Investigation of the Metabolic Profile and Toxigenic Variability of Fungal Species Occurring in Fermented Foods and Beverage from Nigeria and South Africa Using UPLC-MS/MS

 , , ,
, , ,  , ,
, ,  and
and 
Abstract
1. Introduction
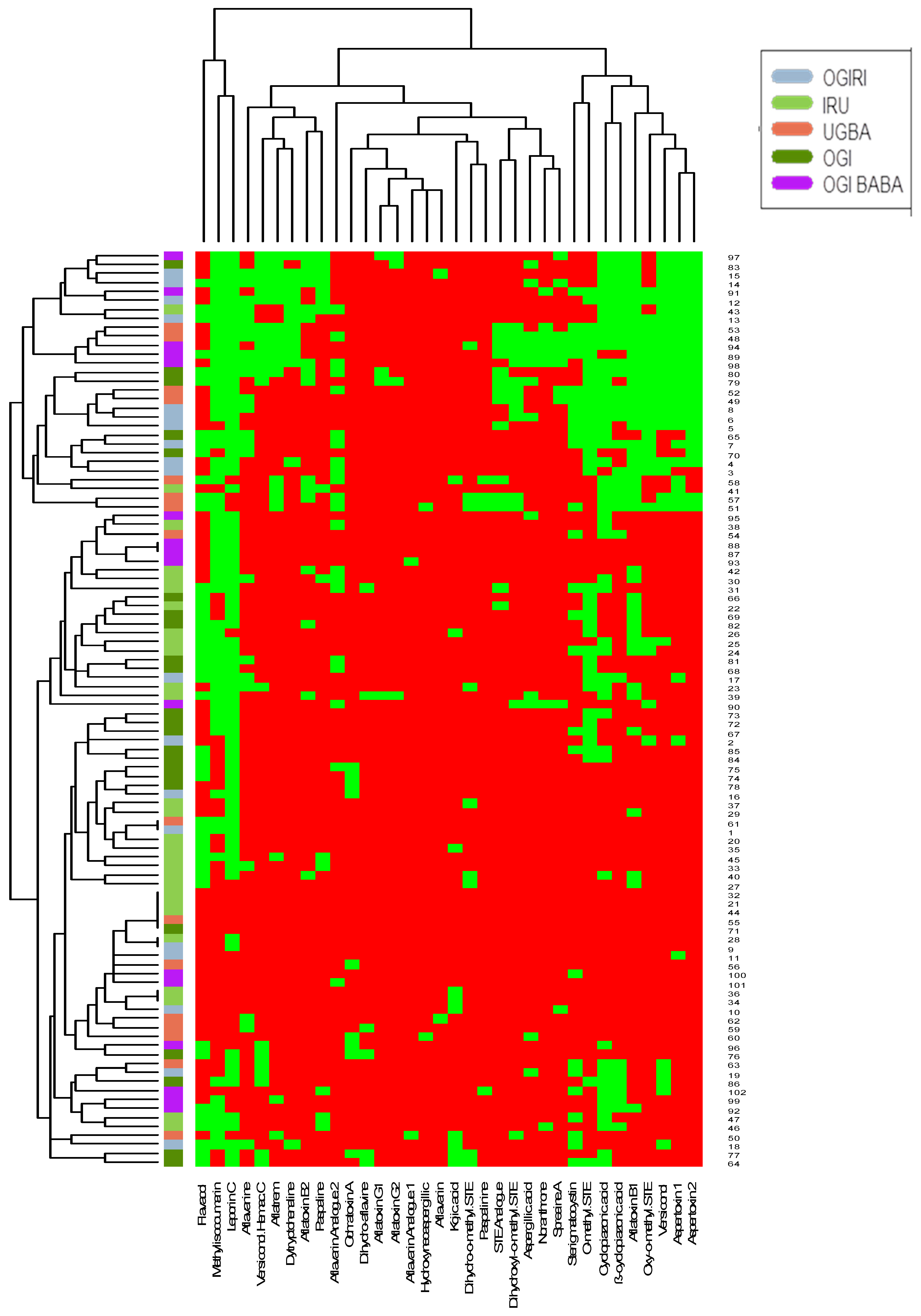
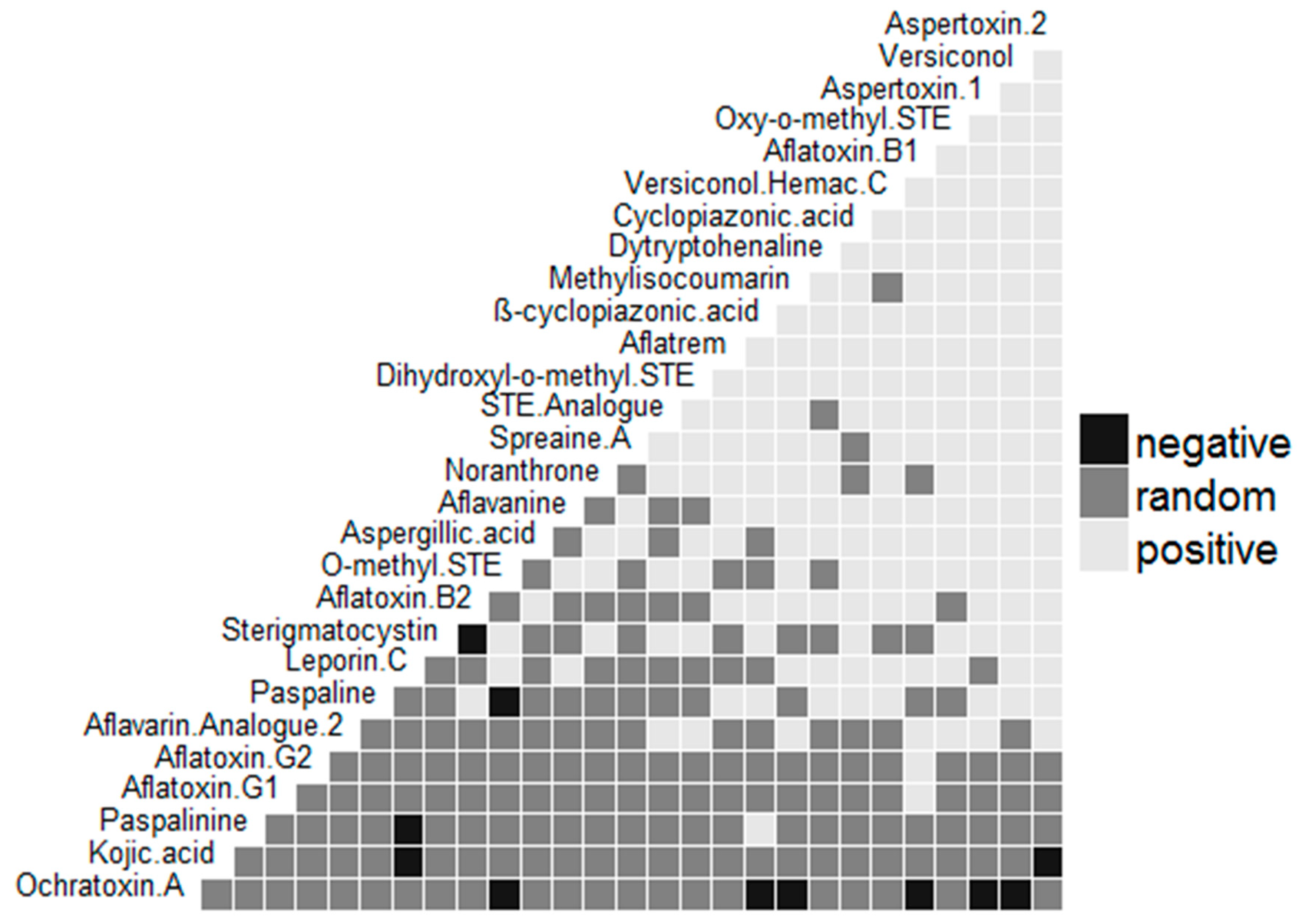
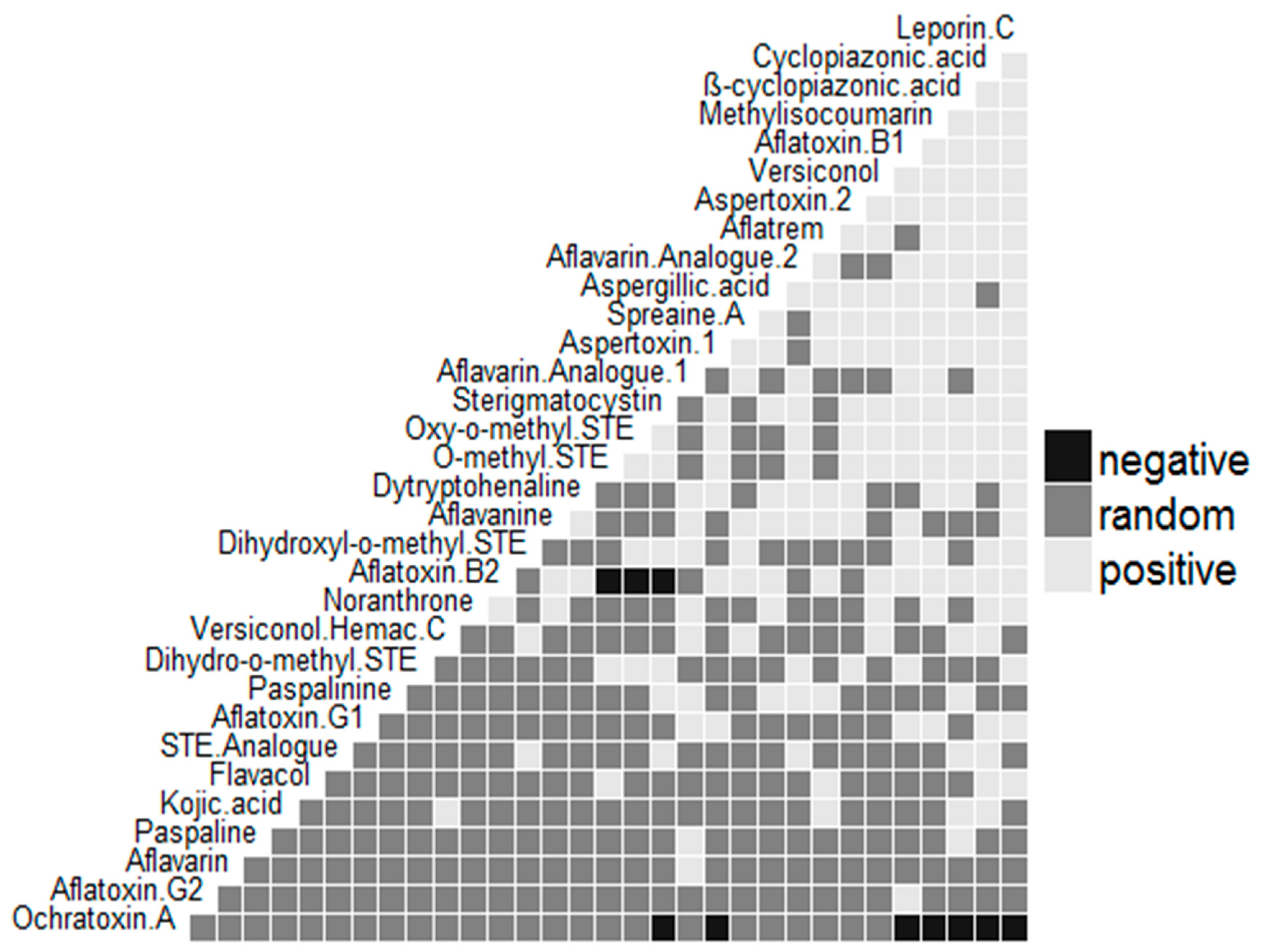
2. Results and Discussion
3. Conclusions
4. Materials and Methods
4.1. Reagents and Standards
4.2. Standards
4.3. Origin of Fermented Food Samples
4.4. Sample Preparation
4.5. Multi-Mycotoxin Extraction
4.6. Instrumentation Identification and Quantification of Aspergillus Metabolites
4.7. Detection and Quantification of Penicillium and Fusarium Metabolites
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Osbourn, A. Secondary metabolic gene clusters: Evolutionary toolkits for chemical innovation. Trends Genet. 2010, 26, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Jayani, R.S.; Saxena, S.; Gupta, R. Microbial pectinolytic enzymes: A review. Process Biochem. 2005, 10, 2931–2944. [Google Scholar] [CrossRef]
- Adekoya, I.; Obadina, A.; Phoku, J.; Nwinyi, O.; Njobeh, P. Contamination of fermented foods in Nigeria with fungi. LWT Food Sci. Technol. 2017, 86, 76–84. [Google Scholar] [CrossRef]
- Adekoya, I.O.; Obadina, A.O.; Chilaka, A.C.; De Boevre, M.; Okoth, S.; De Saeger, S.; Njobeh, P.B. Mycobiota and co-occurrence of mycotoxins in South African maize-based opaque beer. Int. J. Food Microbiol. 2018, 270, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Adekoya, I.O.; Obadina, A.O.; Phoku, J.Z.; De Boevre, M.; De Saeger, S.; Njobeh, P.B. Fungal and mycotoxin contamination of fermented foods from selected South African markets. Food Control 2018, 90, 295–303. [Google Scholar] [CrossRef]
- Adekoya, I.; Njobeh, P.; Obadina, A.; Chilaka, C.; Okoth, S.; De Boevre, M.; De Saeger, S. Awareness and prevalence of mycotoxin contamination in selected Nigerian fermented foods. Toxins 2017, 9, 363. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Varga, J. Aspergillus systematics in the genomic era. Stud. Mycol. 2007, 59, 1–206. [Google Scholar] [CrossRef]
- IARC (International Agency for Research on Cancer). Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. IARC Monogr. Eval. Carcinog. Risks to Hum. 2002, 96, 1–390. [Google Scholar]
- Jiang, H.L.; Liu, X.Y.; Qiu, Y.X.; Yao, D.S.; Xie, C.F.; Liu, D.L. Development of an aptasensor for the fast detection of versicolorin A. Food Control 2015, 56, 202–210. [Google Scholar] [CrossRef]
- Yu, J.; Bhatnagar, D.; Cleveland, T.E. Completed sequence of aflatoxin pathway gene cluster in Aspergillus parasiticus. FEBS Lett. 2004, 564, 126–130. [Google Scholar] [CrossRef]
- Yazar, S.; Omurtag, G.Z. Fumonisins, trichothecenes and zearalenone in cereals. Int. J. Mol. Sci. 2008, 9, 2062–2090. [Google Scholar] [CrossRef] [PubMed]
- Phoku, J.Z.; Barnard, T.G.; Potgieter, N.; Dutton, M.F. Mycotoxigenic potentials of the genera: Aspergillus, Fusarium and Penicillium isolated from houseflies (Musa domestica L.). Acta Trop. 2017, 168, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Karlovsky, P.; Suman, M.; Berthiller, F.; De Meester, J.; Eisenbrand, G.; Perrin, I.; Oswals, I.; Speijers, G.; Chiodini, A.; Recker, T.; et al. Impact of food processing and detoxification treatments on mycotoxin contamination. Mycotoxin Res. 2016, 32, 179–205. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Tan, Y.; Wang, S.; Gardiner, D.M.; De Saeger, S.; Liao, Y.; Wang, C.; Fan, Y.; Wang, Z.; Wu, A. Mycotoxigenic potentials of Fusarium species in various culture matrices revealed by mycotoxin profiling. Toxins 2017, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Lamboni, Y.; Nielsen, K.F.; Linnemann, A.R.; Gezgin, Y.K.; Hell, K.; Nout, M.J.R.; Smid, E.J.; Tamo, M.; Van Boekel, M.A.; Hoof, J.B.; et al. Diversity in secondary metabolites including mycotoxins from strains of Aspergillus section Nigri isolated from raw cashew nuts from Benin, West Africa. PLoS ONE 2016, 11, e0164310. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.F.; Mogensen, J.M.; Johansen, M.; Larsen, T.O.; Frisvad, J.C. Review of secondary metabolites and mycotoxins from the Aspergillus niger group. Anal. Bioanal. Chem. 2009, 395, 1225–1242. [Google Scholar] [CrossRef] [PubMed]
- Van Egmond, H.P.; Schothorst, R.C.; Jonker, M.A. Regulations relating to mycotoxins in food: Perspectives in a global and European context. Anal. Bioanal. Chem. 2007, 389, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Aldars-Garcia, L.; Marin, S.; Sanchis, V.; Magan, N.; Medina, A. Assessment of intraspecies variability in fungal growth initiation of Aspergillus flavus and aflatoxin B1 production under static and changing temperature levels using different initial conidial inoculum levels. Int. J. Microbiol. 2018, 271, 1–11. [Google Scholar] [CrossRef]
- Larsen, T.O.; Smedsgaard, J.; Nielsen, K.F.; Hansen, M.E.; Frisvad, J.C. Phenotypic taxonomy and metabolite profiling in microbial drug. Nat. Prod. Rep. 2005, 22, 672–695. [Google Scholar] [CrossRef]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.B.; Hubka, V.; Klaassen, C.H.W.; Perrones, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef]
- Rodrigues, P.; Santos, C.; Venancio, A.; Lima, N. Species identification of Aspergillus section Flavi isolates from Portuguese almonds using phenotypic, including MALDI-TOF ICMS, and molecular approaches. J. Appl. Microbiol. 2011, 111, 877–892. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.W.; Geiser, D.M.; Burt, A.; Koufopanou, V. The evolutionary biology and population genetics underlying fungal strain typing. Clin. Microbiol. Rev. 1999, 12, 126. [Google Scholar] [CrossRef] [PubMed]
- Okoth, S.; De Boevre, M.; Corominas, A.V.; Di Mavungu, J.; Landschoot, S.; Kyalo, M.; Njuguna, J.; Jagger, H.; De Saeger, S. Genetic and toxigenic variability within Aspergillus flavus population isolated from maize in two diverse environments in Kenya. Front. Microbiol. 2018, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Due, M.; Frisvad, J.C.; Samson, R.A. Taxonomic revision of Aspergillus section Clavati based on molecular, morphological and physiological data. Stud. Mycol. 2007, 59, 89–106. [Google Scholar] [CrossRef] [PubMed]
- Vaamonde, G.; Patriarca, A.; Fernandez, V.; Comerio, R.; Degrossi, C. Variability of aflatoxin and cyclopiazonic acid production by Aspergillus section Flavi from different substrates in Argentina. Int. J. Food Microbiol. 2003, 88, 79–84. [Google Scholar] [CrossRef]
- Yabe, K.; Nakajima, H. Enzyme reactions and genes in aflatoxin biosynthesis. App. Microbiol. Biotechnol. 2004, 64, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Pildain, M.B.; Frisvad, J.C.; Vaamonde, G.; Cabral, D.; Varga, J.; Samson, R.A. Two novel aflatoxin-producing Aspergillus species from Argentinean peanuts. Int. J. Syst. Evol. Microbiol. 2008, 58, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Zorzete, P.; Baquiao, A.C.; Atayde, D.D.; Reis, T.A.; Goncalez, E.; Correa, B. Mycobiota, aflatoxins and cyclopiazonic acid in stored peanut cultivars. Food Res. Int. 2013, 52, 380–386. [Google Scholar] [CrossRef]
- Mogensen, J.M.; Frisvad, J.C.; Thrane, U.; Nielsen, K.F. Production of fumonisin B2 and B4 by Aspergillus niger on grapes and raisins. J. Agric. Food Chem. 2010, 58, 954–958. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Calado, T.; Venancio, A. Incidence of fumonisin B2 production by Aspergillus niger in Portuguese wine regions. J. Agric. Food Chem. 2011, 13, 7514–7518. [Google Scholar] [CrossRef]
- Storari, M.; Bigler, L.; Gessler, C.; Broggini, G.A.L. Assessment of the ochratoxin A production ability of Aspergillus tubingensis. Food Addit. Contam. 2012, 29, 1450–1454. [Google Scholar] [CrossRef] [PubMed]
- Versilovskis, A.; De Saeger, S. Sterigmatocystin: Occurrence in foodstuffs and analytical methods—An overview. Mol. Nutr. Food Res. 2010, 54, 136–147. [Google Scholar] [CrossRef]
- Logrieco, A.; Mule, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium species and mycotoxins associated with maize ear rot in Europe. Euro. J. Plant Pathol. 2002, 108, 597–609. [Google Scholar] [CrossRef]
- Wulff, E.G.; Sorensen, J.L.; Lubeck, M.; Nielsen, K.F.; Thrane, U.; Torp, J. Fusarium spp. associated with rice bakanae: Ecology, genetic diversity, pathogenicity and toxigenicity. Environ. Microbiol. 2010, 12, 649–657. [Google Scholar]
- Molina, M.; Gianuzzi, L. Modelling of aflatoxin production by Aspergillus parasiticus in a solid medium at different temperatures, pH and propionic acid concentrations. Food Res. Int. 2002, 35, 585–594. [Google Scholar] [CrossRef]
- Foroud, N.A.; Eudes, F. Trichothecenes in cereal grains. Int. Mol. Sci. 2009, 10, 147–173. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Larsen, T.O.; De Vries, R.; Meijer, M.; Houbraken, J.; Cabanes, F.J.; Ehrlich, K.; Samson, R.A. Secondary metabolite profiling, growth profiles and other tools for species recognition and important Aspergillus mycotoxins. Stud. Mycol. 2007, 59, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Okoth, S.; Nyongesa, B.; Ayugi, V.; Kan’gethe, E.; Korhonen, H.; Joutsjoki, V. Toxigenic potential of Aspergillus species occurring on maize kernels from two Agro-Ecological zones in Kenya. Toxins 2012, 4, 991–1007. [Google Scholar] [CrossRef]
- Bankole, S.A.; Adebanjo, A. Mycotoxins in food in West Africa: Current situation and possibilities of controlling it. J. Biotechnol. 2003, 2, 254–263. [Google Scholar]
- Olotu, I.O. Safety Assessment of Traditionally Fermented Food Produced in Nigeria and South Africa. Ph.D. Thesis, University of Johannesburg, Johannesburg, South Africa, June 2018. Available online: https://ujcontent.uj.ac.za/vital/access/manager/Repository/uj:28710 (accessed on 12 January 2019).
- European Commission (EC). Commission Regulation (EC) No 401/2006 of 23 February 2006 laying down the methods of sampling and analysis for the official control of the levels of mycotoxins in foodstuffs. Off. J. Eur. Union 2006, L70, 12–32. [Google Scholar]
- European Commission (EC). Commission Decision 2002/657/EC implementing Council Directive 96/23/EC concerning the performance of analytical methods and the interpretation of results. Off. J. Eur. Union 2002, L221, 8–36. [Google Scholar]
- Ediage, E.N.; Di Mavungu, J.D.; Monbaliu, S.; Van Peteghem, C.; De Saeger, S. A validated multi-analyte LC-MS/MS method for quantification of 25 mycotoxins in cassava flour, peanut cake and maize samples. J. Agric. Food Chem. 2011, 59, 5173–5180. [Google Scholar] [CrossRef] [PubMed]
- De Boevre, M.; Di Mavungu, J.D.; Landschoot, S.; Audenaert, K.; Eeckhout, M.; Maene, P.; De Saeger, S. Natural occurrence of mycotoxins and their masked forms in food and feed products. World Mycotoxin J. 2012, 5, 207–219. [Google Scholar] [CrossRef]
- Veech, J.A. A probabilistic model for analysing species co-occurrence. Glob. Ecol. Biogeogr. 2013, 22, 252–260. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolated Species | Accession No. | Ogi | Ogi baba | Umqombothi | Mahewu | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of Iso. Spp. (%) | No. of Tox. Spp. | Toxin Produced (Range: µg/kg) | No. of Iso. Spp. (%) | No. of Tox. Spp. | Toxin Produced (Range: µg/kg) | No. of Iso. Spp. (%) | No. of Tox. Spp. | Toxin Produced (Range: µg/kg) | No. of Iso. Spp. (%) | No. of Tox. Spp. | Toxin Produced (Range: µg/kg) | ||
| Aspergillus Species | |||||||||||||
| A. amstelodami | AY373885.1 | 1 (2) | 1 | STE (371) | - | - | - | - | - | - | - | - | - |
| A. clavatus | KU052566 | 2 (5) | - | - | 2 (13) | - | - | - | - | - | 3 (14) | - | - |
| A. flavus | KR611584 | 12 (29) | 7, 3 | AFB1 (109–231), STE (434–600) | 6 (38) | 4, 3 | AFB1 (217–1556), STE (49–701) | 6 (26) | 6, 3 | AFB1 (127–1117), STE (4–32) | 7 (32) | 4, 1 | AFB1 (69–1931), STE (119) |
| A. fumigatus | KX215145.1 | 4 (10) | - | - | - | - | - | 2 (9) | - | - | 3 (14) | - | - |
| A. minisclerotigenes | JF412778 | - | - | - | 1 (6) | - | - | 2 (9) | 1 | AFB1 (96) AFB2 (106) | - | - | - |
| A. niger | KX215115.1 | 7 (17) | 7 | OTA (197–1302) | 1 (6) | 1 | OTA (411) | 3 (13) | 2 | OTA (28–55) | 3 (14) | 2 | OTA (89–212) |
| A. parasiticus | DQ467988.1 | 7 (17) | 5, 5, 4, 3 | AFB1 (89–1018), AFB2 (118–324), AFG1 (95–158), AFG2 (978–664) | 2 (13) | 2, 2, 1, 1 | AFB1 (721–1030), AFB2 (185–498), AFG1 (48), AFG2 (34) | 3 (13) | 2, 2, 1 | AFB1 (172–3602), AFB2 (288–566), AFG1 (322) | 3 (14) | 2, 2, 1 | AFB1 (109–117), AFB2 (58–192), AFG1 (109) |
| A. ruber | KX215127.1 | 1 (2) | - | - | - | - | - | - | - | - | - | - | - |
| A. spp | KX215117.1 | - | - | - | 1 (13) | - | - | - | - | - | - | - | - |
| A. sclerotiorum | KT717312 | - | - | - | - | - | - | 2 (9) | - | - | - | - | - |
| A. sydowii | KR611596 | 1 (2) | 1 | STE (91) | 1 (13) | 1 | STE (433) | 2 (9) | 1 | STE (53) | - | - | - |
| A. tritici | KP780810 | 2 (5) | - | - | 1 (13) | - | - | 2 (9) | - | - | - | - | - |
| A. tubingensis | KT717311 | - | - | - | - | - | - | - | - | - | 1 (5) | - | - |
| A. ustus | HQ607918.1 | - | - | - | - | - | - | - | - | - | - | - | - |
| A. versicolor | LC105698 | 4 (10) | 2 | STE (97–477) | 1 (13) | 1 | STE (422) | 1 (4) | 1 | STE (54) | 2 (9) | - | - |
| Penicillium species | |||||||||||||
| P. chrysogenum | KP836338 | 5 (39) | 1 | ROQ C (22) | - | - | - | 3 (38) | 1 | ROQ C (278) | 2 (4) | - | - |
| P. expansum | AY818338 | - | - | - | 2 (33) | - | - | - | - | - | - | - | - |
| P. aethiopicum | KX215125.1 | - | - | - | - | - | - | 3 (38) | - | - | - | - | - |
| P. camemberti | KT355012 | 2 (15) | - | - | - | - | - | - | - | - | - | - | - |
| P. citrinum | KX215134.1 | 3 (23) | - | - | 1 (17) | - | - | - | - | - | - | - | - |
| P. crustosum | KT735107.1 | 2 (15) | - | - | - | - | - | 2 (25) | - | - | 3 (6) | - | - |
| P. raistrickii | KX215126.1 | - | - | - | 2 (33) | - | - | - | - | - | - | - | - |
| P. mallochi | KX215135.1 | - | - | - | 1 (17) | - | - | - | - | - | - | - | - |
| Fusarium species | |||||||||||||
| F. chlamydosporum | KX215137.1 | 1 (43) | - | - | - | - | - | - | - | - | - | - | - |
| F. fujikuroi | KX215132.1 | - | - | - | 1 (33) | - | - | - | - | - | - | - | - |
| F. sporotrichioides | AF404149.1 | 2 (29) | 1, 1, 2 | HT-2 (127), DAS, NEO | - | - | - | - | - | - | - | - | - |
| Fusarium sp. | KP003945 | - | - | - | 2 (67) | 2, 2, 1 | FB3 (79), DON (27-300), 3-AcDON | - | - | - | 2 (40) | 2 | ZEN (180–309) |
| F. verticillioides | KX215138.1 | 4 (57) | 4 | FB1 (77–218) | - | - | - | 3 (100) | 3 | FB1 (92–109) | 3 (60) | 3, 1 | FB2 (103–195) FB3 (88) |
| Isolated Species | Accession No. | Ugba | Ogiri | Iru | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| No. of Iso. Spp. (%) | No. of Tox. Spp. | Toxin Produced (Range: µg/kg) | No. of Iso. Spp. (%) | No. of Tox. Spp. | Toxin Produced (Range: µg/kg) | No. of Iso. Spp. (%) | No. of Tox. Spp. | Toxin Produced (Range: µg/kg) | ||
| A. amstelodami | AY373885.1 | - | - | - | - | - | - | 3 (5) | - | - |
| A. candidus | KT223337 | 2 (7) | - | - | - | - | - | 3 (5) | - | - |
| A. clavatus | KUO52566 | - | - | - | 1 (2) | - | - | 3 (5) | - | - |
| A. flavus | KR611584 | 12 (39) | 10, 12 | AFB1 (27–1889), STE (28–325) | 15 (36) | 9, 9 | AFB1 (96–7406), STE (94–736) | 22 (34) | 16, 4 | AFB1 (82–1723), STE (77–128) |
| A. fumigatus | KU684451 | 3 (8) | - | - | 6 (14) | - | - | 10 (15) | - | - |
| A. minisclerotigenes | JF412778 | - | - | - | 3 (7) | 1 | AFB1 (242) | - | - | - |
| A. niger | KX215111.1 | 3 (8) | 3 | OTA (76–1265) | 4 (10) | 2 | OTA (118–229) | 3 (5) | 1 | OTA (78) |
| A. oryzae | KX215113.1 | - | - | - | 2 (5) | - | - | - | - | - |
| A. parasiticus | DQ467988.1 | 4 (13) | 4, 4, 1 | AFB1 (391–1132), AFB2 (37–504), AFG1 (46) | 6 (14) | 5, 5, 1, 1 | AFB1 (120–1470), AFB2 (83–323), AFG1 (69), AFG2 (97) | 11 (17) | 8, 3, 2, 2 | AFB1 (206–445), AFB2 (51–340), AFG1 (36–59), AFG2 (49–71) |
| A. ruber | KX215127.1 | - | - | - | - | - | - | 1 (2) | - | - |
| A. spp | KX215117.1 | 1 (3) | - | - | - | - | - | - | - | - |
| A. sclerotiorum | KT717312 | 3 (10) | 1 | OTA (161) | - | - | - | 2 (3) | - | - |
| A. sydowii | KX215130.1 | - | - | - | - | - | - | 2 (3) | - | - |
| A. tritici | KX215119.1 | - | - | - | - | - | - | - | - | - |
| A. tubingensis | KT717311 | 2 (7) | - | - | - | - | - | 2 (3) | - | - |
| A. ustus | HQ607918.1 | - | - | 3 (7) | - | - | - | - | - | |
| A. versicolor | LC105698 | 1 (3) | 1 | STE (101) | 2 (5) | 2 | STE (89–500) | 3 (5) | 1 | STE (89) |
| P. chrysogenum | KX215133.1 | 4 (29) | 4 | ROQ C (360–1260) | 6 (30) | - | - | 9 (30) | 6 | ROQ C (13–57) |
| P. expansum | AY818338 | 1 (7) | - | - | - | - | - | 2 (7) | - | - |
| P. polonicum | KX215146.1 | - | - | - | - | - | - | 1 (3) | - | - |
| P. lanosocoeruleum | JX997110 | - | - | - | - | - | - | 2 (7) | - | - |
| P. aethiopicum | KX215125.1 | 2 (14) | - | - | 2 (10) | - | - | - | - | - |
| P. camemberti | KT355012 | - | - | - | - | - | - | 1 (3) | - | - |
| p. citrinum | KX215122.1 | - | 2 (10) | - | - | 2 (7) | - | |||
| p. verrucosum | KM115130 | 2 (14) | 1 | OTA (15) | 1 (5) | 1 | OTA (19) | 3 (10) | 2 | OTA (18–32) |
| p. crustosum | KT735107.1 | 2 (14) | - | - | 3 (15) | - | - | - | - | - |
| P. flavigenum | LN809058 | - | - | - | 2 (10) | - | - | - | - | - |
| P. raistrickii | KX215126.1 | - | - | - | 4 (20) | - | - | 3 (10) | - | - |
| P. glabrum | JN887323.1 | 2 (14) | - | - | - | - | - | 1 (3) | - | - |
| P. rubens | LC105692 | - | - | - | - | - | - | 2 (7) | - | - |
| P. steckii | KX215128.1 | 1 (7) | - | - | - | - | - | 1 (3) | - | - |
| P. mallochi | KX215135.1 | - | - | - | - | - | - | 3 (10) | - | - |
| F. andiyazi | KX215140.1 | 2 (25) | - | - | - | - | - | - | - | - |
| F. chlamydosporum | KP769538.1 | - | - | - | - | - | - | 4 (32) | 1 | DAS |
| F. fujikuroi | KT192328 | - | - | - | 1 (10) | - | - | - | - | - |
| F. proliferatum | KP773280 | 2 (25) | - | - | 2 (20) | - | - | 2 (16) | - | - |
| F. sporotrichioides | AF404149.1 | 1 (13) | 1 | NEO | 2 (20) | 2, 2 | T-2 (134), DAS | 2 (8) | 2, 2, 2 | T-2 (139–1749), DAS, FUS-X |
| Fusarium sp. | JQ350882 | 1 (13) | 1, 1, 1 | DON (870), ZEN (197), 3-AcDON | - | - | - | 1 (8) | 1, 1 | ZEN (139) |
| F. verticillioides | KX215131.1 | 2 (38) | 1, 1, 1 | FB1 (81), FB2 (63), FB3 (205) | 5 (50) | 1, 3 | FB2 (234), FB3 (79–148) | 4 (31) | - | - |
| Component | Abbreviation | Precursor Ion (m/z) | Product Ion (m/z) | Cone Voltage (V) | Collision Energy (eV) | Expected Retention Time (min) |
|---|---|---|---|---|---|---|
| * Kojic acid | KA | 143.1 | 69.1 125.1 | 35 | 30 | 3.70 |
| * Methylisocoumarin | ME-ISOC | 307.1 | 149.1 247.1 | 35 | 35 | 8.68 |
| Aflatoxin G2 | AFG2 | 331.1 | 245.1 313.1 | 25 | 18 | 8.95 |
| Aflatoxin G1 | AFG1 | 329.1 | 243.1 311.0 | 35 | 25 | 9.00 |
| Aflatoxin B2 | AFB2 | 315.1 | 259.1 286.9 | 25 | 35 | 9.43 |
| * Speradine A | SPRE | 367.2 | 160.1 266.1 | 35 | 35 | 9.72 |
| Aflatoxin B1 | AFB1 | 313.1 | 270.1 285.1 | 70 | 35 | 9.92 |
| * Sterigmatocystin Analogue | STE-A | 325.1 | 281.1 310.1 | 35 | 34 | 9.95 |
| * Ochratoxin A | OTA | 214.0 | 142.0 152.0 | 51 | 25 | 9.96 |
| * Oxy-o-methyl Sterigmatocystin | OxyHOMST | 371.1 | 282.1 315.1 | 35 | 35 | 10.10 |
| * Dihydroxyl-o-methyl STE | DHoxyHOMST | 373.1 | 322.1 355.1 | 35 | 35 | 10.32 |
| * Aflavarin | AFV | 455.2 | 379.2 413.2 | 35 | 40 | 10.35 |
| * Aspertoxin 1 | ASPT | 355.1 | 322.1 340.1 | 35 | 30 | 10.51 |
| * Aspertoxin 2 | ASPT-2 | 355.1 | 322.1 340.1 | 35 | 40 | 10.53 |
| * Aspergillic acid | AA | 225.2 | 165.1 207.2 | 35 | 40 | 10.70 |
| * Versiconol | VOH | 361.1 | 285.1 325.1 | 35 | 35 | 11.13 |
| * Aflavarin-Analog 2 | AFV-2 | 425.1 | 334.1 383.1 | 35 | 35 | 11.24 |
| * Dihydro-o-methyl Sterigmatocystin | DHOMST | 341.1 | 285.1 326.1 | 35 | 35 | 11.49 |
| * Versiconal Hemiacetal Acetate | VHA | 401.1 | 283.1 307.1 | 35 | 35 | 11.52 |
| * Flavacol | FLV | 209.2 | 123.1 137.1 | 35 | 40 | 11.78 |
| * Dehydro-Aflavanine | DH-AF | 438.3 | 285.2 402.3 | 35 | 40 | 12.17 |
| * O-methyl Sterigmatocystin | OMST | 339.1 | 306.1 324.1 | 35 | 35 | 12.69 |
| * Aflavarin-Analog 1 | AFV-1 | 439.1 | 365.1 397.1 | 35 | 35 | 12.93 |
| * Noranthrone | NORA | 357.2 | 245.1 273.1 | 35 | 35 | 13.02 |
| * Aflatrem | AFTR | 502.3 | 156.1 198.1 | 35 | 40 | 13.17 |
| * Paspalinine | PASL | 434.2 | 130.1 376.2 | 35 | 40 | 13.78 |
| Sterigmatocystin | STE | 325.0 | 281.1 310.1 | 35 | 34 | 14.28 |
| * Cyclopiazonic acid | CPA | 337.2 | 140.1 196.1 | 35 | 40 | 14.29 |
| * Hydroxyneoaspergillic acid | OH-AA | 241.2 | 137.1 163.1 | 35 | 40 | 14.31 |
| * ß-Cyclopiazonic acid | ß-CPA | 339.2 | 154.1 198.1 | 35 | 40 | 14.75 |
| * Dytryptohenaline | DYT | 693.3 | 318.2 346.2 | 35 | 40 | 14.80 |
| * Paspaline | PAS | 422.3 | 158.1 386.3 | 35 | 35 | 15.04 |
| * Leporin C | LEO-C | 336.2 | 200.1 214.1 | 35 | 40 | 17.40 |
| * Aflavinine | AFN | 406.3 | 180.1 224.3 | 35 | 45 | 19.38 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adekoya, I.; Njobeh, P.; Obadina, A.; Landschoot, S.; Audenaert, K.; Okoth, S.; De Boevre, M.; De Saeger, S. Investigation of the Metabolic Profile and Toxigenic Variability of Fungal Species Occurring in Fermented Foods and Beverage from Nigeria and South Africa Using UPLC-MS/MS. Toxins 2019, 11, 85. https://doi.org/10.3390/toxins11020085
Adekoya I, Njobeh P, Obadina A, Landschoot S, Audenaert K, Okoth S, De Boevre M, De Saeger S. Investigation of the Metabolic Profile and Toxigenic Variability of Fungal Species Occurring in Fermented Foods and Beverage from Nigeria and South Africa Using UPLC-MS/MS. Toxins. 2019; 11(2):85. https://doi.org/10.3390/toxins11020085
Chicago/Turabian StyleAdekoya, Ifeoluwa, Patrick Njobeh, Adewale Obadina, Sofie Landschoot, Kris Audenaert, Sheila Okoth, Marthe De Boevre, and Sarah De Saeger. 2019. "Investigation of the Metabolic Profile and Toxigenic Variability of Fungal Species Occurring in Fermented Foods and Beverage from Nigeria and South Africa Using UPLC-MS/MS" Toxins 11, no. 2: 85. https://doi.org/10.3390/toxins11020085
APA StyleAdekoya, I., Njobeh, P., Obadina, A., Landschoot, S., Audenaert, K., Okoth, S., De Boevre, M., & De Saeger, S. (2019). Investigation of the Metabolic Profile and Toxigenic Variability of Fungal Species Occurring in Fermented Foods and Beverage from Nigeria and South Africa Using UPLC-MS/MS. Toxins, 11(2), 85. https://doi.org/10.3390/toxins11020085