Mycotoxins during the Processes of Nixtamalization and Tortilla Production

Abstract
1. Introduction
2. Processes Involved in Nixtamalization and Tortilla Production
3. Aflatoxins during Nixtamalization and Tortilla Production
3.1. Impact on Aflatoxin Concentrations by Traditional Nixtamalization
3.2. Aflatoxin Reductions by Alternative Nixtamalization Processes
3.3. Potential Reconversion of Modified Aflatoxins
4. Fumonisins during Nixtamalization and Tortilla Production
4.1. General Impact on Fumonisin Concentrations and Fumonisin Hydrolyzation
4.2. Potential Further Transformations of Fumonisins
5. Other Mycotoxins during Nixtamalization and Tortilla Production
6. Conclusions and Outlook
Funding
Acknowledgments
Conflicts of Interest
References
- Pitt, J.; Wild, C.; Baan, R.; Gelderblom, W.; Miller, J.; Riley, R.; Wu, F. Improving Public Health through Mycotoxin Control. Chapter 1: Fungi Producing Significant Mycotoxins; International Agency for Research on Cancer, WHO Press: Geneva, Switzerland, 2012; pp. 1–30. [Google Scholar]
- Taniwaki, M.H.; Pitt, J.I.; Magan, N. Aspergillus species and mycotoxins: Occurrence and importance in major food commodities. Curr. Opin. Food Sci. 2018, 23, 38–43. [Google Scholar] [CrossRef]
- EFSA CONTAM. Opinion of the scientific panel on contaminants in the food chain [CONTAM] related to the potential increase of consumer health risk by a possible increase of the existing maximum levels for aflatoxins in almonds, hazelnuts and pistachios and derived products. EFSA J. 2007, 446, 1–127. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain; Knutsen, H.-K.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; et al. Appropriateness to set a group health-based guidance value for fumonisins and their modified forms. EFSA J. 2018, 16, e05172. [Google Scholar]
- Metzler, M. Proposal for a uniform designation of zearalenone and its metabolites. Mycotoxin Res. 2011, 27, 1–3. [Google Scholar] [CrossRef] [PubMed]
- EFSA CONTAM. Scientific Opinion on the risks for public health related to the presence of zearalenone in food. EFSA J. 2011, 9, 2197. [Google Scholar] [CrossRef]
- EFSA. Deoxynivalenol in food and feed: Occurrence and exposure. EFSA J. 2013, 11, 3379. [Google Scholar]
- Jestoi, M. Emerging Fusarium-mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin—A review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef] [PubMed]
- Kovač, M.; Šubarić, D.; Bulaić, M.; Kovač, T.; Šarkanj, B. Yesterday masked, today modified; what do mycotoxins bring next? Arh. Hig. Rada Toksikol. 2018, 69, 196–214. [Google Scholar] [CrossRef] [PubMed]
- Dall’Asta, C.; Berthiller, F. Masked Mycotoxins in Food: Formation, Occurrence and Toxicological Relevance; Royal Society of Chemistry: Cambridge, UK, 2016. [Google Scholar]
- Rychlik, M.; Humpf, H.-U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Rooney, L.W.; Serna-Saldivar, S.O. Tortillas. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Serna-Saldivar, S.O. Chapter 1—History of corn and wheat tortillas. In Tortillas; Rooney, L.W., Serna-Saldivar, S.O., Eds.; AACC International Press: St. Paul, MN, USA, 2015; pp. 1–28. [Google Scholar]
- Santiago-Ramos, D.; Figueroa-Cárdenas, J.d.D.; Mariscal-Moreno, R.M.; Escalante-Aburto, A.; Ponce-García, N.; Véles-Medina, J.J. Physical and chemical changes undergone by pericarp and endosperm during corn nixtamalization—A review. J. Cereal Sci. 2018, 81, 108–117. [Google Scholar] [CrossRef]
- Serna-Saldivar, S.O. Chapter 2—Nutrition and fortification of corn and wheat tortillas. In Tortillas; Rooney, L.W., Serna-Saldivar, S.O., Eds.; AACC International Press: St. Paul, MN, USA, 2015; pp. 29–63. [Google Scholar]
- Bressani, R.; Scrimshaw, N.S. Lime-heat effects on corn nutrients, effect of lime treatment on in vitro availability of essential amino acids and solubility of protein fractions in corn. J. Agric. Food Chem. 1958, 6, 774–778. [Google Scholar] [CrossRef]
- Rendon-Villalobos, R.; Bello-Pérez, L.A.; Osorio-Díaz, P.; Tovar, J.; Paredes-López, O. Effect of storage time on in vitro digestibility and resistant starch content of nixtamal, masa, and tortilla. Cereal Chem. 2002, 79, 340–344. [Google Scholar] [CrossRef]
- Mariscal-Moreno, R.M.; de Dios Figueroa Cárdenas, J.; Santiago-Ramos, D.; Rayas-Duarte, P.; Veles-Medina, J.J.; Martínez-Flores, H.E. Nixtamalization process affects resistant starch formation and glycemic index of tamales. J. Food Sci. 2017, 82, 1110–1115. [Google Scholar] [CrossRef]
- Pilcher, J. Taste, Smell, and Flavor in Mexico. In Oxford Research Encyclopedia of Latin American History; Oxford University Press: New York, NY, USA, 2016; Available online: http://oxfordre.com/latinamericanhistory/ (accessed on 5 April 2019).
- de Arriola, M.d.C.; de Porres, E.; de Cabrera, S.; de Zepeda, M.; Rolz, C. Aflatoxin fate during alkaline cooking of corn for tortilla preparation. J. Agric. Food Chem. 1988, 36, 530–533. [Google Scholar] [CrossRef]
- Ulloa-Sosa, M.; Schroeder, H.W. Note on aflatoxin decomposition in the process of making tortillas from corn. Cereal Chem. 1969, 46, 397–400. [Google Scholar]
- Abbas, H.K.; Mirocha, C.J.; Rosiles, R.; Carvajal, M. Effect of tortilla-preparation process on aflatoxins B1 and B2 in corn. Mycotoxin Res. 1988, 4, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Price, R.L.; Jorgensen, K.V. Effects of processing on aflatoxin levels and on mutagenic potential of tortillas made from naturally contaminated corn. J. Food Sci. 1985, 50, 347–349. [Google Scholar] [CrossRef]
- Moreno-Pedraza, A.; Valdés-Santiago, L.; Hernández-Valadez, L.J.; Rodríguez-Sixtos Higuera, A.; Winkler, R.; Guzmán-de Peña, D.L. Reduction of aflatoxin B1 during tortilla production and identification of degradation by-products by direct-injection electrospray mass spectrometry. Salud Pública de México 2015, 57, 50–56. [Google Scholar] [CrossRef]
- Guzman-de-Pena, D.; Trudel, L.; Wogan, G.N. Corn “nixtamalización” and the fate of radiolabelled aflatoxin B1 in the tortilla making process. Bull. Environ. Contam. Toxicol. 1995, 55, 858–864. [Google Scholar] [CrossRef]
- Pérez-Flores, G.C.; Moreno-Martínez, E.; Méndez-Albores, A. Effect of microwave heating during alkaline-cooking of aflatoxin contaminated maize. J. Food Sci. 2011, 76, T48–T52. [Google Scholar] [CrossRef]
- Rosentrater, K.A. A review of corn masa processing residues: Generation, properties, and potential utilization. Waste Manag. 2006, 26, 284–292. [Google Scholar] [CrossRef]
- Bazúa, C.D.; Guerra, R.; Sterner, H. Extruded corn flour as an alternative to lime-heated corn flour for tortilla preparation. J. Food Sci. 1979, 44, 940–941. [Google Scholar] [CrossRef]
- Gómez-Aldapa, C.; Martínez-Bustos, F.; Figueroa, C.J.D.; Ordorica, F.C.A. A comparison of the quality of whole corn tortillas made from instant corn flours by traditional or extrusion processing. Int. J. Food Sci. Technol. 1999, 34, 391–399. [Google Scholar] [CrossRef]
- Elias-Orozco, R.; Castellanos-Nava, A.; Gaytán-Martínez, M.; Figueroa-Cárdenas, J.D.; Loarca-Piña, G. Comparison of nixtamalization and extrusion processes for a reduction in aflatoxin content. Food Addit. Contam. 2002, 19, 878–885. [Google Scholar] [CrossRef]
- Lutz, W.K.; Jaggi, W.; Lüthy, J.; Sagelsdorff, P.; Schlatter, C. In vivo covalent binding of aflatoxin B1 and aflatoxin M1 to liver DNA of rat, mouse and pig. Chem.-Biol. Interact. 1980, 32, 249–256. [Google Scholar] [CrossRef]
- Pallavi, R.M.V.; Vidyasagar, T.; Rao, B. Production of aflatoxin M1 by Aspergillus parasiticus (AP 456) in a semisynthetic medium. Indian J. Exp. Biol. 1997, 35, 735–741. [Google Scholar]
- Pai, M.R.; Bai, N.J.; Venkitasubramanian, T.A. Production of aflatoxin M in a liquid medium. Appl. Microbiol. 1975, 29, 850–851. [Google Scholar] [PubMed]
- Vesonder, R.; Haliburton, J.; Stubblefield, R.; Gilmore, W.; Peterson, S. Aspergillus flavus and aflatoxins B1, B2, and M1 in corn associated with equine death. Arch. Environ. Contam. Toxicol. 1991, 20, 151–153. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Albores, J.A.; Arámbula-Villa, G.; Loarca-Piña, M.G.; González-Hernández, J.; Castaño-Tostado, E.; Moreno-Martínez, E. Aflatoxins’ fate during the nixtamalization of contaminated maize by two tortilla-making processes. J. Stored Prod. Res. 2004, 40, 87–94. [Google Scholar] [CrossRef]
- Torres, P.; Guzmán-Ortiz, M.; Ramírez-Wong, B. Revising the role of pH and thermal treatments in aflatoxin content reduction during the tortilla and deep frying processes. J. Agric. Food Chem. 2001, 49, 2825–2829. [Google Scholar] [CrossRef]
- Lee, L.S.; Dunn, J.J.; DeLucca, A.J.; Ciegler, A. Role of lactone ring of aflatoxin B1 in toxicity and mutagenicity. Experientia 1981, 37, 16–17. [Google Scholar] [CrossRef]
- Vázquez-Durán, A.; Díaz-Torres, R.; Ramírez-Noguera, P.; Moreno-Martínez, E.; Méndez-Albores, A. Cytotoxic and genotoxic evaluation of tortillas produced by microwave heating during alkaline-cooking of aflatoxin-contaminated maize. J. Food Sci. 2014, 79, T1024–T1029. [Google Scholar] [CrossRef]
- Méndez-Albores, J.; Villa, G.; Del Rio-García, J.; Martínez, E. Aflatoxin-detoxification achieved with Mexican traditional nixtamalization process (MTNP) is reversible. J. Sci. Food Agric. 2004, 84, 1611–1614. [Google Scholar] [CrossRef]
- Beckwith, A.C.; Vesonder, R.F.; Ciegler, A. Action of weak bases upon aflatoxin B1 in contact with macromolecular reactants. J. Agric. Food Chem. 1975, 23, 582–587. [Google Scholar] [CrossRef]
- Lee, L.S.; Conkerton, E.J.; Ory, R.L.; Bennett, J.W. [14C]Aflatoxin B1 as an indicator of toxin destruction during ammoniation of contaminated peanut meal. J. Agric. Food Chem. 1979, 27, 598–602. [Google Scholar] [CrossRef]
- Brekke, O.L.; Peplinski, A.J.; Lancaster, E.B. Aflatoxin inactivation in corn by aqua ammonia. Trans. ASAE 1977, 20, 1160–1168. [Google Scholar] [CrossRef]
- Weng, C.Y.; Martinez, A.J.; Park, D.L. Efficacy and permanency of ammonia treatment in reducing aflatoxin levels in corn. Food Addit. Contam. 1994, 11, 649–658. [Google Scholar] [CrossRef]
- Sydenham, E.W.; Stockenstrom, S.; Thiel, P.G.; Shephard, G.S.; Koch, K.R.; Marasas, W.F.O. Potential of alkaline hydrolysis for the removal of fumonisins from contaminated corn. J. Agric. Food Chem. 1995, 43, 1198–1201. [Google Scholar] [CrossRef]
- Voss, K.A.; Poling, S.M.; Meredith, F.I.; Bacon, C.W.; Saunders, D.S. Fate of fumonisins during the production of fried tortilla chips. J. Agric. Food Chem. 2001, 49, 3120–3126. [Google Scholar] [CrossRef]
- Dombrink-Kurtzman, M.A.; Dvorak, T.J.; Barron, M.E.; Rooney, L.W. Effect of nixtamalization (alkaline cooking) on fumonisin-contaminated corn for production of masa and tortillas. J. Agric. Food Chem. 2000, 48, 5781–5786. [Google Scholar] [CrossRef]
- Palencia, E.; Torres, O.; Hagler, W.; Meredith, F.I.; Williams, L.D.; Riley, R.T. Total fumonisins are reduced in tortillas using the traditional nixtamalization method of mayan communities. J. Nutr. 2003, 133, 3200–3203. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Albores, A.; Cardenas-Rodriguez, D.A.; Vazquez-Duran, A. Efficacy of microwave-heating during alkaline processing of fumonisin-contaminated maize. Iran J. Public Health 2014, 43, 147–155. [Google Scholar]
- Scudamore, K.; Scriven, F.; Patel, S. Fusarium mycotoxins in the food chain: Maize-based snack foods. World Mycotoxin J. 2009, 2, 441–450. [Google Scholar] [CrossRef]
- De La Campa, R.; Miller, J.D.; Hendricks, K. Fumonisin in tortillas produced in small-scale facilities and effect of traditional masa production methods on this mycotoxin. J. Agric. Food Chem. 2004, 52, 4432–4437. [Google Scholar] [CrossRef]
- De Girolamo, A.; Lattanzio, V.M.T.; Schena, R.; Visconti, A.; Pascale, M. Effect of alkaline cooking of maize on the content of fumonisins B1 and B2 and their hydrolysed forms. Food Chem. 2016, 192, 1083–1089. [Google Scholar] [CrossRef]
- Gelderblom, W.C.A.; Cawood, M.E.; Snyman, S.D.; Vleggaar, R.; Marasas, W.F.O. Structure-activity relationships of fumonisins in short-term carcinogenesis and cytotoxicity assays. Food Chem. Toxicol. 1993, 31, 407–414. [Google Scholar] [CrossRef]
- Grenier, B.; Bracarense, A.-P.F.L.; Schwartz, H.E.; Trumel, C.; Cossalter, A.-M.; Schatzmayr, G.; Kolf-Clauw, M.; Moll, W.-D.; Oswald, I.P. The low intestinal and hepatic toxicity of hydrolyzed fumonisin B1 correlates with its inability to alter the metabolism of sphingolipids. Biochem. Pharmacol. 2012, 83, 1465–1473. [Google Scholar] [CrossRef]
- Voss, K.A.; Riley, R.T.; Snook, M.E.; Waes, J.G.-V. Reproductive and sphingolipid metabolic effects of fumonisin B1 and its alkaline hydrolysis product in LM/Bc mice: Hydrolyzed fumonisin B1 did not cause neural tube defects. Toxicol. Sci. 2009, 112, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Scott, P.M.; Lau, B.P.-Y.; Lewis, D.A. Analysis of heat-processed corn foods for fumonisins and bound fumonisins. Food Addit. Contam. 2004, 21, 1168–1178. [Google Scholar] [CrossRef]
- Burns, T.D. Fate of the Mycotoxin Fumonisin B1 During Alkaline Cooking of Cultured and Whole Kernel Corn. Ph.D. Thesis, University of Georgia, Georgia, Athens, 2008. [Google Scholar]
- Cortez-Rocha, M.O.; Trigo-Stockli, D.M.; Wetzel, D.L.; Reed, C.R. Effect of extrusion processing on fumonisin B1 and hydrolyzed fumonisin B1 in contaminated alkali-cooked corn. Bull. Environ. Contam. Toxicol. 2002, 69, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Howard, P.C.; Churchwell, M.I.; Couch, L.H.; Marques, M.M.; Doerge, D.R. Formation of N-(carboxymethyl)fumonisin B1, following the reaction of fumonisin B1 with reducing sugars. J. Agric. Food Chem. 1998, 46, 3546–3557. [Google Scholar] [CrossRef]
- Seefelder, W.; Knecht, A.; Humpf, H.-U. Bound fumonisin B1: Analysis of fumonisin-B1 glyco and amino acid conjugates by liquid chromatography−electrospray ionization−tandem mass spectrometry. J. Agric. Food Chem. 2003, 51, 5567–5573. [Google Scholar] [CrossRef] [PubMed]
- Voss, K.A.; Riley, R.T.; Jackson, L.S.; Jablonski, J.E.; Bianchini, A.; Bullerman, L.B.; Hanna, M.A.; Ryu, D. Extrusion cooking with glucose supplementation of fumonisin-contaminated corn grits protects against nephrotoxicity and disrupted sphingolipid metabolism in rats. Mol. Nutr. Food Res. 2011, 55, S312–S320. [Google Scholar] [CrossRef]
- Park, J.W.; Scott, P.M.; Lau, B.P.Y. Analysis of N-fatty acyl fumonisins in alkali-processed corn foods. Food Sci. Biotechnol. 2013, 22, 147–152. [Google Scholar] [CrossRef]
- Burns, T.D.; Snook, M.E.; Riley, R.T.; Voss, K.A. Fumonisin concentrations and in vivo toxicity of nixtamalized Fusarium verticillioides culture material: Evidence for fumonisin–matrix interactions. Food Chem. Toxicol. 2008, 46, 2841–2848. [Google Scholar] [CrossRef]
- Voss, K.A.; Riley, R.T.; Moore, N.D.; Burns, T.D. Alkaline cooking (nixtamalisation) and the reduction in the in vivo toxicity of fumonisin-contaminated corn in a rat feeding bioassay. Food Addit. Contam. Part A 2013, 30, 1415–1421. [Google Scholar] [CrossRef] [PubMed]
- Hendrich, S.; Miller, K.A.; Wilson, T.M.; Murphy, P.A. Toxicity of Fusarium proliferatum-fermented nixtamalized corn-based diets fed to rats: Effect of nutritional status. J. Agric. Food Chem. 1993, 41, 1649–1654. [Google Scholar] [CrossRef]
- Abbas, H.K.; Mirocha, C.J.; Rosiles, R.; Carvajal, M. Decomposition of zearalenone and deoxynivalenol in the process of making tortillas from corn. Cereal Chem. 1988, 65, 15–19. [Google Scholar]
- Trenholm, H.L.; Charmley, L.L.; Prelusky, D.B.; Warner, R.M. Washing procedures using water or sodium carbonate solutions for the decontamination of three cereals contaminated with deoxynivalenol and zearalenone. J. Agric. Food Chem. 1992, 40, 2147–2151. [Google Scholar] [CrossRef]
- Schaarschmidt, S.; Fauhl-Hassek, C. The fate of mycotoxins during the processing of wheat for human consumption. Compr. Rev. Food Sci. Food Saf. 2018, 17, 556–593. [Google Scholar] [CrossRef]
- Pineda-Valdes, G.; Ryu, D.; Jackson, D.S.; Bullerman, L.B. Reduction of moniliformin during alkaline cooking of corn. Cereal Chem. 2002, 79, 779–782. [Google Scholar] [CrossRef]
- Pineda-Valdes, G.; Bullerman, L.B. Thermal stability of moniliformin at varying temperature, pH, and time in an aqueous environment. J. Food Prot. 2000, 63, 1598–1601. [Google Scholar] [CrossRef] [PubMed]



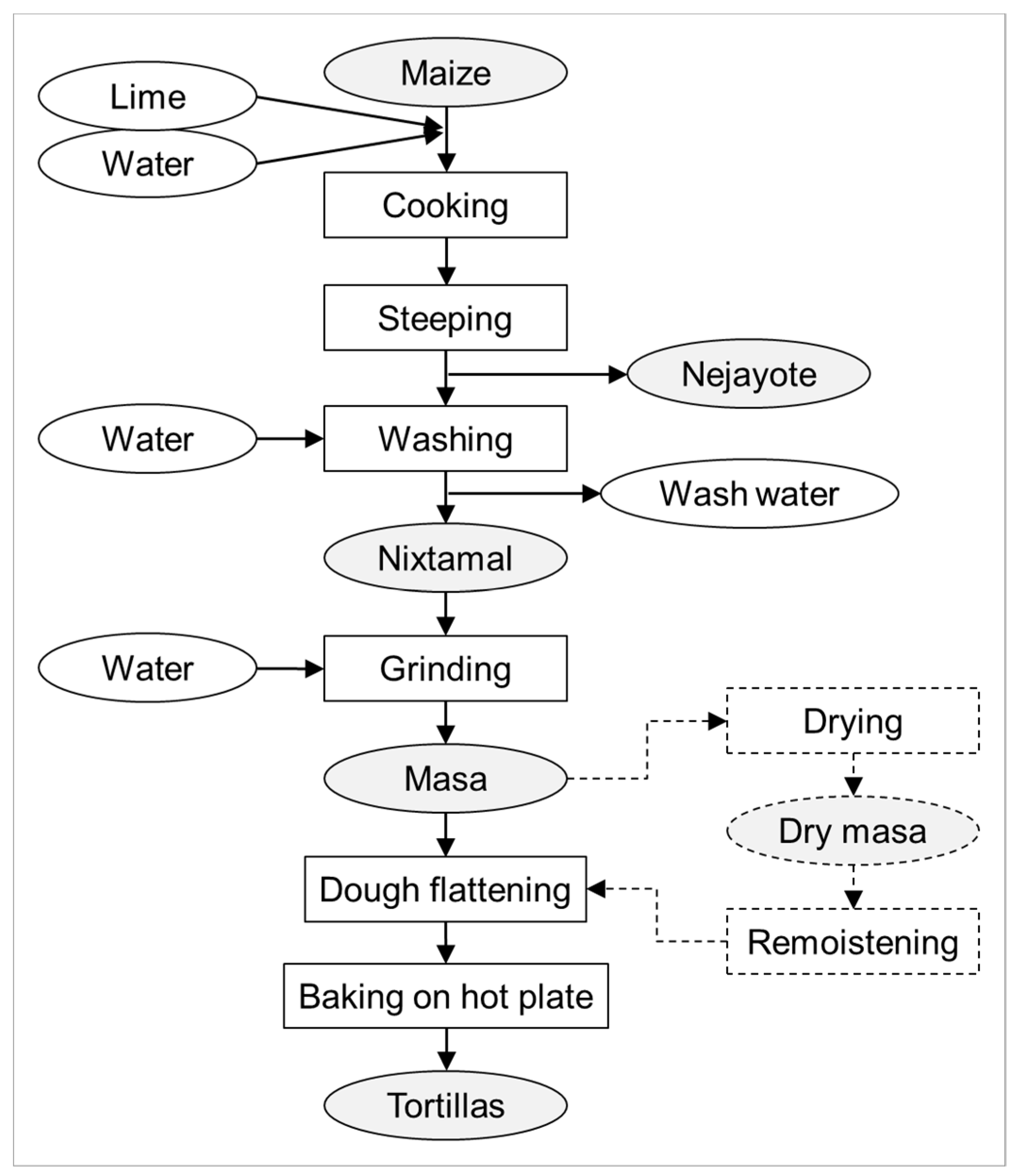{kind=link}
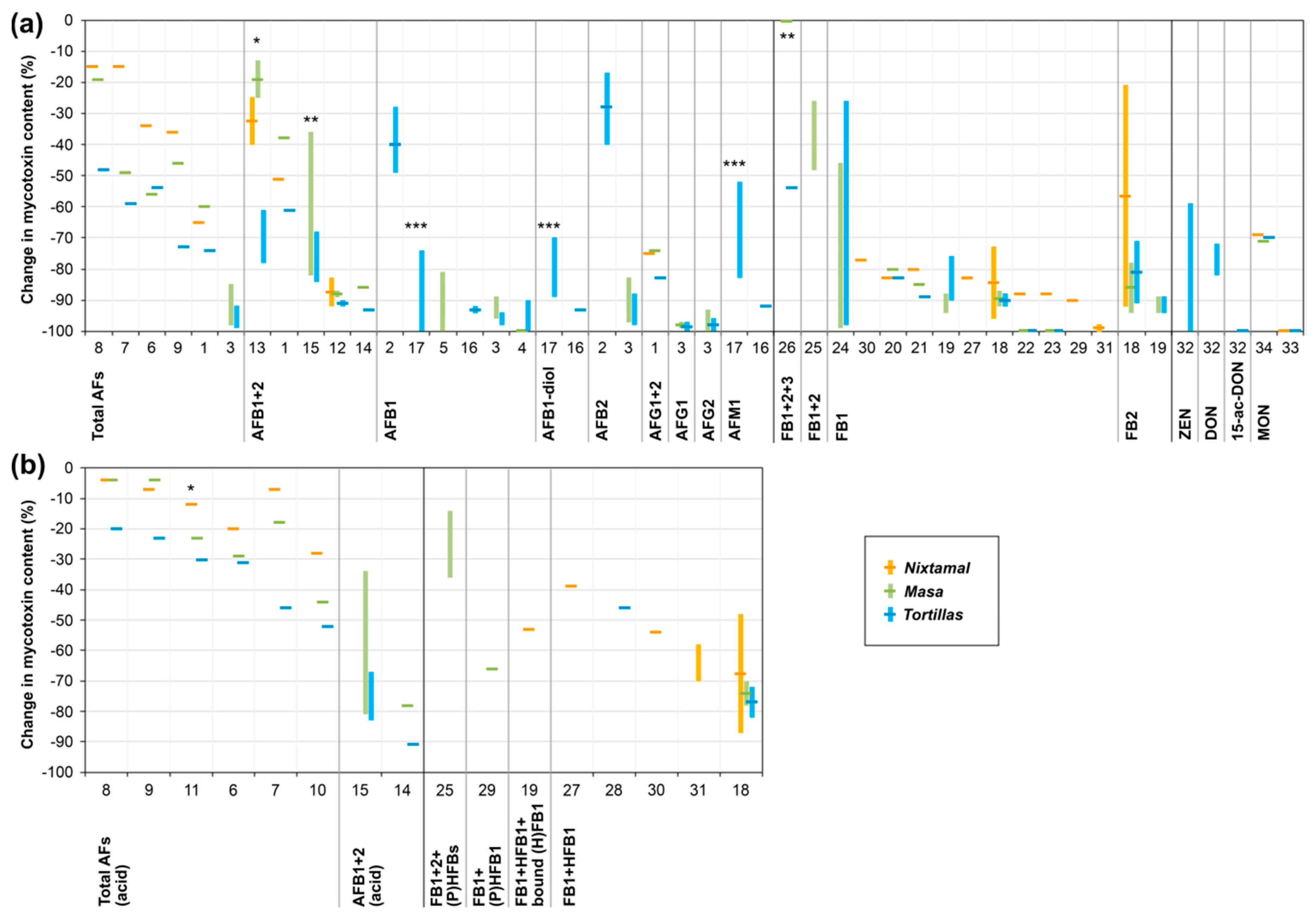{kind=link}
| Study No. in Figure 2 | Nixtamalization | Tortilla Baking on Hot Plate | Afla- toxin(s) | Initial Level in Raw Maize (µg/kg) a | Corrected Change (%) b | Comment(s) | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Alkaline Solution | Additions on Orig. Maize Mass Basis | Cooking | Steeping | Washing of Nixtamal | |||||||
| 1 | ca. 7.5% lime solution | 167% water, ca. 12.5% lime | 60 min | Only cool down after cooking | Not indicated | Performed (further details not provided) | Total AFs | Not provided (inoculated) | Nixtamal: ca. −65 c Nejayote: accum. Masa: ~−60 c Tortillas: ~−74 c | Overall: Relative quantification only and very approximate data | [21] |
| AFB1 + AFB2 | Not provided (inoculated) | Nixtamal: ~−51 c Nejayote: accum. Masa: ~−38 c Tortillas: ~−61 c | Most of the lost AFB was found in the nejayote | ||||||||
| AFG1 + AFG2 | Not provided (inoculated) | Nixtamal: ~−75 c Nejayote: n.d. Masa: ~−74 c Tortillas: ~−83 c | AFG1 + AFG2 were largely decomposed | ||||||||
| 2 | 2% Ca(OH)2 solution | Not specified | 5 min (incl. stirring) | 12 h | Thorough rinse with distilled water | 110–120 °C, 7–8 min on each side | AFB1 | 417–476 | Tortillas: −28 to −49 (mean: −40) | - | [22] |
| AFB2 | 54–59 | Tortillas: −17 to −40 (mean: −28) | - | ||||||||
| 3 | 0.6%–1.87% lime solution | 1480% water, 1–3% lime | 95 °C, 40 min or 121 °C, 0.34 bar, 30 min | O/N | Several washes with tap water | 180–250 °C (internal temp.: 94 °C), 1.5 min | Total AFs | 5245–60,478 (inoculated) | Masa: ~−85 to −98 Tortillas: ~−92 to −99 | No significant differences between different treatments | [20] |
| AFB1 | 3265–37,696 (inoculated) | Masa: ~−89 to −96 Tortillas: ~−94 to −98 | |||||||||
| AFB2 | 512–9513 (inoculated) | Masa: ~−83 to −97 Tortillas: ~−88 to −98 | |||||||||
| AFG1 | 1236–11,131 (inoculated) | Masa: ~−97 to −99 Tortillas: ~−97 to −100 | |||||||||
| AFG2 | 226–1639 (inoculated) | Masa: ~−93 to −100 Tortillas: ~−96 to −100 | |||||||||
| 4 | 0.33% lime solution | 300% water, 1% lime | 90 °C, 45 min | 24 °C, 18 h | 1 rinse with tap water | - | AFB1 | 0.68 | Masa: ca. −100 (<LOD) c | - | [24] |
| 150 °C, 5 min on each side | AFB1 | 125 (spiked to masa) | Tortillas: ca. −90 to −100 c | Range is due to different detection methods | |||||||
| 5 | Lime solution | 1% lime (water: not specified) | 94 °C, 50 min | 17 h | 2–3 washes | - | AFB1 | 37|251 | Masa: −100|−97 d | - | [25] |
| 3H-AFB1 | ~200 (spiked) | Masa: −81 to −84 d Nejayote: accum. Washing water: accum. | Range is due to different detection methods | ||||||||
| 6 | 0.33% Ca(OH)2 solution | 300% water, 1% Ca(OH)2 | 20 min | 15 h | Thorough rinse with water | 1 min on each side | AF | 135 | Nixtamal: ~−34/~−20 * Masa: ~−56/~−29 * Tortillas: ~−54/~−31 * | - | [23] |
| 7 | As described above | As described above | As described above | As described above | As described above | As described above | AF | 142 | Nixtamal: ~−15/~−7 * Masa: ~−49/~−18 * Tortillas: ~−59/~−46 * | Steeping was performed before cooking! | |
| - | As described above | As described above | - | As described above | As described above | As described above | AF | 145 | Nixtamal: ~−17/~−6 * Masa: ~−14/~−4 * Tortillas: ~−54/~−42 * | - | |
| 8 | 7.8% Ca(OH)2 solution | 160% water, 12.5% Ca(OH)2 | 60 min | Left to cool down for 1 h | As described above | As described above | AF | 142 | Nixtamal: ~−15/~−4 * Masa: ~−19/~−4 * Tortillas: ~−48/~−20 * | - | |
| 9 | 0.25% Ca(OH)2 solution | 300% water, 0.75% Ca(OH)2 | 75 min | 24 h | - | As described above | AF | 142 | Nixtamal: ~−36/~−7 * Masa: ~−46/~−4 * Tortillas: ~−73/~−23 * | - | |
| 10 | 0.33% lime solution | 300% water, 1% lime | 98 °C, 40 min | 14 h | 2 washes with 300% water (based on orig. maize mass) | Baking in a three-tiered, gas-fired oven for 39 s. Average temp. at the three levels: 177 °C, 233 °C, 453 °C | Total AFs | ca. 110 | Nixtamal: ~−28 * Nejayote: n.d. * Masa: ~−44 * Tortillas: −52 * Maize chips: −79 * Tortilla chips: −85 * | Both processes: Corn chips and tortilla chips were prepared by frying masa and tortilla strips in oil at 190 °C for 2 and 3 min, respectively | [36] |
| 11 | As described above | As described above | No cooking, but manual mixing of maize and lime with boiling water | As described above | As described above | As described above | Total AFs | ca. 43 | Nixtamal: ~−12 * Nejayote: accum. * Masa: ~−23 * Tortillas: −30 * Maize chips: −71 * Tortilla chips: −71 * | ||
| 12 | 1% lime solution | 200% distilled water, 2% lime | 85 °C, 70 min | 22 °C, 12 h | 1 wash with 200% tap water (based on orig. maize mass) | 270 °C, in total 50–54 s on each side | AFB1 + AFB2 | 29|93 (inoculated) | Nixtamal: −92|−83 c Masa: −89|−87 c Tortillas: −92|−90 c | ― | [35] |
| 13 | 0.375% lime solution | 80% distilled water, 0.3% lime | No cooking, but manual mixing of maize and hot lime water (92 °C) for 10 min | 22 °C, 2 h; no removal of water indicated | - | As described above | AFB1 + AFB2 | 29|93 (inoculated) | Nixtamal: −40|−25 c Masa: −25|−13 c Tortillas: −78|−61 c | Use of maize meal (particle size: 800 µm) | |
| 14 | 1% lime solution | 300% distilled water, 3% lime | 85 °C, 35 min | 22 °C, 14 h | 1 wash 200% with tap water (based on orig. maize mass) | 270 °C, in total 50–54 s on each side | AFB1 + AFB2 | 678/680 ** (inoculated) | Nejayote: n.d./slight accum. ** Masa: ~−86/~−78 ** Tortillas: −93/~−91 ** | Most aflatoxins appear to be degraded | [39] |
| 15 | 0.5% Ca(OH)2 solution | 100% water, 0.5% Ca(OH)2 | Microwave (1650 W, 2450 Hz), 5.5 min | 22 °C, 3 h; no water removal | - | 270 °C, in total 54–55 s on each side | AFB1 + AFB2 | 22–141 (inoculated) | Masa: −36 to −82/~−34 to −81 ** Tortillas: −68 to −84/~−67 to −83 ** | Use of maize grits. Higher reduction at higher initial concentration | [26] |
| 16 | 1% lime solution | 300% water, 3% lime | 90–96 °C, 30 min | O/N | Several rinses with tap water | 290 °C, in total 40–80 s on each side | AFB1 | 495|29 | Tortillas: −94|~−92 | - | [30] |
| AFB1-diol | 30|- | Tortillas: ~−93|- | |||||||||
| AFM1 | 402|- | Tortillas: −92|- | |||||||||
| - | Water, only | 75% water | Extruder (low shear, single- screw, 35 rpm screw speed, 87 °C barrel temp.) | - | - | As described above | AFB1 | 495|29 | Tortillas: ~−46|~−68 | All extrusion treatments: Use of maize meal (particle size: 800 µm) | |
| AFB1-diol | 30|- | Tortillas: −54|- | |||||||||
| AFM1 | 402|- | Tortillas: −20|- | |||||||||
| 17 | 0.4%–0.67% lime solution | 75% water, 0.3%–0.5% lime | Extruder (for details see above) | - | - | As described above | AFB1 | 495|29 | Tortillas: −74 to −85|~−100 | Higher reduction at higher lime concentration | |
| AFB1-diol | 30|- | Tortillas: ~−70 to −89|- | |||||||||
| AFM1 | 402|- | Tortillas: ~−52 to −83|- | |||||||||
| - | 0.4% lime, 1%–4% H2O2 solution | 75% water, 0.3% lime, 0.75%–3% H2O2 | Extruder (for details see above) | - | - | As described above | AFB1 | 495|29 | Tortillas: ~−67 to −78|~−100 | Higher reduction at higher H2O2 concentration. At 0.3% lime + 3% H2O2: affection of taste | |
| AFB1-diol | 30|- | Tortillas: ~−68 to −84|- | |||||||||
| AFM1 | 402|- | Tortillas: ~−69 to −81|- | |||||||||
| Study No. in Figure 2 | Nixtamalization | Tortilla Baking on Hot Plate | Fumo- nisin(s) | Initial level in Raw Maize (µg/kg) a | Corrected Change (%) b | Comment(s) | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Alkaline Solution | Additions on Orig. Maize Mass Basis | Cooking | Steeping | Washing of Nixtamal | |||||||
| 18 | ~0.37% lime solution | ~1290% water, ~4.8% lime | 100 °C, 5 min in a steam kettle (in a perforated nylon bag) | 15 h | Wash with water | Baking in a gas-fired oven with three moving tiers (further details not provided) | FB1 | 8790 | Nixtamal, unwashed: ~−76|~−97 Nixtamal, washed: ~−73|~−96 Nejayote: n.d.|slight accum. Washing water: n.d.|slight accum. Masa: ~−87|~−92 Tortillas: ~−88|~−92 Tortilla chips: n.a.|~−94 | Overall: Data of two production runs are given; tortilla chips were produced in only one run by frying in oil at 190 °C for 60 s. Of the initial FB1, a total of 62%–90% was recovered as FB1 or HFB1 e from the nejayote, mostly as HFB1 | [46] |
| HFB1 | Probably n.d. | (Slight) accum. in all intermediate products and (by)products, highest accum. in nejayote | |||||||||
| FB1 + HFB1 | ~12,000 nmol/kg | Nixtamal, unwashed: ~−48|~−85 f Nixtamal, washed: ~−48|~−87 f Masa: ~−70|~−78 f Tortillas: ~−72|~−82 f Tortilla chips: n.a.|~−86 f | |||||||||
| FB2 | 1,970 | Nixtamal, unwashed: ~−32|~−97 Nixtamal, washed: ~−21|~−92 Nejayote: n.d.|n.d. Washing water: n.d.|n.d. Masa: ~−78|~−94 Tortillas: ~−71|~−91 Tortilla chips: n.a.|~−90 | - | ||||||||
| 19 | Processing in a pilot plant according to commercial procedures including alkaline cooking, steeping, and washing of maize kernels, as well as baking and deep frying of masa to produce tortilla chips (details not provided) | FB1 | 220–46,500 | Nejayote: slight accum. Masa: −88 to −94 d Baked tortilla chips: −76 to −90 d Fried tortilla chips: −36 to −78 d | Ca. 34% and 45% of the initial FB1 f was detected as FB1, PHFB1, or HFB1 in the masa and nejayote, respectively. No indications for significant fumonisin–sugar adduct formation/accumulation | [45] | |||||
| PHFB1 | n.d.−1340 | Nejayote: n.d. or slight accum. Masa: ~−33 d to slight accum. | |||||||||
| HFB1 | n.d.−950 | Nejayote: accum. Masa: no change to slight accum. | |||||||||
| FB2 | (Not provided) | Masa: ca. −89 to −94 d Baked tortilla chips: ca. −89 to −94 d | - | ||||||||
| - | - | - | - | - | - | Baking in a tortilla oven at 260 °C, 20 s | FB1 + FB2 | 100–281 | Tortilla chips: ‘as is’: −32 to −78 (mean: −59) | Use of maize flour mixtures. Commercial processing. Tortilla chips were prepared by frying tortilla strips at 170–175 °C for 40 s | [49] |
| 20 | ~1% lime solution | ~77% water, ~0.8% lime | 7 min | 18 h | No information | 250 °C | FB1 | 1001 | Cooked maize: −56 c Nixtamal: −83 c Masa: −80 c Tortillas: −83 c | Commercial processing. Cooked maize was sampled before steeping | [50] |
| 21 | ~0.7% lime solution | ~200% water, ~1.5% lime | 150 min | 16 h | No information | 250 °C | FB1 | 681 | Cooked maize: −73 c Nixtamal: −80 c Masa: −85 c Tortillas: −89 c | ||
| 22 | ~0.56% lime solution | ~160% water, ~0.9% lime | 10 min | 16 h | No information | 375 °C | FB1 | 1441 | Cooked maize: −83 c Nixtamal: −88 c Masa: −100 c Tortillas: −100 c | ||
| 23 | ~1.1% lime solution | ~70% water, ~0.8% lime | 120 min | 18 h | No information | 232 °C | FB1 | 1653 | Cooked maize: −78 c Nixtamal: −88 c Masa: −100 c Tortillas: −100 c | ||
| 24 | ~0.25% lime solution | 200% water; ~0.5% lime | 60 min | 18 h | Rinse in 430% water (based on orig. maize mass) | Baking over an iron grill at 190–200 °C for ca. 4 min | FB1 | 150–11,800 (inoculated) | Masa: −46 to −99 c Tortillas: −26 to −98 c | Higher reduction at higher initial FB1 level | |
| - | ~0.25–1.6% lime solution | 200% water; ~0.5–3.2% lime | 15 or 60 min | 18 h | As described above | As described above | FB1 | 150–11,800 (inoculated) | Masa/Tortillas: up to −100 c | Higher reduction at higher lime concentration (and at higher initial FB1 level) | |
| 25 | 0.33 or 1.67% lime solution | 300% distilled water, 1% or 5% lime | 90 °C, 15–60 min | 17 h | 2 rinses with 200% tap water (based on orig. maize mass) | - | FB1 + FB2 | 6,480–8,930 | Masa: −26 to −48 c | Of the initial FB1 + FB2f, a total of 64%–86% retained as parent form, PHFB, or HFB in the masa | [51] |
| PHFB1 + PHFB2 | 110–260 | Masa: no change to accum. | Increase at higher lime concentration | ||||||||
| HFB1 + HFB2 | n.d. | Masa: accum. | Higher accum. at higher lime concentration | ||||||||
| - | Distilled water, only | 300% distilled water | As described above | As described above | As described above | - | FB1 + FB2 | 5230–20,380 | Maize dough: −45 to −78 c | Here, a total of 21%–55% of the initial FB1 + FB2f retained as parent form, PHFB, or HFB in the maize dough | |
| PHFB1 + PHFB2 | 140–510 | Maize dough: −29 to −75 c | |||||||||
| HFB1 + HFB2 | n.d. | Maize dough: n.d. | |||||||||
| 26 | 0.5% Ca(OH)2 solution | 100% water, 0.5% Ca(OH)2 | Microwave (1650 W, 2450 Hz), 3.75 min | 25 °C, 3.5 h; no removal of water indicated | - | 270 °C, in total 54–55 s on each side | FB1 + FB2 + FB3 | 2137 (inoculated) | Masa: −6 (n.s.) c Tortillas: −54 c | Use of maize grits | [48] |
| 27 | 1.2% lime solution | Not specified | 95–100 °C, 55 min | 14 h | Wash with ca. 300% tap water | - | FB1 | 239,000 (inoculated) | Nixtamal: −83 c Nejayote: slight accum. | Pericarp removal during washing was avoided | [57] |
| HFB1 | n.d. | Nixtamal: accum. Nejayote: accum. | |||||||||
| FB1 + HFB1e | 239,000 | Nixtamal: −39 c Nejayote: accum. | |||||||||
| 28 | ~1.3% lime solution | 325% water, ~4.1% lime | ca. 105 min | 15 h | 3 rinses with 275% water (based on orig. maize mass) | 170–212 °C, ca. 3.5 min | FB1 + HFB1 e | ca. 38,100 | Tortillas: −46 f | - | [47] |
| 29 | 1.2% Ca(OH)2 solution | 750% water, 9% lime | 90–100 °C, 60 min | O/N | 3 rinses with 750% water (based on orig. maize mass) | - | FB1 | ~23,314 | Nixtamal: ~−90 c | Of the initial FB1 f, a total of ~47% retained as FB1, HFB1, or bound (H)FB1 in the nixtamal | [56] |
| HFB1 | ~329 | Nixtamal: ca. +1400 c | |||||||||
| Protein- bound (H)FB1 | ~69 (recovered as HFB1) | Nixtamal: ~+476 c | |||||||||
| Total bound (H)FB1 | ~89 (recovered as HFB1) | Nixtamal: ~+673 c | |||||||||
| - | Water, only | 750% water | As described above | As described above | As described above | - | FB1 | ~23,314 | ‘Mock-nixtamal’: ~−53 (n.s.) c | Here, a total of ~44% of the initial FB1f retained as FB1, HFB1, or bound (H)FB1 in the ‘mock-nixtamal’ | |
| HFB1 | ~329 | ‘Mock-nixtamal’: ~−60 (n.s.) c | |||||||||
| Protein- bound (H)FB1 | ~69 (recovered as HFB1) | ‘Mock-nixtamal’: no change c | |||||||||
| Total bound (H)FB1 | ~89 (recovered as HFB1) | ‘Mock-nixtamal’: ~+9 (n.s.) c | |||||||||
| 30 | 1.2% Ca(OH)2 solution | 1,200% water, 14.4% lime | 90–100 °C, 60 min | O/N | 3 washes with 1200% distilled water (based on orig. maize mass) | - | FB1 | 9080 (inoculated) | Nixtamal: −77 | Raw material: ground maize used as fungal growth medium. Nixtamal was prepared for a feeding trial. Concentrations and changes are given for the mixed diet | [62] |
| HFB1 | 250 | Nixtamal: ~+408 | |||||||||
| FB1 + HFB1 | 13,200 nmol/kg | Nixtamal: ~−54 f | |||||||||
| - | Water, only | 1,200% water | As described above | As described above | As described above | - | FB1 | 9080 (inoculated) | ‘Mock-nixtamal’: −87 | ||
| HFB1 | 250 | ‘Mock-nixtamal’: ~+120 | |||||||||
| FB1 + HFB1 | 13,200 nmol/kg | ‘Mock-nixtamal’: ~−77 f | |||||||||
| 31 | 1.2% Ca(OH)2 solution | 300% water, 3.6% lime | 80–100 °C, 60 min | O/N | One wash with ca. 300% water (based on orig. maize mass) | - | FB1 | 45,200–48,000 (inoculated) | Nixtamal: ~−98 to −100 | Nixtamal was prepared for a feeding trial. Concentrations and changes are given for the mixed diet | [64] |
| HFB1 | n.d. | Nixtamal: accum. | |||||||||
| FB1 + HFB1 | ~62,600–66,500 nmol/kg | Nixtamal: ~−58 to −70 f | |||||||||
| Study No. in Figure 2 | Nixtamalization | Tortilla Baking on Hot Plate | Myco- toxin(s) | Initial Level in Raw Maize (µg/kg) a | Corrected Change (%) b | Comment(s) | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Alkaline Solution | Additions on Orig. Maize Mass Basis | Cooking | Steeping | Washing of Nixtamal | |||||||
| 32 | 2% Ca(OH)2 solution | Not specified | 5 min (incl. stirring) | 12 h | Thorough rinse with distilled water | 110–120 °C, 7–8 min on each side | ZEN | 230|4,230 | Nejayote: n.d.|weak accum. Tortillas: −100|−59 | - | [65] |
| DON | 3,280|12,260 | Nejayote: n.d.|n.d. Tortillas: −82|−72 | |||||||||
| 15-acetyl-DON | 1,490|9,830 | Nejayote: n.d.|n.d. Tortillas: −100|−100 | |||||||||
| ZEN | 750|3,620 (spiked) | Nejayote: n.d.|weak accum. Tortillas: −71|−74 | Mycotoxins were injected into the maize embryos | ||||||||
| DON | 850|4460|8250 (spiked) | Nejayote: n.d.|n.d.|n.d. Tortillas: −82|−72|–74 | |||||||||
| - | - | - | - | - | - | Baking in a tortilla oven at 260 °C, 20 s | ZEN | 4.5–8.7|19–24 | Tortilla chips: ‘as is’: +116 to −7 (mean: +32)|−35 to −64 (mean: −49) | Use of maize flour mixtures. Commercial processing. Tortilla chips were prepared by frying tortilla | [49] |
| DON | 47–466 | Tortilla chips: ‘as is’: +28 to −76 (mean: −32) | |||||||||
| 33 | 0.25% lime solution | 400% water, 1% lime | 88 °C, 20 min | 16 h | Two rinses with 250% water (based on orig. maize mass) | Baking in a gas-fired oven at ca. 365 °C for ca. 3 min | MON | 1420 | Cooked maize: −97 c Nixtamal: −100 c Nejayote: n.d. Wash water: n.d. Masa: −100 c Tortillas: −100 c | Pilot-scale process | [68] |
| 34 | 0.25% lime solution | 400% water, 1% lime | 88 °C, 20 min | 16 h | Two rinses with 125% water (based on orig. maize mass) | ca. 250 °C, 3 min on each side | MON | 17,640 (inoculated) | Cooked maize: −54 c Nixtamal (before washing): −64 c Nixtamal (washed): ~−69 c Nejayote: n.d. Wash water: n.d. Masa: ~−71 c Tortillas: ~−70 c | Laboratory-scale | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schaarschmidt, S.; Fauhl-Hassek, C. Mycotoxins during the Processes of Nixtamalization and Tortilla Production. Toxins 2019, 11, 227. https://doi.org/10.3390/toxins11040227
Schaarschmidt S, Fauhl-Hassek C. Mycotoxins during the Processes of Nixtamalization and Tortilla Production. Toxins. 2019; 11(4):227. https://doi.org/10.3390/toxins11040227
Chicago/Turabian StyleSchaarschmidt, Sara, and Carsten Fauhl-Hassek. 2019. "Mycotoxins during the Processes of Nixtamalization and Tortilla Production" Toxins 11, no. 4: 227. https://doi.org/10.3390/toxins11040227
APA StyleSchaarschmidt, S., & Fauhl-Hassek, C. (2019). Mycotoxins during the Processes of Nixtamalization and Tortilla Production. Toxins, 11(4), 227. https://doi.org/10.3390/toxins11040227