Fatal Neurotoxicosis in Dogs Associated with Tychoplanktic, Anatoxin-a Producing Tychonema sp. in Mesotrophic Lake Tegel, Berlin

 ,
,
Abstract
1. Introduction
2. Results
2.1. Animal Symptoms and Pathology
2.2. Macrophyte and Phytoplankton Samples
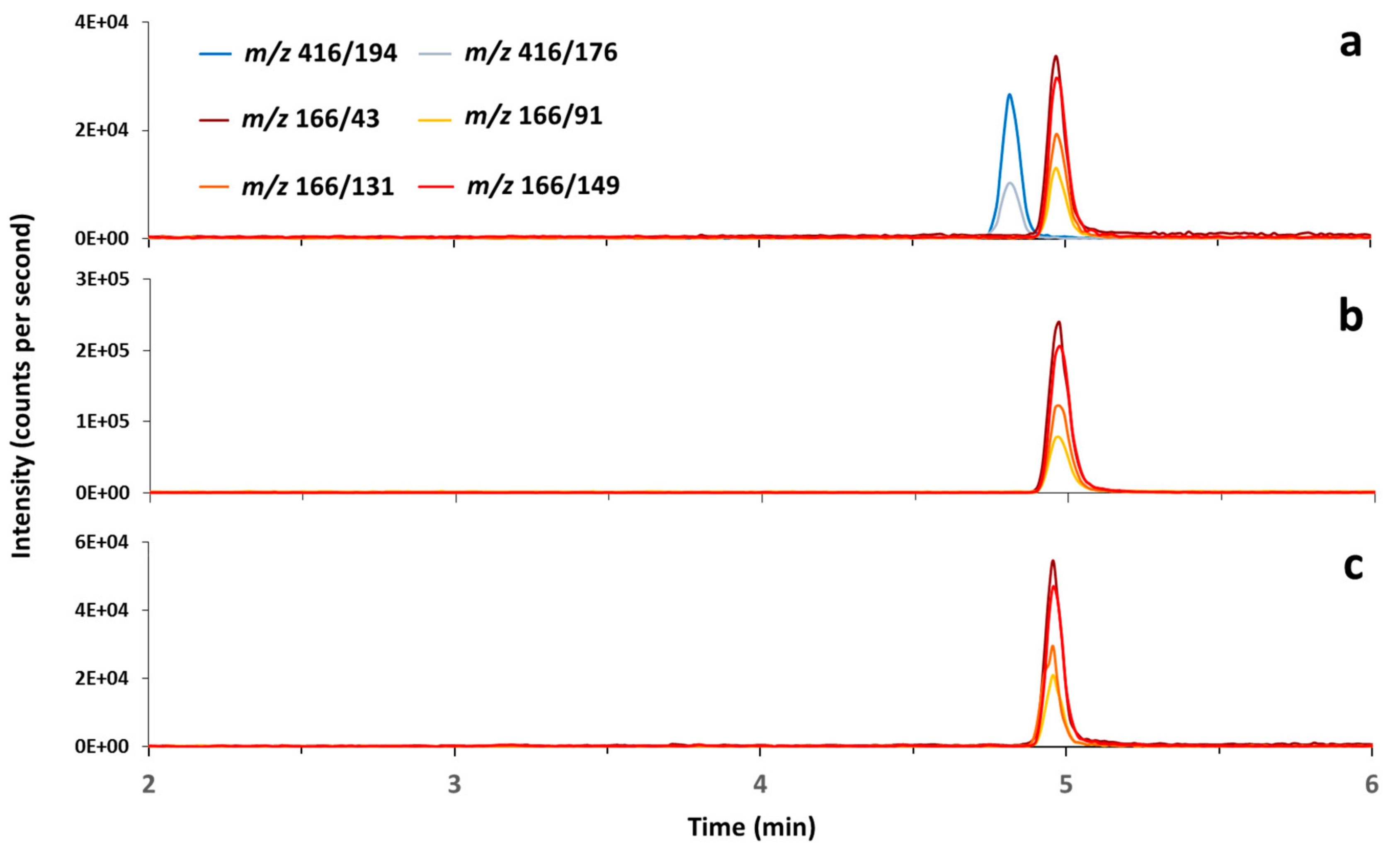
2.3. Cyanobacterial and Other Toxins
3. Discussion
4. Materials and Methods
4.1. Study Site
4.2. Animal Necroscopy
4.3. Macrophyte, Phytoplankton and Toxin Samples
4.4. Toxin Analysis
4.4.1. Cyanotoxins
4.4.2. Other Toxins
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef] [PubMed]
- Fastner, J.; Neumann, U.; Wirsing, B.; Weckesser, J.; Wiedner, C.; Nixdorf, B.; Chorus, I. Microcystins (hepatotoxic heptapeptides) in German fresh water bodies. Environ. Toxicol. 1999, 14, 13–22. [Google Scholar] [CrossRef]
- Mantzouki, E.; Visser, P.M.; Bormans, M.; Ibelings, B.W. Understanding the key ecological traits of cyanobacteria as a basis for their management and control in changing lakes. Aquat. Ecol. 2016, 50, 333–350. [Google Scholar] [CrossRef]
- Directive 2006/7/EC of the European Parliament and of the Council of 15 February 2006 Concerning the Management of Bathing Water Quality and repealing Directive 76/160. EEC 2006.
- Catherine, Q.; Susanna, W.; Isidora, E.-S.; Mark, H.; Aurélie, V.; Jean-François, H. A review of current knowledge on toxic benthic freshwater cyanobacteria - Ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Hamilton, D.P.; Paul, W.J.; Safi, K.A.; Williamson, W.M. New Zealand Guidelines for Cyanobacteria in Recreational fresh WATERS: Interim Guidelines (Commissioned Report for External Body); Ministry for the Environment: Wellington, New Zealand, 2009. [Google Scholar]
- Mez, K.; Beattie, K.A.; Codd, G.A.; Hanselmann, K.; Hauser, B.; Naegeli, H.; Preisig, H.R. Identification of a microcystin in benthic cyanobacteria linked to cattle deaths on alpine pastures in Switzerland. Eur. J. Phycol. 1997, 32, 111–117. [Google Scholar] [CrossRef]
- Gaget, V.; Humpage, A.R.; Huang, Q.; Monis, P.; Brookes, J.D. Benthic cyanobacteria: A source of cylindrospermopsin and microcystin in Australian drinking water reservoirs. Water Res. 2017, 124, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Edwards, C.; Beattie, K.A.; Scrimgeour, C.M.; Codd, G.A. Identification of anatoxin-a in benthic cyanobacteria (blue-green algae) and in associated dog poisonings at Loch Insh, Scotland. Toxicon 1992, 30, 1165–1175. [Google Scholar] [CrossRef]
- Gugger, M.; Lenoir, S.; Berger, C.; Ledreux, A.; Druart, J.-C.; Humbert, J.-F.; Guette, C.; Bernard, C. First report in a river in France of the benthic cyanobacterium Phormidium favosum producing anatoxin-a associated with dog neurotoxicosis. Toxicon 2005, 45, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Puschner, B.; Hoff, B.; Tor, E.R. Diagnosis of anatoxin-a poisoning in dogs from North America. J. Vet. Diagn. Investig. 2008, 20, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Selwood, A.I.; Rueckert, A.; Holland, P.T.; Milne, J.R.; Smith, K.F.; Smits, B.; Watts, L.F.; Cary, C.S. First report of homoanatoxin-a and associated dog neurotoxicosis in New Zealand. Toxicon 2007, 50, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E.J.; Harkema, L.; Begeman, L.; Lürling, M. First report of (homo)anatoxin-a and dog neurotoxicosis after ingestion of benthic cyanobacteria in The Netherlands. Toxicon 2012, 60, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Bouma-Gregson, K.; Power, M.E.; Bormans, M. Rise and fall of toxic benthic freshwater cyanobacteria (Anabaena spp.) in the Eel river: Buoyancy and dispersal. Harmful Algae 2017, 66, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Codd, G.A.; Edwards, C.; Beattle, K.A.; Barr, W.M.; Gunn, G.J. Fatal attraction to cyanobacteria? Nature 1992, 359, 110–111. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.; Mcdonald, C.; de Hoyos, C.; Mischke, U.; Phillips, G.; Borics, G.; Poikane, S.; Skjelbred, B.; Solheim, A.L.; Van Wichelen, J.; et al. Sustaining recreational quality of European lakes: Minimizing the health risks from algal blooms through phosphorus control. J. Appl. Ecol. 2013, 50, 315–323. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Bormans, M.; Fastner, J.; Visser, P.M. CYANOCOST special issue on cyanobacterial blooms: Synopsis—A critical review of the management options for their prevention, control and mitigation. Aquat. Ecol. 2016, 50, 595–605. [Google Scholar] [CrossRef]
- Heinzmann, B.; Chorus, I. Restoration concept for Lake Tegel, a major drinking and bathing water resource in a densely populated area. Environ. Sci. Technol. 1994, 28, 1410–1416. [Google Scholar] [CrossRef] [PubMed]
- Fastner, J.; Abella, S.; Litt, A.; Morabito, G.; Matthews, D.; Phillips, M.G.; Chorus, I. Combating cyanobacterial proliferation by avoiding or treating inflows with high P load - experiences from eight case studies. Aquat. Ecol. 2016, 50, 367–383. [Google Scholar] [CrossRef]
- Chorus, I.; Schauser, I. Oligotrophication of Lake Tegel and Schlachtensee, Berlin—Analysis of System Components, Causalities and Response Thresholds Compared to Responses of Other Waterbodies; Umweltbundesamt: Dessau-Roßlau, Germany, 2011; p. 157ff. [Google Scholar]
- Hilt, S.; Van De Weyer, K.; Köhler, A.; Chorus, I. Submerged macrophyte responses to reduced phosphorus concentrations in two peri-urban lakes. Restor. Ecol. 2010, 18, 452–461. [Google Scholar] [CrossRef]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota-2. Teil: Oscillatoriales. In Süsswasserflora von Mitteleuropa; Büdel, B., Krienitz, L., Gärtner, G., Schagerl, M., Eds.; Elsevier/Spektrum: Heidelberg, Germany, 2005; p. 759. [Google Scholar]
- Beulker, C.; Gunkel, G. Studies on the occurrence and ecology of meiofauna and benthic algae in the littoral sediment of lakes. Limnologica 1996, 26, 311–326. [Google Scholar]
- Algermissen, D.; Mischke, R.; Seehusen, F.; Göbel, J.; Beineke, A. Lymphoid depletion in two dogs with nodularin intoxication. Vet. Rec. 2011, 169, 15. [Google Scholar] [CrossRef] [PubMed]
- Furey, A.; Crowley, J.; Hamilton, B.; Lehane, M.; James, K.J. Strategies to avoid the mis-identification of anatoxin-a using mass spectrometry in the forensic investigation of acute neurotoxic poisoning. J. Chromatogr. A 2005, 1082, 91–97. [Google Scholar] [CrossRef] [PubMed]
- James, K.J.; Crowley, J.; Hamilton, B.; Lehane, M.; Skulberg, O.; Furey, A. Anatoxins and degradation products, determined using hybrid quadrupole time-of-flight and quadrupole ion-trap mass spectrometry: Forensic investigations of cyanobacterial neurotoxin poisoning. Rap. Comm. Mass Spectrom. 2005, 19, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Lawton, L.A.; Edwards, C. Conventional laboratory methods for cyanotoxins. Adv. Exp. Med. Biol. 2016, 619, 513–537. [Google Scholar]
- Stahnke, H.; Kittlaus, S.; Kempe, G.; Alder, L. Reduction of matrix effects in liquid chromatography-electrospray ionization-mass spectrometry by dilution of the sample extracts: How much dilution is needed? Anal. Chem. 2012, 84, 1474–1482. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, R.G. Limnology, 1st ed.; W.B. Saunders: Philadelphia, PA, USA, 1975. [Google Scholar]
- Shams, S.; Capelli, C.; Cerasino, L.; Ballot, A.; Dietrich, D.R.; Sivonen, K.; Salmaso, N. Anatoxin-a producing Tychonema (Cyanobacteria) in European waterbodies. Water Res. 2015, 69, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Song, G.; Shao, J.; Xiang, X.; Li, Q.; Chen, Y.; Yang, P.; Yu, G. Dynamics and polyphasic characterization of odor-producing cyanobacterium Tychonema bourrellyi from Lake Erhai, China. Environ. Sci. Pollut. Res. Int. 2016, 23, 5420–5430. [Google Scholar] [CrossRef] [PubMed]
- Ríos, V.; Moreno, L.; Prieto, A.I.; Soria-Díaz, M.E.; Frías, J.E.; Cameán, A.M. Comparison of Microcystis aeruginosa (PCC7820 and PCC7806) growth and intracellular microcystins content determined by liquid chromatography-mass spectrometry, enzyme-linked immunosorbent assay anti-Adda and phosphatase bioassay. J. Water Health 2014, 12, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Preußel, K.; Chorus, I.; Fastner, J. Nitrogen limitation promotes accumulation and suppresses release of cylindrospermopsins in cells of Aphanizomenon sp. Toxins 2014, 6, 2932–2947. [Google Scholar] [CrossRef] [PubMed]
- Heath, M.; Wood, S.A.; Young, R.G.; Ryan, K.G. The role of nitrogen and phosphorus in regulating Phormidium sp. (cyanobacteria) growth and anatoxin production. FEMS Microbiol. Ecol. 2016, 92, fiw021. [Google Scholar] [CrossRef] [PubMed]
- Salmaso, N.; Cerasino, L.; Boscaini, A.; Capelli, C. Planktic Tychonema (cyanobacteria) in the large lakes south of the Alps: Phylogenetic assessment and toxigenic potential. FEMS Microbiol. Ecol. 2016, 92, fiw021. [Google Scholar] [CrossRef] [PubMed]
- Méjean, A.; Dalle, K.; Paci, G.; Bouchonnet, S.; Mann, S.; Pichon, V.; Ploux, O. Dihydroanatoxin-a is biosynthesized from proline in Cylindrospermum stagnale PCC 7417: Isotopic incorporation experiments and mass spectrometry analysis. J. Nat. Prod. 2016, 79, 1775–1782. [Google Scholar] [CrossRef] [PubMed]
- DIN EN 15204 Water Quality—Guidance Standard on the Enumeration of Phytoplankton Using Inverted Microscopy (Utermöhl technique); Beuth Verlag: Berlin, Germany, 2006.
- Fastner, J.; Flieger, I.; Neumann, U. Optimised extraction of microcystins from field samples—A comparison of different solvents and procedures. Water Res. 1998, 32, 3177–3181. [Google Scholar] [CrossRef]
- Spoof, L.; Vesterkvist, P.; Lindholm, T.; Meriluoto, J. Screening for cyanobacterial hepatotoxins, microcystins and nodularin in environmental water samples by reversed-phase liquid chromatography- electrospray ionisation mass spectrometry. J. Chromatogr. A 2003, 1020, 105–119. [Google Scholar] [CrossRef]
- Maurer, H.H. Methods and Tables for GC-MS. In Mass Spectral and GC Data of Drugs, Poisons, Pesticides, Pollutants and Their Metabolites, 5th ed.; Maurer, H.H., Pfleger, K., Weber, A., Eds.; Wiley-VCH: Weinheim, Germany, 2017; Volume 1, pp. 2–243. [Google Scholar]
- Guale, F.G.; Stair, E.L.; Johnson, B.W.; Edwards, W.C.; Haliburton, J.C. Laboratory diagnosis of zinc phosphide poisoning. Vet. Hum. Toxicol. 1994, 36, 517–519. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| No. | Breed (Sex) | Body Weight (kg) | Age (Years) | Case History and Symptoms | Treatment | Outcome |
|---|---|---|---|---|---|---|
| D1 | American Bulldog (Female) | 38.5 | 2.5 | Swimming in Lake Tegel, sudden onset of neurological failures of hind limbs, collapse, dyspnea and cyanosis | No treatment | Death |
| D2 | Labrador half-breed (Female) | No data | 1 | Swimming in Lake Tegel, sudden onset of paralysis of all limbs, foam at mouth, apnea | Gastric lavage with sodium bicarbonate, doxapram hydrochloride intravenous | Euthanized |
| D3 | French Bulldog (Female) | 8.8 | 0.9 | Playing at Lake Tegel, sudden onset of paralysis of all limbs, foam at mouth, apathy, dyspnea | No treatment | Death |
| Code | Sample | Organ/Geographical Site | Sampling Date | ATX (µg L−1) |
|---|---|---|---|---|
| D1 | Dog 1 S270/17 | Stomach contents | 8700 | |
| D2 | Dog 2 S298/17 | Stomach contents | 5.7 | |
| D3 | Dog 3 S271/17 | Stomach contents | 7320 | |
| FT1 | Fontinalis/Tychonema | L. Tegel, Reiherwerder | 6 June 2017 (1) | 943 |
| 6 June 2017 (2) | 1870 | |||
| FT2 | Fontinalis/Tychonema | L. Tegel, Scharfenberg | 6 June 2017 | 19.8 |
| FT3 | Fontinalis/Tychonema | L. Tegel, Reiswerder | 10 June 2017 (1) | 24.4 |
| 10 June 2017 (2) | 12.6 | |||
| FT4 | Fontinalis/Tychonema | L. Tegel, Reiherwerder (2 m) | 19 June 2017 | <LOQ |
| L. Tegel, Reiherwerder (4 m) | 0.4 | |||
| S1 | Sand | L. Tegel, Reiswerder | 10 June 2017 | 1.0 |
| W1 | Pelagic water | L. Tegel, Reiherwerder | 6 June 2017 | n.d. |
| W2 | Pelagic water | L. Tegel, Reiherwerder | 30 May 2017 | 1.0 |
| W3 | Pelagic water | L. Tegel, Reiswerder | 10 June 2017 | n.d. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fastner, J.; Beulker, C.; Geiser, B.; Hoffmann, A.; Kröger, R.; Teske, K.; Hoppe, J.; Mundhenk, L.; Neurath, H.; Sagebiel, D.; et al. Fatal Neurotoxicosis in Dogs Associated with Tychoplanktic, Anatoxin-a Producing Tychonema sp. in Mesotrophic Lake Tegel, Berlin. Toxins 2018, 10, 60. https://doi.org/10.3390/toxins10020060
Fastner J, Beulker C, Geiser B, Hoffmann A, Kröger R, Teske K, Hoppe J, Mundhenk L, Neurath H, Sagebiel D, et al. Fatal Neurotoxicosis in Dogs Associated with Tychoplanktic, Anatoxin-a Producing Tychonema sp. in Mesotrophic Lake Tegel, Berlin. Toxins. 2018; 10(2):60. https://doi.org/10.3390/toxins10020060
Chicago/Turabian StyleFastner, Jutta, Camilla Beulker, Britta Geiser, Anja Hoffmann, Roswitha Kröger, Kinga Teske, Judith Hoppe, Lars Mundhenk, Hartmud Neurath, Daniel Sagebiel, and et al. 2018. "Fatal Neurotoxicosis in Dogs Associated with Tychoplanktic, Anatoxin-a Producing Tychonema sp. in Mesotrophic Lake Tegel, Berlin" Toxins 10, no. 2: 60. https://doi.org/10.3390/toxins10020060
APA StyleFastner, J., Beulker, C., Geiser, B., Hoffmann, A., Kröger, R., Teske, K., Hoppe, J., Mundhenk, L., Neurath, H., Sagebiel, D., & Chorus, I. (2018). Fatal Neurotoxicosis in Dogs Associated with Tychoplanktic, Anatoxin-a Producing Tychonema sp. in Mesotrophic Lake Tegel, Berlin. Toxins, 10(2), 60. https://doi.org/10.3390/toxins10020060