The Effect of Cyanobacterial LPS Antagonist (CyP) on Cytokines and Micro-RNA Expression Induced by Porphyromonas gingivalis LPS



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
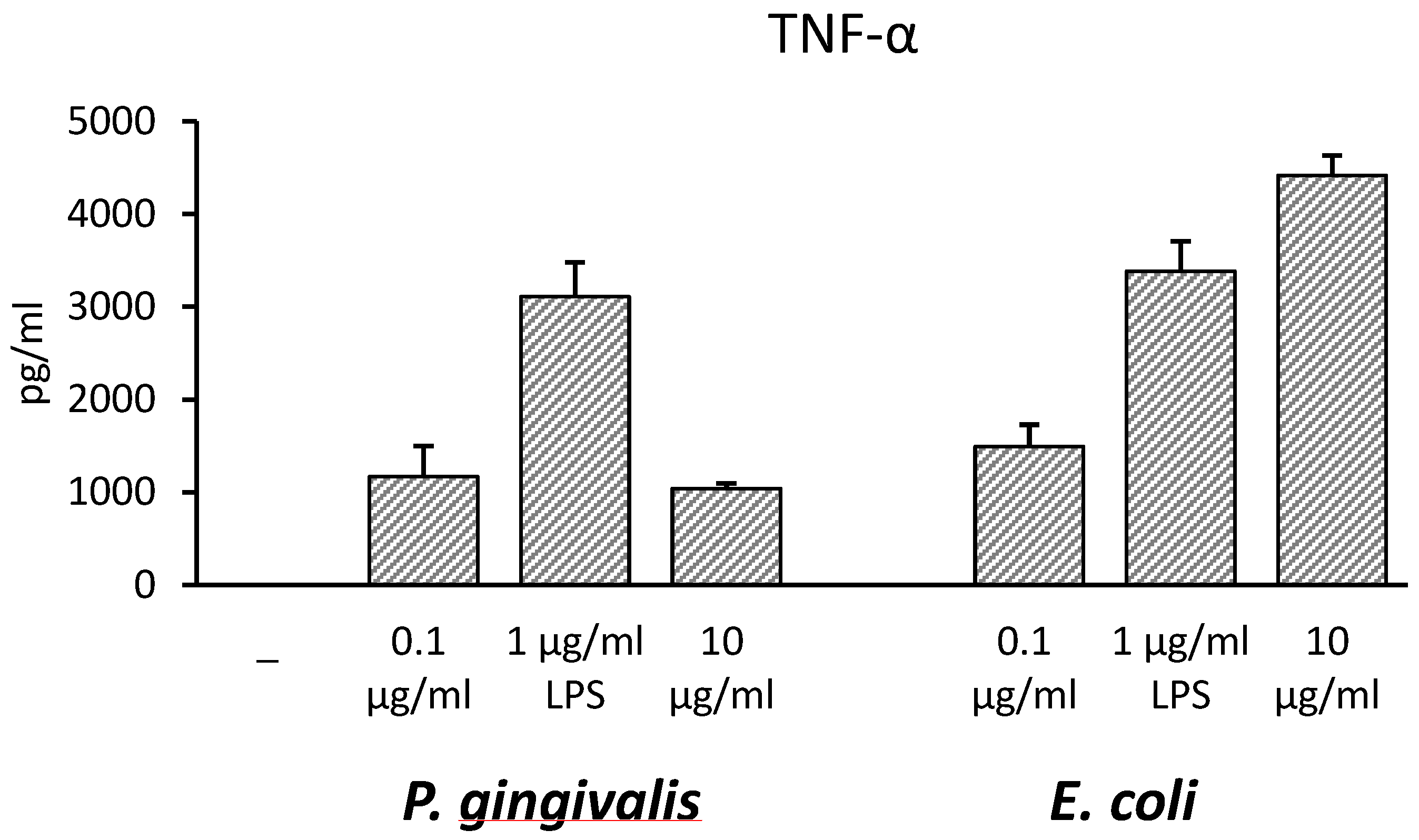
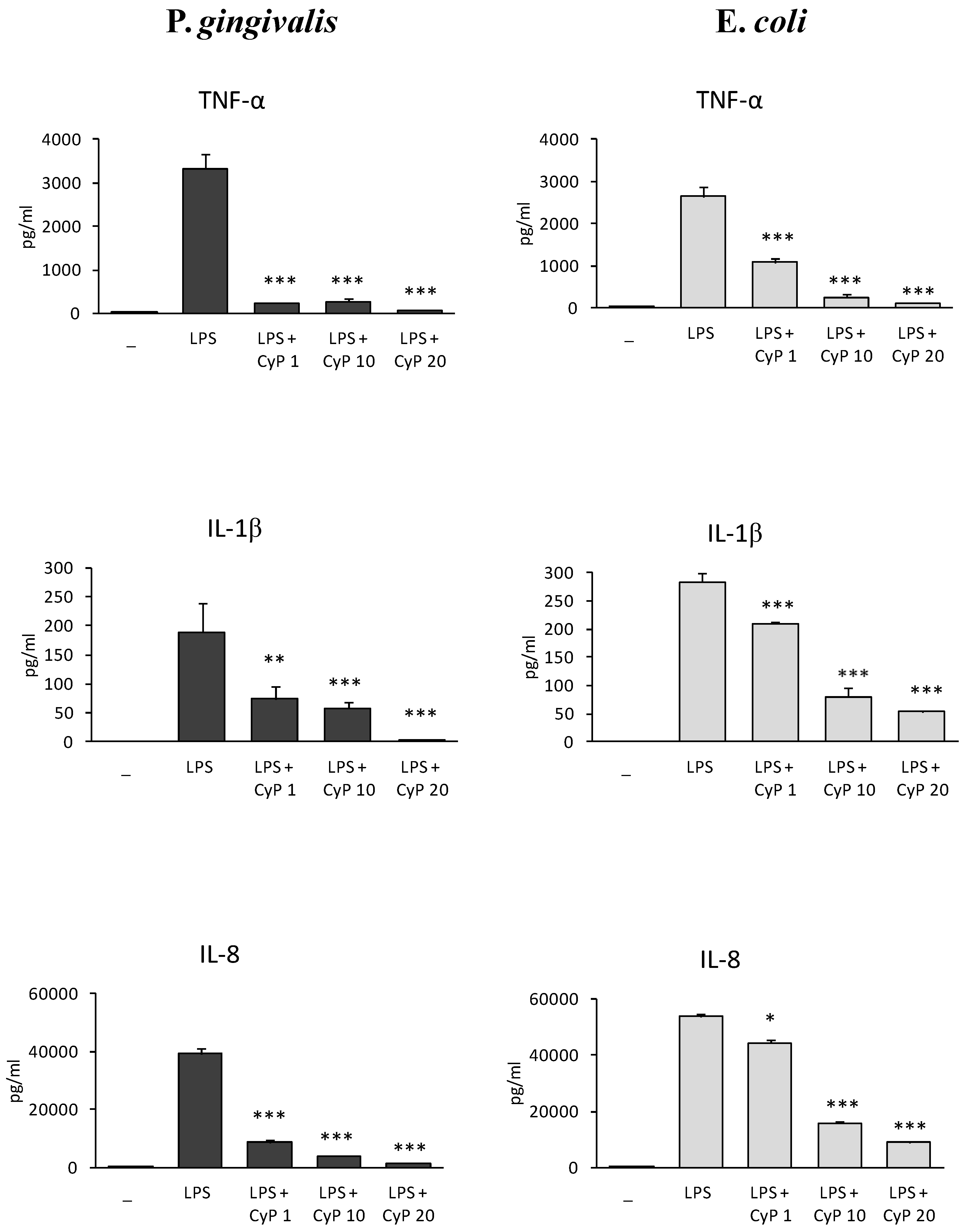
2.1. CyP Dose-Dependently Inhibits Pro-Inflammatory Cytokine Production Induced by Pg-LPS
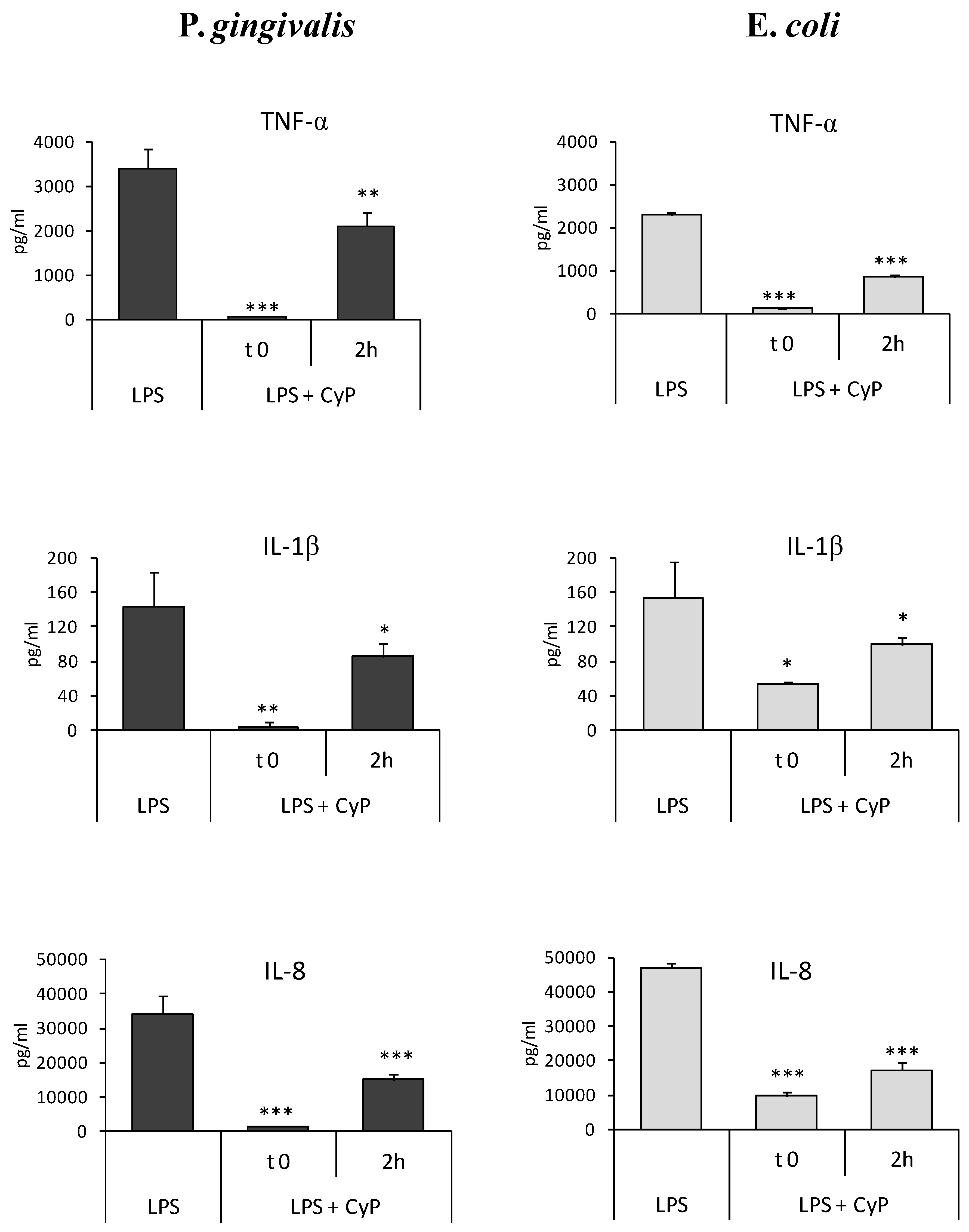
2.2. CyP Inhibits Pro-Inflammatory Cytokine Production Induced by Pg-LPS also When Added Two Hours after LPS
2.3. CyP Differently Affects miRNA Expression in THP-1 Cells Stimulated with Pg-LPS and Ec-LPS
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cells and Reagents
5.2. THP-1 Cell Cultures
5.3. Cytokine ELISA
5.4. RNA Isolation
5.5. Cytokine Quantitative Real-Time PCR
5.6. MiRNA Quantitative Real-Time PCR
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- How, K.Y.; Song, P.K.; Chan, K.G. Porphyromonas gingivalis: An overview of periodontopathic pathogen below the gum line. Front. Microbiol. 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Nussbaum, G.; Shapira, L. How has neutrophil research improved our understanding of periodontal pathogenesis? J. Clin. Periodontol. 2011, 38, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Cekici, A.; Kantarci, A.; Hasturk, H.; Van Dyke, T.E. Inflammatory and immune pathways in the pathogenesis of periodontal disease. Periodontol 2000 2014, 64, 57–80. [Google Scholar] [CrossRef] [PubMed]
- Al-Qutub, M.N.; Braham, P.H.; Karimi-Naser, L.M.; Liu, X.; Genco, C.A.; Darveau, R.P. Hemin-dependent modulation of the lipid A structure of Porphyromonas gingivalis lipopolysaccharide. Infect. Immun. 2006, 74, 4474–4485. [Google Scholar] [CrossRef] [PubMed]
- Curtis, M.A.; Percival, R.S.; Devine, D.; Darveau, R.P.; Coats, S.R.; Rangarajan, M.; Tarelli, E.; Marsh, P.D. Temperature-dependent modulation of Porphyromonas gingivalis lipid A structure and interaction with the innate host defenses. Infect. Immun. 2011, 79, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Graves, D.T.; Oates, T.; Garlet, G.P. Review of osteoimmunology and the host response in endodontic and periodontal lesions. J. Oral. Microbiol. 2011, 3, 1–15. [Google Scholar] [CrossRef]
- Caroff, M.; Karibian, D. Structure of Bacterial Lipopolysaccharides. Carbohydr. Res. 2003, 338, 2431–2447. [Google Scholar] [CrossRef] [PubMed]
- Darveau, R.P.; Pham, T.T.; Lemley, K.; Reife, R.A.; Bainbridge, B.W.; Coats, S.R.; Howald, W.N.; Way, S.S.; Hajjar, A.M. Porphyromonas gingivalis lipopolysaccharide contains multiple lipid A species that functionally interact with both toll-like receptor 2 and 4. Infect. Immun. 2004, 72, 5041–5051. [Google Scholar] [CrossRef] [PubMed]
- Heart, T.D.K.; Darveau, R.P.; Seneviratne, C.; Wang, C.Y.; Wang, Y.; Jin, L. Tetra and penta-acylated lipid A structures of Porphyromonas gingivalis LPS differentially activate TLR4-mediated NF-κB signal transduction cascade and immune-inflammatory response in human gingival fibroblasts. PLoS ONE 2013, 8, e58496. [Google Scholar] [CrossRef]
- Molteni, M.; Gemma, S.; Rossetti, C. The role of Toll-like receptor 4 (TLR4) in infectious and non-infectious inflammation. Mediat. Inflamm. 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kumada, H.; Haishima, Y.; Watanabe, K.; Hasegawa, C.; Tsuchiya, T.; Tanamoto, K.; Umemoto, T. Biological properties of the native and synthetic lipid A of Porphyromonas gingivalis lipopolysaccharide. Oral Microbiol. Immunol. 2008, 23, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Yasuyuki, A.; Hashimoto, M.; Takeuchi, O.; Kurita, T.; Yoshikai, Y.; Miyake, K.; Akira, S. Cell activation by Porphyromonas gingivalis lipid A molecule through Toll-like receptor 4- and myeloid differentiation factor 88-dependent signaling pathway. Int. Immunol. 2002, 14, 1325–1332. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Coats, S.R.; Chang, A.M.; Darveau, R.P. A novel class of lipoprotein lipase-sensitive molecules mediates toll-like receptor 2 activation by Porphyromonas gingivalis. Infect. Immun. 2013, 81, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Nativel, B.; Couret, D.; Giraud, P.; Meilhac, O.; d’Hellencourt, C.L.; Viranaïcken, W.; Silva, C.R. Porphyromonas gingivalis lipopolysaccharides act exclusively through TLR4 with a resilience between mouse and human. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Park, B.S.; Song, D.H.; Kim, H.M.; Choi, B.S.; Lee, H.; Lee, J.O. The structural basis of lipopolysaccharide recognition by the TLR4-MD-2 complex. Nature 2009, 458, 1191–1195. [Google Scholar] [CrossRef] [PubMed]
- Heart, T.D.K.; Wang, Y.; Seneviratne, C.J.; Lu, Q.; Darveau, R.P.; Wang, C.Y.; Jin, L. Porphyromonas gingivalis lipopolysaccharide lipid A heterogeneity differentially modulates the expression of IL-6 and IL-8 in human gingival fibroblasts. J. Clin. Periodontol. 2011, 38, 694–701. [Google Scholar] [CrossRef]
- Olsen, I.; Singhrao, S.K.; Osmudsen, H. Periodontitis, pathogenesis and progression: miRNA-mediated cellular responses to Porphyromonas gingivalis. J. Oral. Microbiol. 2017, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nahid, M.A.; Pauley, K.M.; Satoh, M.; Chan, E.K.L. MiR146a is critical for endotoxin-induced tolerance. J. Biol. Chem. 2009, 284, 34590–34599. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; O’Neill, L.A. MicroRNAs and the resolution phase of inflammation in macrophages. Eur. J. Immunol. 2011, 41, 2470–2485. [Google Scholar] [CrossRef] [PubMed]
- Renzi, T.A.; Rubino, M.; Gornati, L.; Garlanda, C.; Locati, M.; Curtale, G. MiR-146b mediates endotoxin tolerance in human phagocytes. Mediat. Inflamm. 2015, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Sun, M.; Xia, Y.; Shu, R. An RNA-seq screen of P. gingivalis LPS treated human gingival fibroblasts. Arch. Oral Biol. 2018, 88, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ji, Z.; Li, S.; Sun, Y.N.; Liu, J.; Liu, Y.; Tian, W.; Zhou, Y.T.; Shang, X.M. MiR-146a-5p antagonized AGEs- and P.g-LPS-induced ABCA1 and ABCG1 dysregulation in macrophages via IRAK-1 downregulation. Inflammation 2015, 38, 1761–1768. [Google Scholar] [CrossRef] [PubMed]
- Gemma, S.; Molteni, M.; Rossetti, C. Lipopolysaccharides in cyanobacteria: A brief overview. Adv. Microbiol. 2016, 6, 391–397. [Google Scholar] [CrossRef]
- Carillo, S.; Pieretti, G.; Bedini, E.; Parrilli, M.; Lanzetta, R.; Corsaro, M.M. Structural investigation of the antagonist LPS from the cyanobacterium Oscillatoria planktothrix FP1. Carbohydr. Res. 2014, 388, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Molteni, M.; Bosi, A.; Rossetti, C. Natural products with Toll-Like Receptor 4 (TLR4) antagonist activity. Int. J. Inflamm. 2018. [Google Scholar] [CrossRef] [PubMed]
- Macagno, A.; Molteni, M.; Rinaldi, A.; Bertoni, F.; Lanzavecchia, A.; Rossetti, C.; Sallusto, F. A cyanobacterial LPS antagonist prevents endotoxin shock and blocks sustained TLR4 stimulation required for cytokine expression. J. Exp. Med. 2006, 203, 1481–1492. [Google Scholar] [CrossRef] [PubMed]
- Jemmett, K.; Macagno, A.; Molteni, M.; Heckels, J.E.; Rossetti, C.; Christodoulides, M. A cyanobacterial LPS antagonist inhibits cytokine production induced by Neisseria meningitidis in a human whole blood model of septicaemia. Infect. Immun. 2008, 76, 3156–3163. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, A.; Hara, Y.; Kaneko, T.; Kato, I. Secretion of IL-1 beta, TNF-alpha, IL-8 and IL-1ra by human polymorphonuclear leukocytes in response to lipopolysaccharides from periodontopathic bacteria. J. Periodontal Res. 1997, 32, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Nahid, M.A.; Rivera, M.; Lucas, A.; Chan, E.K.L.; Kesavalu, L. Polymicrobial infection with periodontal pathogens specifically enhances microRNA miR-146a in ApoE-/- mice during experimental periodontal disease. Infect. Immun. 2011, 79, 1597–1605. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.-F.; Shu, R.; Jiang, S.-Y.; Liu, D.-L.; Zhang, X.-L. Comparison of micro RNA profiles of human periodontal diseased and healthy gingival tissues. Int. J. Oral Sci. 2011, 3, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Huck, O.; Al-Hashemi, J.; Poidevin, L.; Poch, O.; Davideau, J.L.; Tenenbaum, H.; Amar, S. Identification and characterization of microRNA differentially expressed in macrophages exposed to Porphyromonas gingivalis infection. Infect. Immun. 2017, 85, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Honda, T.; Takahashi, N.; Miyauchi, S.; Yamazaki, K. Porphyromonas gingivalis lipopolysaccharide induces miR-146a without altering the production of inflammatory cytokines. Biochem. Biophys. Res. Commun. 2012, 420, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.-Y.; Xue, D.; Xie, Y.-F.; Zhu, D.W.; Dong, Y.Y.; Wei, C.C.; Deng, J.Y. The negative feedback regulation of microRNA-146a in human periodontal ligament cells after Porphyromonas gingivalis lipopolysaccharide stimulation. Inflamm. Res. 2015, 64, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Molteni, M.; Bosi, A.; Saturni, V.; Rossetti, C. MiR-146a induction by cyanobacterial lipopolysaccharide antagonist (CyP) mediates endotoxin cross-tolerance. Sci. Rep. 2018. under review. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molteni, M.; Bosi, A.; Rossetti, C. The Effect of Cyanobacterial LPS Antagonist (CyP) on Cytokines and Micro-RNA Expression Induced by Porphyromonas gingivalis LPS. Toxins 2018, 10, 290. https://doi.org/10.3390/toxins10070290
Molteni M, Bosi A, Rossetti C. The Effect of Cyanobacterial LPS Antagonist (CyP) on Cytokines and Micro-RNA Expression Induced by Porphyromonas gingivalis LPS. Toxins. 2018; 10(7):290. https://doi.org/10.3390/toxins10070290
Chicago/Turabian StyleMolteni, Monica, Annalisa Bosi, and Carlo Rossetti. 2018. "The Effect of Cyanobacterial LPS Antagonist (CyP) on Cytokines and Micro-RNA Expression Induced by Porphyromonas gingivalis LPS" Toxins 10, no. 7: 290. https://doi.org/10.3390/toxins10070290
APA StyleMolteni, M., Bosi, A., & Rossetti, C. (2018). The Effect of Cyanobacterial LPS Antagonist (CyP) on Cytokines and Micro-RNA Expression Induced by Porphyromonas gingivalis LPS. Toxins, 10(7), 290. https://doi.org/10.3390/toxins10070290