Control of Fusarium verticillioides (Sacc.) Nirenberg and Fumonisins by Using a Combination of Crop Protection Products and Fertilization


 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Influence of Growing Season on Insect and Fungal Incidence
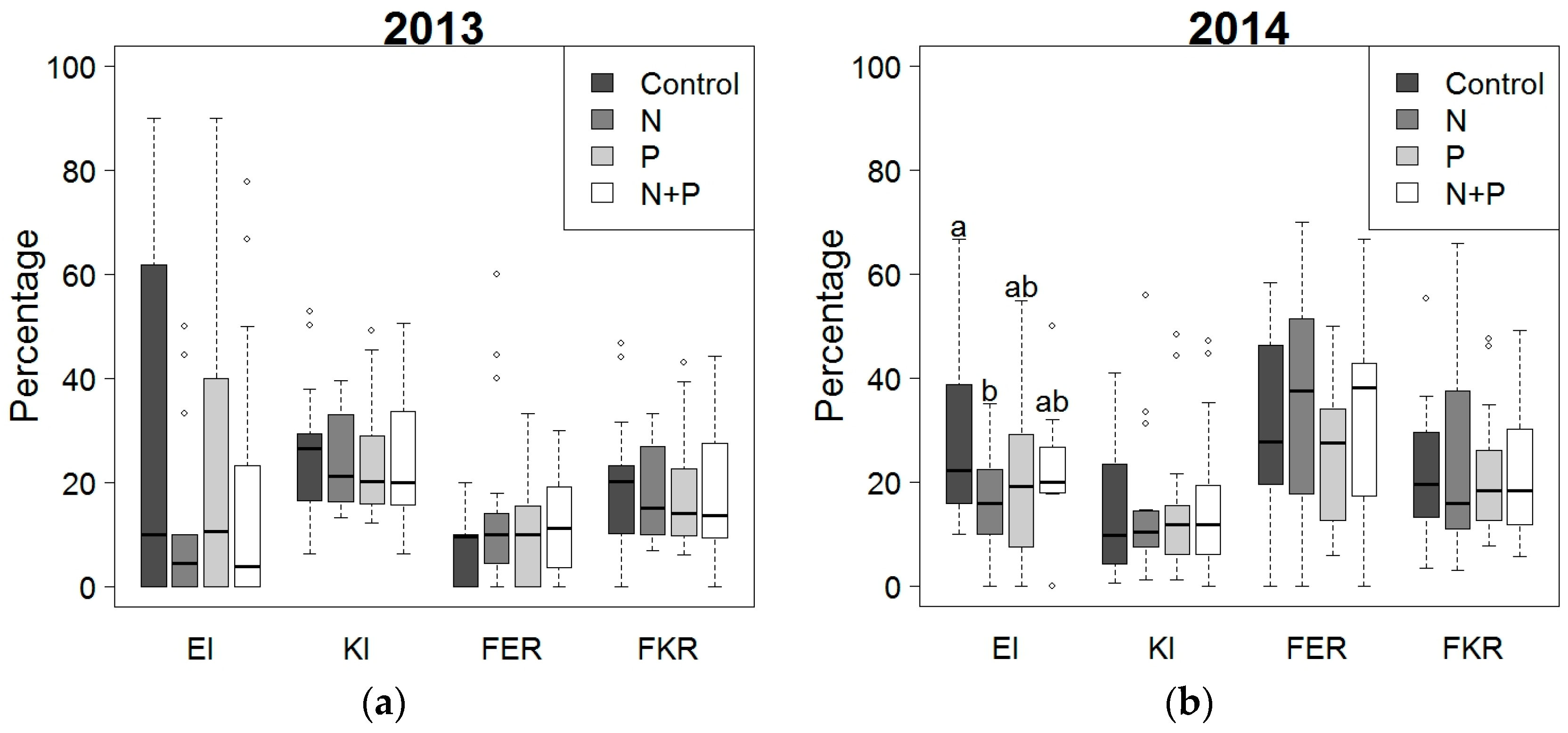
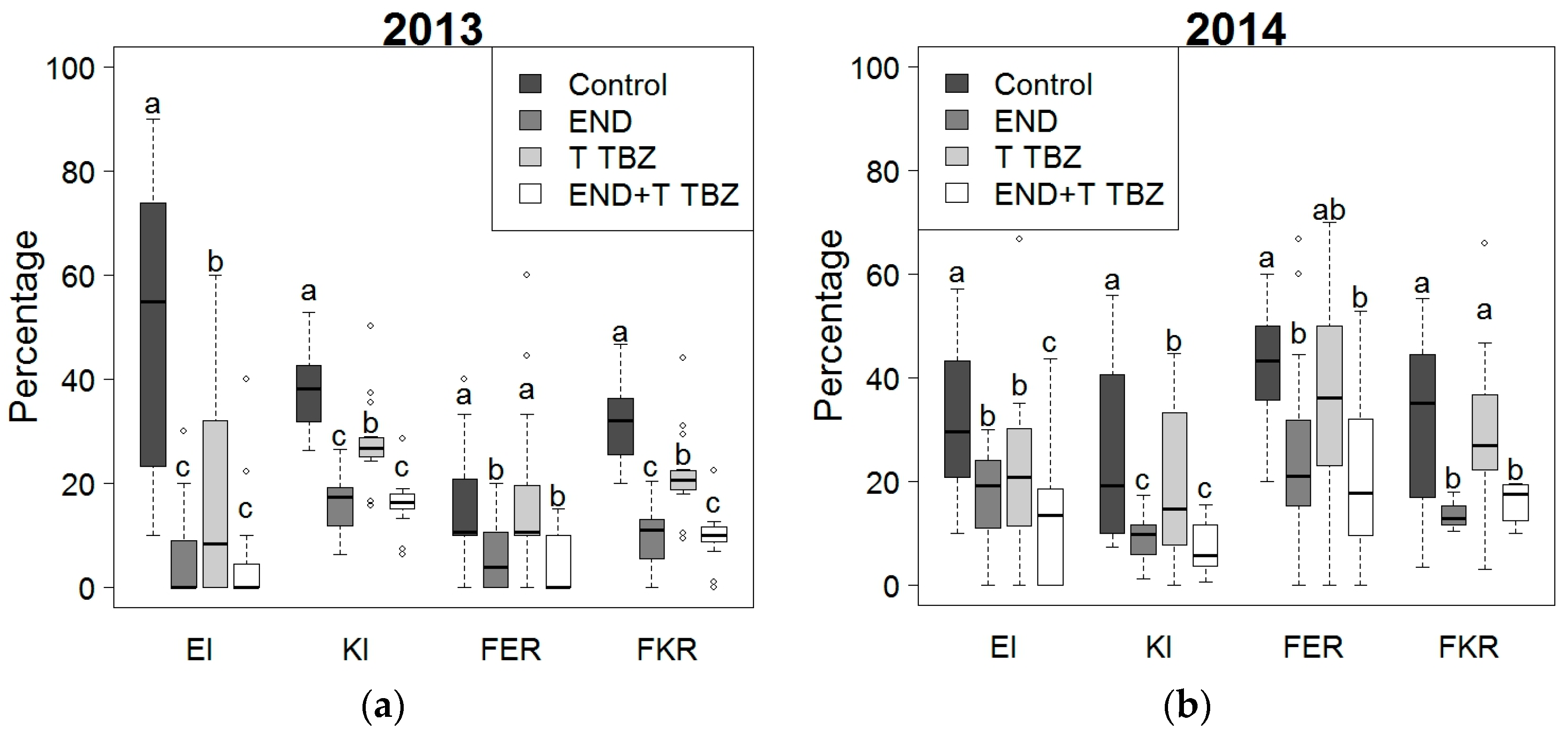
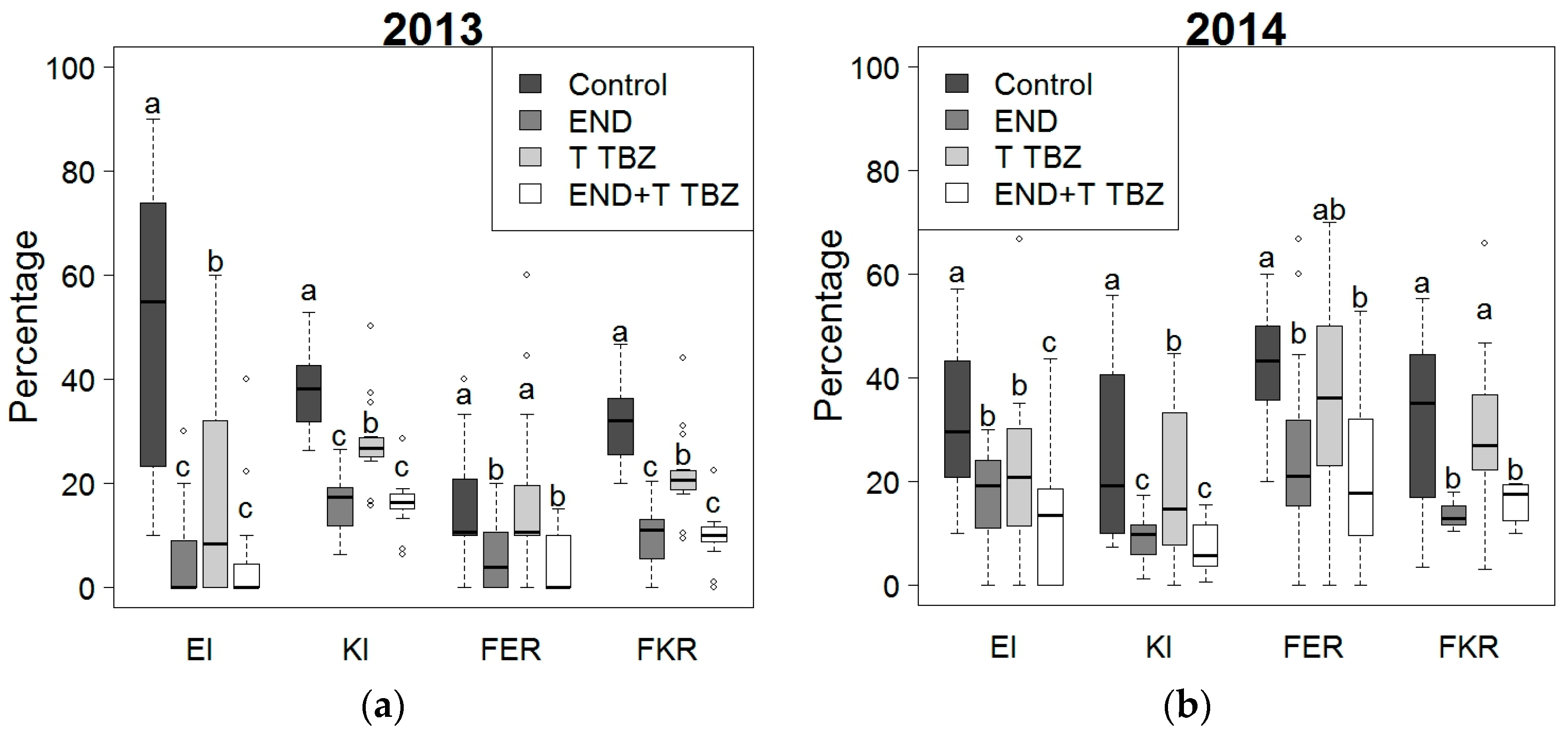
2.2. Effect of Fertilization and Crop Protection on Maize Stalk Borer (MSB) and Fusarium Symptoms
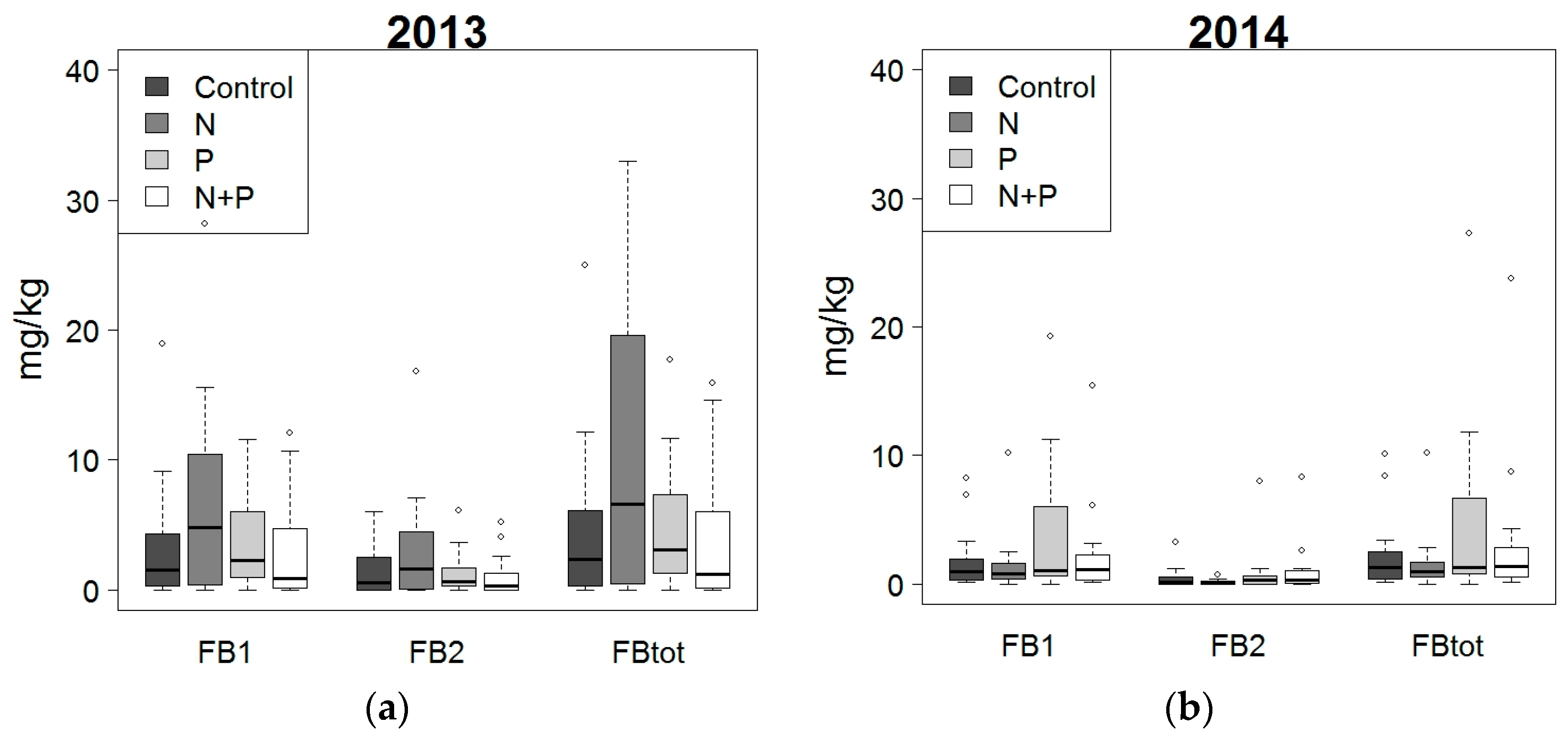
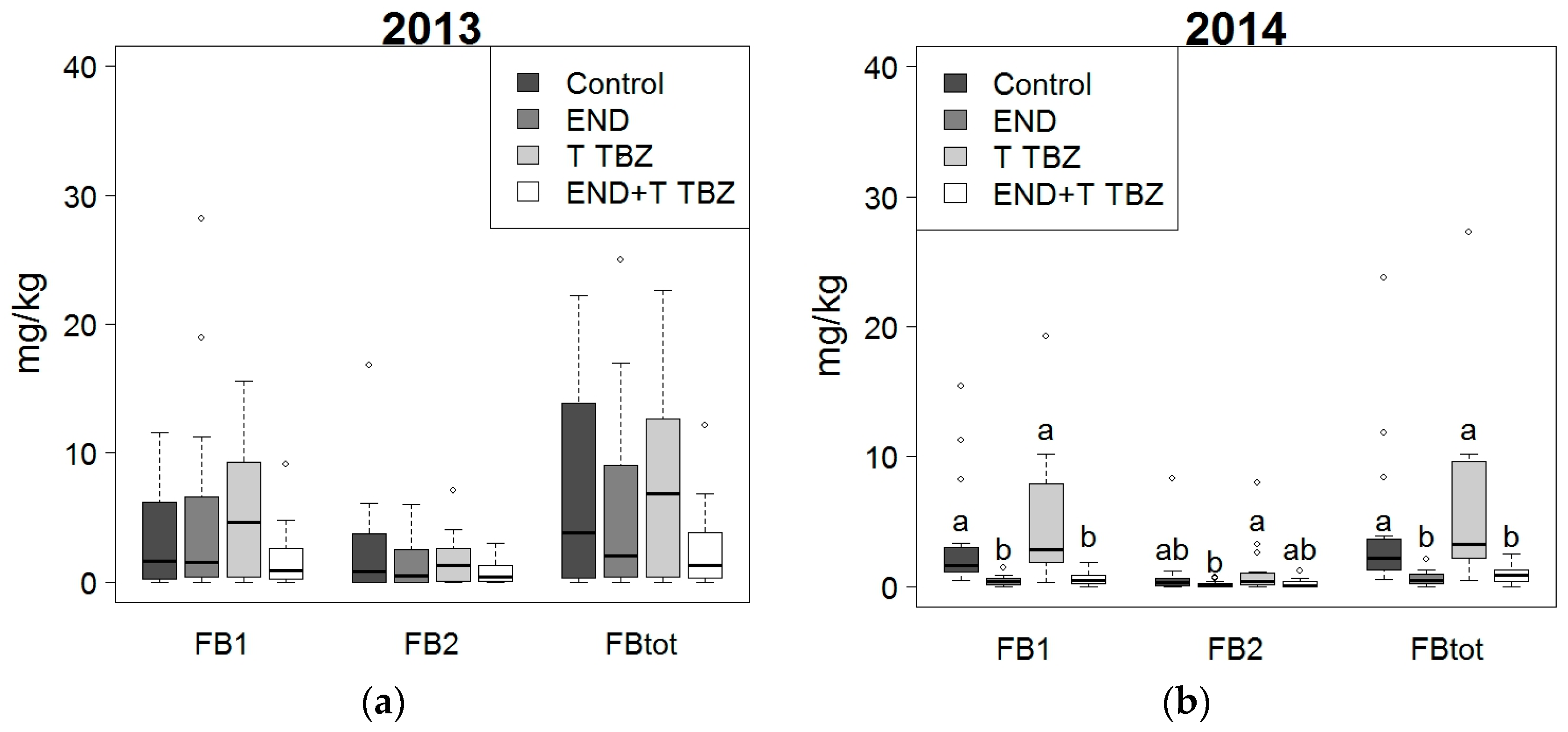
2.3. Effect of Fertilization and Crop Protection on Fumonisins
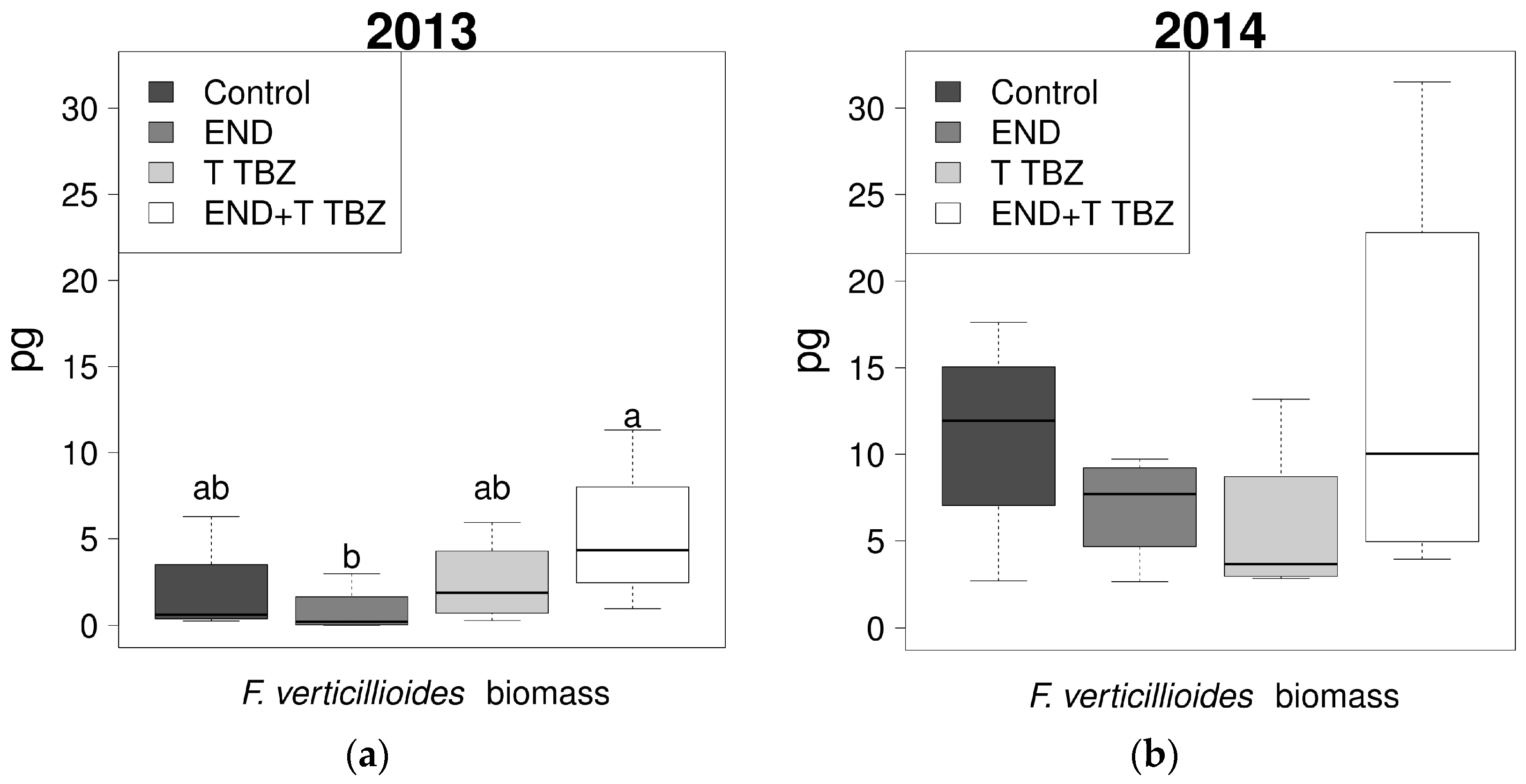
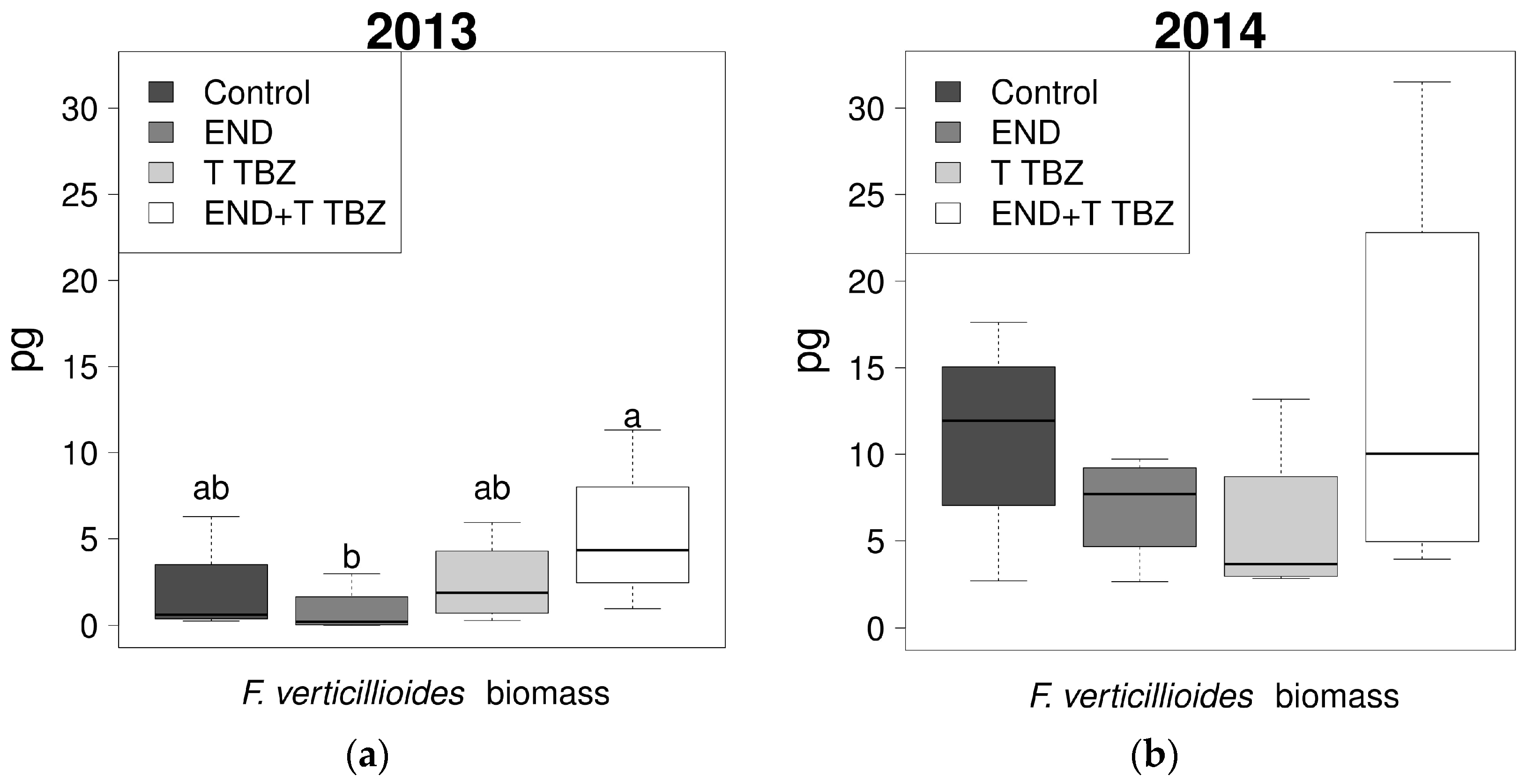
2.4. Effect of Fertilization and Crop Protection on the Biomass of F. verticillioides
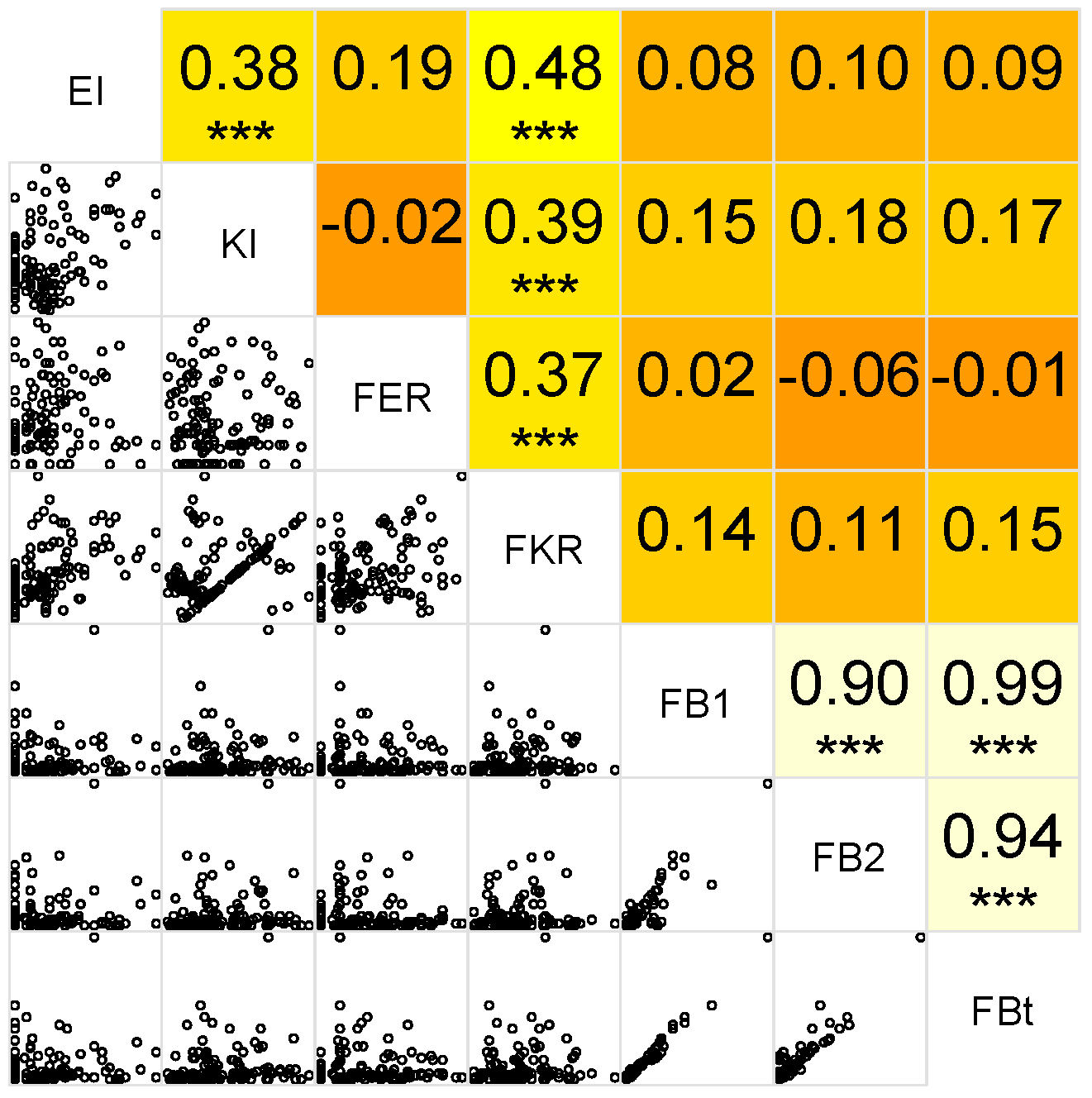
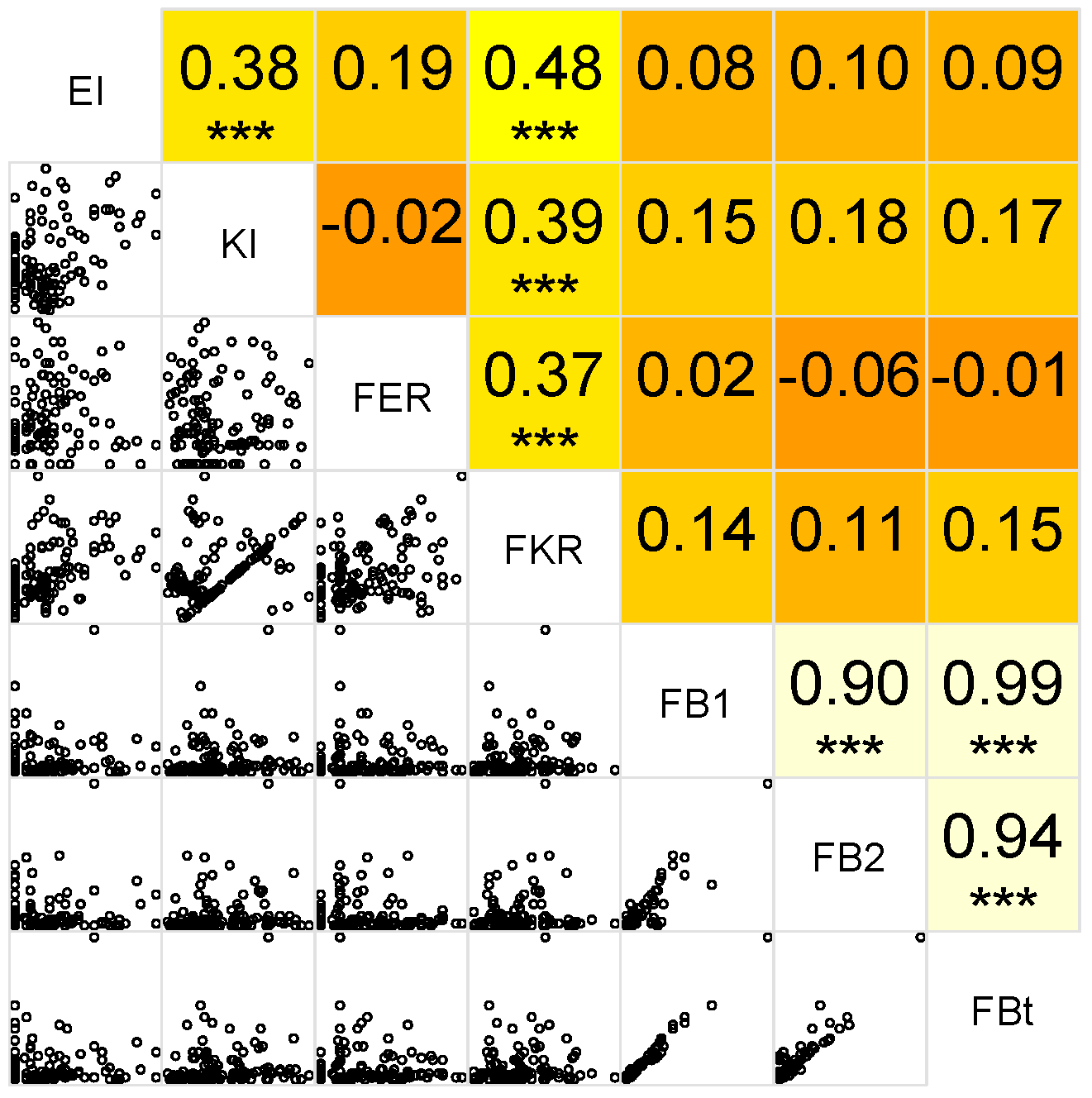
2.5. Correlation between Variables
3. Discussion
4. Materials and Methods
4.1. Description of the Sampling Site
4.2. Experimental Design
4.3. Entomological and Mycological Observations
4.4. Quantification of F. verticillioides Biomass and Fumonisins
4.5. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ranum, P.; Peña-Rosas, J.P.; Garcia-Casal, M.N. Global maize production, utilization, and consumption. Ann. N. Y. Acad. Sci. 2014, 1312, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Lukuyu, B.; Franzel, S.; Ongadi, P.; Duncan, A. Livestock feed resources: Current production and management practices in central and northern rift valley provinces of Kenya. Livest. Res. Rural Dev. 2011, 23, 112. [Google Scholar]
- Hertel, T.W.; Golub, A.A.; Jones, A.D.; O’Hare, M.; Plevin, R.J.; Kammen, D.M. Effects of US maize ethanol on global land use and greenhouse gas emissions: Estimating market-mediated responses. BioScience 2010, 60, 223–231. [Google Scholar] [CrossRef]
- Amare, M.; Asfaw, S.; Shiferaw, B. Welfare impacts of maize-pigeonpea intensification in Tanzania. Agric. Econ. 2012, 43, 27–43. [Google Scholar] [CrossRef]
- Rowhani, P.; Lobell, D.B.; Linderman, M.; Ramankutty, N. Climate variability and crop production in Tanzania. Agric. For. Meteorol. 2011, 151, 449–460. [Google Scholar] [CrossRef]
- Meissle, M.; Mouron, P.; Musa, T.; Bigler, F.; Pons, X.; Vasileiadis, V.; Otto, S.; Antichi, D.; Kiss, J.; Pálinkás, Z. Pests, pesticide use and alternative options in European maize production: Current status and future prospects. J. Appl. Entomol. 2010, 134, 357–375. [Google Scholar] [CrossRef]
- Covarelli, L.; Beccari, G.; Salvi, S. Infection by mycotoxigenic fungal species and mycotoxin contamination of maize grain in Umbria, central Italy. Food Chem. Toxicol. 2011, 49, 2365–2369. [Google Scholar] [CrossRef] [PubMed]
- Menniti, A.; Gregori, R.; Neri, F. Activity of natural compounds on Fusarium verticillioides and fumonisin production in stored maize kernels. Int. J. Food Microbiol. 2010, 136, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Marin, S.; Ramos, A.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Kamala, A.; Ortiz, J.; Kimanya, M.; Haesaert, G.; Donoso, S.; Tiisekwa, B.; De Meulenaer, B. Multiple mycotoxin co-occurrence in maize grown in three agro-ecological zones of Tanzania. Food Control 2015, 54, 208–215. [Google Scholar] [CrossRef]
- Kimanya, M.E.; De Meulenaer, B.; Roberfroid, D.; Lachat, C.; Kolsteren, P. Fumonisin exposure through maize in complementary foods is inversely associated with linear growth of infants in Tanzania. Mol. Nutr. Food Res. 2010, 54, 1659–1667. [Google Scholar] [CrossRef] [PubMed]
- Kimanya, M.E.; De Meulenaer, B.; Tiisekwa, B.; Ndomondo-Sigonda, M.; Devlieghere, F.; Van Camp, J.; Kolsteren, P. Co-occurrence of fumonisins with aflatoxins in home-stored maize for human consumption in rural villages of Tanzania. Food Addit. Contam. 2008, 25, 1353–1364. [Google Scholar] [CrossRef] [PubMed]
- Shirima, C.P.; Kimanya, M.E.; Kinabo, J.L.; Routledge, M.N.; Srey, C.; Wild, C.P.; Gong, Y.Y. Dietary exposure to aflatoxin and fumonisin among Tanzanian children as determined using biomarkers of exposure. Mol. Nutr. Food Res. 2013, 57, 1874–1881. [Google Scholar] [CrossRef] [PubMed]
- Venturini, G.; Assante, G.; Vercesi, A. Fusarium verticillioides contamination patterns in Northern Italian maize during the growing season. Phytopathol. Mediterr. 2011, 50, 110–120. [Google Scholar]
- Murillo-Williams, A.; Munkvold, G. Systemic infection by Fusarium verticillioides in maize plants grown under three temperature regimes. Plant Dis. 2008, 92, 1695–1700. [Google Scholar] [CrossRef]
- Dohlman, E. Mycotoxin Hazards and Regulations Impacts on Food and Animal Feed Crop Trade. In International Trade and Food Safety Economic Theory and Case Studies; Buzby, J.C., Ed.; 2003; Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.201.8435&rep=rep1&type=pdf (accessed on 2 February 2018).
- Jacobsen, B.J. Good agricultural and harvest practices to reduce mycotoxin contamination in wheat in temperate countries. In Mycotoxin Reduction in Grain Chains; Leslie, J.F., Logrieco, A.F., Eds.; Wiley Blackwell: New Delhi, India, 2014; pp. 209–219. [Google Scholar]
- Parsons, M.; Munkvold, G. Associations of planting date, drought stress, and insects with Fusarium ear rot and fumonisin B1 contamination in California maize. Food Addit. Contam. 2010, 27, 591–607. [Google Scholar] [CrossRef] [PubMed]
- Abbas, H.K.; Zablotowicz, R.M.; Weaver, M.A.; Shier, W.T.; Bruns, H.A.; Bellaloui, N.; Accinelli, C.; Abel, C.A. Implications of Bt traits on mycotoxin contamination in maize: Overview and recent experimental results in Southern United States. J. Agric. Food Chem. 2013, 61, 11759–11770. [Google Scholar] [CrossRef] [PubMed]
- Folcher, L.; Delos, M.; Marengue, E.; Jarry, M.; Weissenberger, A.; Eychenne, N.; Regnault-Roger, C. Lower mycotoxin levels in Bt maize grain. Agron. Sustain. Dev. 2010, 30, 711–719. [Google Scholar] [CrossRef]
- Gatch, E.; Munkvold, G. Fungal species composition in maize stalks in relation to European corn borer injury and transgenic insect protection. Plant Dis. 2002, 86, 1156–1162. [Google Scholar] [CrossRef]
- Van Rensburg, J.B.J. First report of field resistance by the stem borer, Busseola fusca(Fuller) to Bt-transgenic maize. S. Afr. J. Plant Soil 2007, 24, 147–151. [Google Scholar] [CrossRef]
- Mourice, S.; Rweyemamu, C.; Nyambilila, A.; Tumbo, S. Narrowing Maize Yield Gaps Under Rain-fed conditions in Tanzania: Effect of Small Nitrogen Dose. Tanzan. J. Agric. Sci. 2014, 12, 821–833. [Google Scholar]
- Walters, D.; Bingham, I. Influence of nutrition on disease development caused by fungal pathogens: Implications for plant disease control. Ann. Appl. Biol. 2007, 151, 307–324. [Google Scholar] [CrossRef]
- Munkvold, G.P. Epidemiology of Fusarium diseases and their mycotoxins in maize ears. Eur. J. Plant Pathol. 2003, 109, 705–713. [Google Scholar] [CrossRef]
- Cao, A.; Santiago, R.; Ramos, A.J.; Souto, X.C.; Aguín, O.; Malvar, R.A.; Butrón, A. Critical environmental and genotypic factors for Fusarium verticillioides infection, fungal growth and fumonisin contamination in maize grown in northwestern Spain. Int. J. Food Microbiol. 2014, 177, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Samapundo, S.; Devliehgere, F.; De Meulenaer, B.; Debevere, J. Effect of water activity and temperature on growth and the relationship between fumonisin production and the radial growth of Fusarium verticillioides and Fusarium proliferatum on corn. J. Food Prot. 2005, 68, 1054–1059. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, J.M.; Nielsen, K.F.; Samson, R.A.; Frisvad, J.C.; Thrane, U. Effect of temperature and water activity on the production of fumonisins by Aspergillus niger and different Fusarium species. BMC Microbiol. 2009, 9, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popovski, S.; Celar, F.A. The impact of environmental factors on the infection of cereals with Fusarium species and mycotoxin production—A review/Vpliv okoljskih dejavnikov na okuzbo zit z glivami Fusarium spp. in tvorbo mikotoksinov-pregledni clanek. Acta Agric. Slov. 2013, 101–105. [Google Scholar] [CrossRef]
- Altieri, M.A.; Nichols, C.I. Soil fertility management and insect pests: Harmonizing soil and plant health in agroecosystems. Soil Tillage Res. 2003, 72, 203–211. [Google Scholar] [CrossRef]
- Hoffland, E.; Jeger, M.J.; van Beusichem, M.L. Effect of nitrogen supply rate on disease resistance in tomato depends on the pathogen. Plant Soil 2000, 218, 239–247. [Google Scholar] [CrossRef]
- Harrison, U.J.; Shew, H. Effects of soil pH and nitrogen fertility on the population dynamics of Thielaviopsis basicola. Plant Soil 2001, 228, 147–155. [Google Scholar] [CrossRef]
- Ochola, D.; Ocimati, W.; Tinzaara, W.; Blomme, G.; Karamura, E. Interactive effects of fertilizer and inoculum concentration on subsequent development of xanthomonas wilt in banana. Aust. J. Agric. Res. 2014, 9, 2727–2735. [Google Scholar]
- Phelan, L.P.; Mason, J.F.; Stinner, B.R. Soil-fertility management and host preference by European corn borer, Ostrinia mubilalis (Hubner), on Zea mays L.: A comparison of organic and conventional chemical farming. Agric. Ecosyst. Environ. 1995, 56, 1–8. [Google Scholar] [CrossRef]
- Ferrigo, D.; Raiola, A.; Causin, R. Plant stress and mycotoxin accumulation in maize. Agrochimica 2014, 58, 116–127. [Google Scholar]
- Reid, L.M.; Zhu, C.; Ma, B.L. Crop rotation and nitrogen effects on maize susceptibility to gibberella (Fusarium graminearum) ear rot. Plant Soil 2001, 237, 1–14. [Google Scholar] [CrossRef]
- Ariño, A.; Herrera, M.; Juan, T.; Estopañan, G.; Carramiñana, J.; Rota, C.; Herrera, A. Influence of agricultural practices on the contamination of maize by fumonisin mycotoxins. J. Food Prot. 2009, 72, 898–902. [Google Scholar] [CrossRef] [PubMed]
- Khattak, A.R.A.; Khalil, S.K. Plant density and nitrogen effects on maize phenology and grain yield. J. Plant Nutr. 2009, 32, 246–260. [Google Scholar]
- Blandino, M.; Reyneri, A.; Vanara, F.; Pascale, M.; Haidukowski, M.; Saporiti, M. Effect of sowing date and insecticide application against European corn borer (Lepidoptera: Crambidae) on fumonisin contamination in maize kernels. Crop Prot. 2008, 27, 1432–1436. [Google Scholar] [CrossRef]
- De Curtis, F.; De Cicco, V.; Haidukowski, M.; Pascale, M.; Somma, S.; Moretti, A. Effects of agrochemical treatments on the occurrence of Fusarium ear rot and fumonisin contamination of maize in Southern Italy. Field Crops Res. 2011, 123, 161–169. [Google Scholar] [CrossRef]
- Saladini, M.A.; Blandino, M.; Reyneri, A.; Alma, A. Impact of insecticide treatments on Ostrinia nubilalis (Hübner) (Lepidoptera: Crambidae) and their influence on the mycotoxin contamination of maize kernels. Pest Manag. Sci. 2008, 64, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- De La Campa, R.; Hooker, D.C.; Miller, J.D.; Schaafsma, A.W.; Hammond, B.G. Modeling effects of environment, insect damage, and Bt genotypes on fumonisin accumulation in maize in Argentina and the Philippines. Mycopathologia 2005, 159, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, E.; Scandolara, A.; Giorni, P.; Pietri, A.; Battilani, P. Field control of Fusarium ear rot, Ostrinia nubilalis (Hubner), and fumonisins in maize kernels. Pest Manag. Sci. 2011, 67, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Alma, A.; Lessio, F.; Reyneri, A.; Blandino, M. Relationships between Ostrinia nubilalis (Lepidoptera: Crambidae) feeding activity, crop technique and mycotoxin contamination of corn kernel in northwestern Italy. Int. J. Pest Manag. 2005, 51, 165–173. [Google Scholar] [CrossRef]
- Miguel Tde, A.; Bordini, J.G.; Saito, G.H.; Andrade, C.G.; Ono, M.A.; Hirooka, E.Y.; Vizoni, E.; Ono, E.Y. Effect of fungicide on Fusarium verticillioides mycelial morphology and fumonisin B(1) production. Braz. J. Microbiol. 2015, 46, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Ebelhar, S.; Hart, C.; Bradley, C. Corn insecticide seed treatment and foliar fungicide effects on corn response to fertilizer nitrogen. In Proceedings of the Illinois Fertilizer Conference 2010, Champaign, IL, USA, 2010. [Google Scholar]
- Rossouw, J.; Van Rensburg, J.; Van Deventer, C. Breeding for resistance to ear rot of maize, caused by Stenocarpella maydis (Berk) Sutton. 1. Evaluation of selection criteria. S. Afr. J. Plant Soil 2002, 19, 182–187. [Google Scholar] [CrossRef]
- Audenaert, K.; Landschoot, S.; Vanheule, A.; Waegeman, W.; De Baets, B.; Haesaert, G. Impact of Fungicide Timing on the Composition of the Fusarium Head Blight Disease Complex and the Presence of deoxynivalenol (DON) in Wheat. In Fungicides-Beneficial and Harmfull Aspects; Thajuddin, N., Ed.; InTech: London, UK, 2011; pp. 79–98. [Google Scholar]
- Becher, R.; Hettwer, U.; Karlovsky, P.; Deising, H.B.; Wirsel, S.G. Adaptation of Fusarium graminearum to tebuconazole yielded descendants diverging for levels of fitness, fungicide resistance, virulence, and mycotoxin production. Phytopathology 2010, 100, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Sydenham, E.W.; Shephard, G.S.; Thiel, P.G. Liquid chromatographic determination of fumonisins B1, B2, and B3 in foods and feeds. J. Agric. Food Chem. 1992, 75, 313–318. [Google Scholar]
- Samapundo, S.; De Meulenaer, B.; De Muer, N.; Debevere, J.; Devlieghere, F. Influence of experimental parameters on the fluorescence response and recovery of the high–performance liquid chromatography analysis of fumonisin B1. J. Chromatogr. A 2006, 1109, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Kamala, A.; Kimanya, M.; Haesaert, G.; Tiisekwa, B.; Madege, R.; Degraeve, S.; Cyprian, C.; De Meulenaer, B. Local post-harvest practices associated with aflatoxin and fumonisin contamination of maize in three agro ecological zones of Tanzania. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2016, 33, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Nicolaisen, M.; Supronienè, S.; Nielsen, L.K.; Lazzaro, I.; Spliid, N.H.; Justesen, A.F. Real-time PCR for quantification of eleven individual Fusarium species in cereals. J. Microbiol. Methods 2009, 76, 234–240. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Rainfall (mm) | Dry Days | Rain Days | Temperature (°C) | Humidity (%) | |
|---|---|---|---|---|---|---|
| 2013 | March | 278.0 | 16 | 15 | 27.09 | 58.71 |
| April | 230.4 | 11 | 19 | 23.40 | 64.93 | |
| May | 82.0 | 19 | 12 | 24.57 | 56.35 | |
| June | 16.7 | 26 | 5 | 22.69 | 44.7 | |
| Total/average | 607.1 | 72 | 51 | 24.44 | 56.17 | |
| 2014 | March | 182.7 | 9 | 21 | 26.97 | 57.61 |
| April | 231.0 | 10 | 21 | 25.78 | 75.88 | |
| May | 113.0 | 14 | 17 | 24.17 | 65.43 | |
| June | 24.0 | 28 | 3 | 22.86 | 58.4 | |
| Total/average | 550.7 | 61 | 62 | 24.95 | 64.33 | |
| Year | EI (%) | KI (%) | FER (%) | FKR (%) | F. verticillioides (pg) | FB1 (μg/kg) | FB2 (μg/kg) |
|---|---|---|---|---|---|---|---|
| 2013 | 19.36 a | 24.59 a | 11.04 b | 18.34 a | 2.39 b | 4863.25 a | 1674.61 a |
| 2014 | 19.13 a | 15.24 b | 34.19 a | 22.25 a | 9.43 a | 2407.10 b | 626.73 b |
| Trade Name | Active Ingredient | Dose/ha |
|---|---|---|
| Thionex (35EC) | Endosulfan (350 g/L) | 2000 mL |
| Rustal (375EC) | 125 g/L triadimenol + 250 g/L tebuconazole | 500 mL |
| Triple super phosphate | Phosphorus (45% P2O5) | 30 kg P2O5 |
| Urea | Nitrogen (46% N) | 40 kg N |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madege, R.R.; Audenaert, K.; Kimanya, M.; Tiisekwa, B.; De Meulenaer, B.; Bekaert, B.; Landschoot, S.; Haesaert, G. Control of Fusarium verticillioides (Sacc.) Nirenberg and Fumonisins by Using a Combination of Crop Protection Products and Fertilization. Toxins 2018, 10, 67. https://doi.org/10.3390/toxins10020067
Madege RR, Audenaert K, Kimanya M, Tiisekwa B, De Meulenaer B, Bekaert B, Landschoot S, Haesaert G. Control of Fusarium verticillioides (Sacc.) Nirenberg and Fumonisins by Using a Combination of Crop Protection Products and Fertilization. Toxins. 2018; 10(2):67. https://doi.org/10.3390/toxins10020067
Chicago/Turabian StyleMadege, Richard Raphael, Kris Audenaert, Martin Kimanya, Bendantukuka Tiisekwa, Bruno De Meulenaer, Boris Bekaert, Sofie Landschoot, and Geert Haesaert. 2018. "Control of Fusarium verticillioides (Sacc.) Nirenberg and Fumonisins by Using a Combination of Crop Protection Products and Fertilization" Toxins 10, no. 2: 67. https://doi.org/10.3390/toxins10020067
APA StyleMadege, R. R., Audenaert, K., Kimanya, M., Tiisekwa, B., De Meulenaer, B., Bekaert, B., Landschoot, S., & Haesaert, G. (2018). Control of Fusarium verticillioides (Sacc.) Nirenberg and Fumonisins by Using a Combination of Crop Protection Products and Fertilization. Toxins, 10(2), 67. https://doi.org/10.3390/toxins10020067