] [CrossRef]
- Gates, R.D.; Baghdasarian, G.; Muscatine, L. Temperature stress causes host cell detachment in symbiotic Cnidarians: Implications for coral bleaching. Biol. Bull. 1992, 182, 324–332. [Google Scholar] [CrossRef]
- Saenz-Agudelo, P.; Jones, G.P.; Thorrold, S.R.; Planes, S. Detrimental effects of host anemone bleaching on anemonefish populations. Coral Reefs 2011, 30, 497–506. [Google Scholar] [CrossRef]
- Hobbs, J.P.; Frisch, A.J.; Ford, B.M.; Thums, M.; Saenz-Agudelo, P.; Furby, K.A.; Berumen, M.L. Taxonomic, spatial and temporal patterns of bleaching in anemones inhabited by anemonefishes. PLoS ONE 2012, 8, e70966. [Google Scholar] [CrossRef] [PubMed]
- Conklin, E.J.; Mariscal, R.N. Increase in nematocyst and spirocyst discharge in a sea anemone in response to mechanical stimulation. In Coelenterate Ecology and Behavior; Mackie, G.O., Ed.; Springer: Boston, MA, USA, 1976; pp. 549–558. [Google Scholar] [CrossRef]
- Lonnstedt, O.M.; Frisch, A.J. Habitat bleaching disrupts threat responses and persistence in anemonefish. Mar. Ecol. Prog. Ser. 2014, 517, 265–270. [Google Scholar] [CrossRef]
- Hill, R.; Fernance, C.; Wilkinson, S.P.; Davy, S.K.; Scott, A. Symbiont shuffling during thermal bleaching and recovery in the sea anemone Entacmaea quadricolor. Mar. Biol. 2014, 161, 2931–2937. [Google Scholar] [CrossRef]
- Hill, R.; Scott, A. The influence of irradiance on the severity of thermal bleaching in sea anemones that host anemonefish. Coral Reefs 2012, 31, 273–284. [Google Scholar] [CrossRef]
- Malcolm, H.; Scott, A. Range extensions in anemonefishes and host sea anemones in eastern Australia: Potential constraints to tropicalisation. Mar. Freshw. Res. 2017, 68, 1224–1232. [Google Scholar] [CrossRef]
- Reitzel, A.M.; Chu, T.; Edquist, S.; Genovese, C.; Church, C.; Tarrant, A.M.; Finnerty, J.R. Physiological and developmental responses to temperature by the sea anemone Nematostella vectensis. Mar. Ecol. Prog. Ser. 2013, 484, 115–130. [Google Scholar] [CrossRef]
- Pontasch, S.; Hill, R.; Deschaseaux, E.; Fisher, P.L.; Davy, S.K.; Scott, A. Photochemical efficiency and antioxidant capacity in relation to Symbiodinium genotype and host phenotype in a symbiotic cnidarian. Mar. Ecol. Prog. Ser. 2014, 516, 195–208. [Google Scholar] [CrossRef]
- Hoepner, C.M. Irradiance and Toxcity: Does Bleaching Effect Venom Toxicity in Anemones? First Class Honours Thesis, Flinders University, Adelaide, South Australia, 1 May 2017. [Google Scholar]
- Takahashi, S.; Nakamura, T.; Sakamizu, M.; van Woesik, R.; Yamasaki, H. Repair machinery of symbiotic photosynthesis as the primary target of heat stress for reef-building corals. Plant Cell Physiol. 2004, 45, 251–255. [Google Scholar] [CrossRef]
- García-Arredondo, A.; Rojas-Molina, A.; Ibarra-Alvarado, C.; Iglesias-Prieto, R. Effects of bleaching on the pharmacological and toxicological activities elicited by the aqueous extracts prepared from two “fire corals” collected in the Mexican Caribbean. J. Exp. Mar. Biol. Ecol. 2011, 396, 171–176. [Google Scholar] [CrossRef]
- Miller, J.; Muller, E.; Rogers, C.; Waara, R.; Atkinson, A.; Whelan, K.R.T.; Patterson, M.; Witcher, B. Coral disease following massive bleaching in 2005 causes 60% decline in coral cover on reefs in the US Virgin Islands. Coral Reefs 2009, 28, 925–937. [Google Scholar] [CrossRef]
- Bruno, J.F.; Selig, E.R.; Casey, K.S.; Page, C.A.; Willis, B.L.; Harvell, C.D.; Sweatman, H.; Melendy, M. Thermal Stress and Coral Cover as Drivers of Coral Disease Outbreaks. PLoS ONE 2007, 5, e124. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Alvarez-Noriega, M.; Alvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef]
- Bates, A.E.; McLean, L.; Laing, P.; Raeburn, L.A.; Hare, C. Distribution patterns of zoochlorellae and zooxanthellae hosted by two Pacific Northeast anemones, Anthopleura elegantissima and A. xanthogrammica. Biol. Bull. 2010, 218, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Bucklin, A. Adaptive advantages of patterns of growth and asexual reproduction of the sea anemone Metridium senile in intertidal and submerged populations. J. Exp. Mar. Biol. Ecol. 1987, 110, 225–243. [Google Scholar] [CrossRef]
- Molina, V.H.; Castillo-Medina, R.E.; Thomé, P.E. Experimentally induced bleaching in the sea anemone Exaiptasia supports glucose as a main metabolite associated with its symbiosis. J. Mar. Biol. 2017, 2017, 1–7. [Google Scholar] [CrossRef]
- Sencic, L.; Macek, P. New method for isolation of venom from the sea anemone Actinia cari purification and characterization of cytolytic toxins. Comp. Biochem. Phys. B 1990, 97, 687–693. [Google Scholar] [CrossRef]
- Li, Y.; Qin, J.G.; Abbott, C.A.; Li, X.; Benkendorff, K. Synergistic impacts of heat shock and spawning on the physiology and immune health of Crassostrea gigas: An explanation for summer mortality in Pacific oysters. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R2353–R2362. [Google Scholar] [CrossRef]
- Meakin, C.A.; Qin, J.G.; Pogson, L.D.; Abbott, C.A. Thermal tolerance in juvenile King George whiting (Sillaginodes punctata) reduces as fish age and this reduction coincides with migration to deeper colder water. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2014, 172, 46–51. [Google Scholar] [CrossRef]
- Yang, C.Y.; Sierp, M.T.; Abbott, C.A.; Li, Y.; Qin, J.G. Responses to thermal and salinity stress in wild and farmed Pacific oysters Crassostrea gigas. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2016, 201, 22–29. [Google Scholar] [CrossRef]
- Feder, M.E.; Hofman, G.E. Heat-shock proteins, molecular chaperones, and the stress responce: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [PubMed]
- Sharp, V.A.; Miller, D.; Bythell, J.C.; Brown, B.E. Expression of low molecular weight HSP 70 related polypeptides from the symbiotic sea anemone Anemonia viridis in response to heat shock. J. Exp. Mar. Biol. Ecol. 1994, 179, 179–193. [Google Scholar] [CrossRef]
- Choresh, O.; Ron, E.; Loya, Y. The 60-kDa heat shock protein (HSP60) of the sea anemone Anemonia viridis: a potential early warning system for environmental changes. Mar. Biotechnol. (NY) 2001, 3, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Cubillos, V.M.; Lamare, M.D.; Peake, B.M.; Burritt, D.J. Cellular changes associated with the acclimation of the intertidal sea anemone Actinia tenebrosa to ultraviolet radiation. Photochem. Photobiol. 2014, 90, 1314–1323. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671. [Google Scholar] [CrossRef] [PubMed]
- Chow, M.H.; Tsang, R.H.L.; Lam, E.K.Y.; Ang, P. Quantifying the degree of coral bleaching using digital photographic technique. J. Exp. Mar. Biol. Ecol. 2016, 479, 60–68. [Google Scholar] [CrossRef]
- Schagger, H.; von Jagow, G. Tricine sodium dodecyl sulfate polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef]
- Vanhaecke, P.; Persoone, G. The ARC Test: A Standardised Short-Term Routine Toxcitiy Test with Artemia Nauplii: Methedology and Evaluation. Available online: http://agris.fao.org/agris-search/search.do?recordID=AV20120151533 (accessed on 15 September 2016).
- Evans, B.C.; Nelson, C.E.; Yu, S.S.; Beavers, K.R.; Kim, A.J.; Li, H.; Nelson, H.M.; Giorgio, T.D.; Duvall, C.L. Ex vivo red blood cell hemolysis assay for the evaluation of pH-responsive endosomolytic agents for cytosolic delivery of biomacromolecular drugs. J. Vis. Exp. 2013, 73, e50166. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoepner, C.M.; Abbott, C.A.; Burke da Silva, K. The Ecological Importance of Toxicity: Sea Anemones Maintain Toxic Defence When Bleached. Toxins 2019, 11, 266. https://doi.org/10.3390/toxins11050266
Hoepner CM, Abbott CA, Burke da Silva K. The Ecological Importance of Toxicity: Sea Anemones Maintain Toxic Defence When Bleached. Toxins. 2019; 11(5):266. https://doi.org/10.3390/toxins11050266
Chicago/Turabian StyleHoepner, Cassie M., Catherine A. Abbott, and Karen Burke da Silva. 2019. "The Ecological Importance of Toxicity: Sea Anemones Maintain Toxic Defence When Bleached" Toxins 11, no. 5: 266. https://doi.org/10.3390/toxins11050266
APA StyleHoepner, C. M., Abbott, C. A., & Burke da Silva, K. (2019). The Ecological Importance of Toxicity: Sea Anemones Maintain Toxic Defence When Bleached. Toxins, 11(5), 266. https://doi.org/10.3390/toxins11050266