Transcriptomic Characterization of the South American Freshwater Stingray Potamotrygon motoro Venom Apparatus

 , and
, and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Denovo Assembly
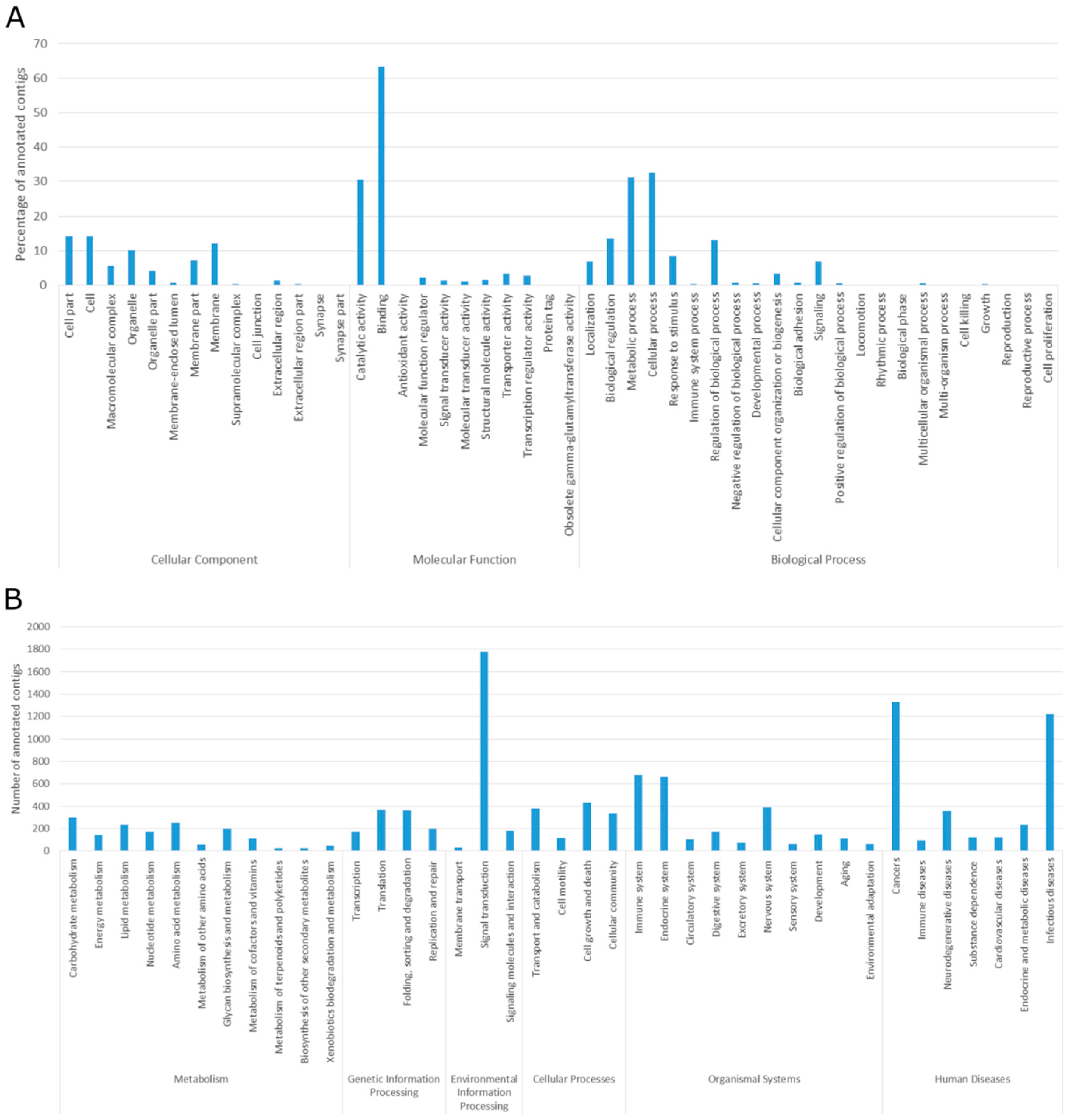
2.2. Transcriptome Functional and Pathway Annotation
2.3. Gene Ontology and Metabolic Pathways
2.4. Venom Expression
Unique Toxins Identified in P. motoro and Fish Venom Comparison
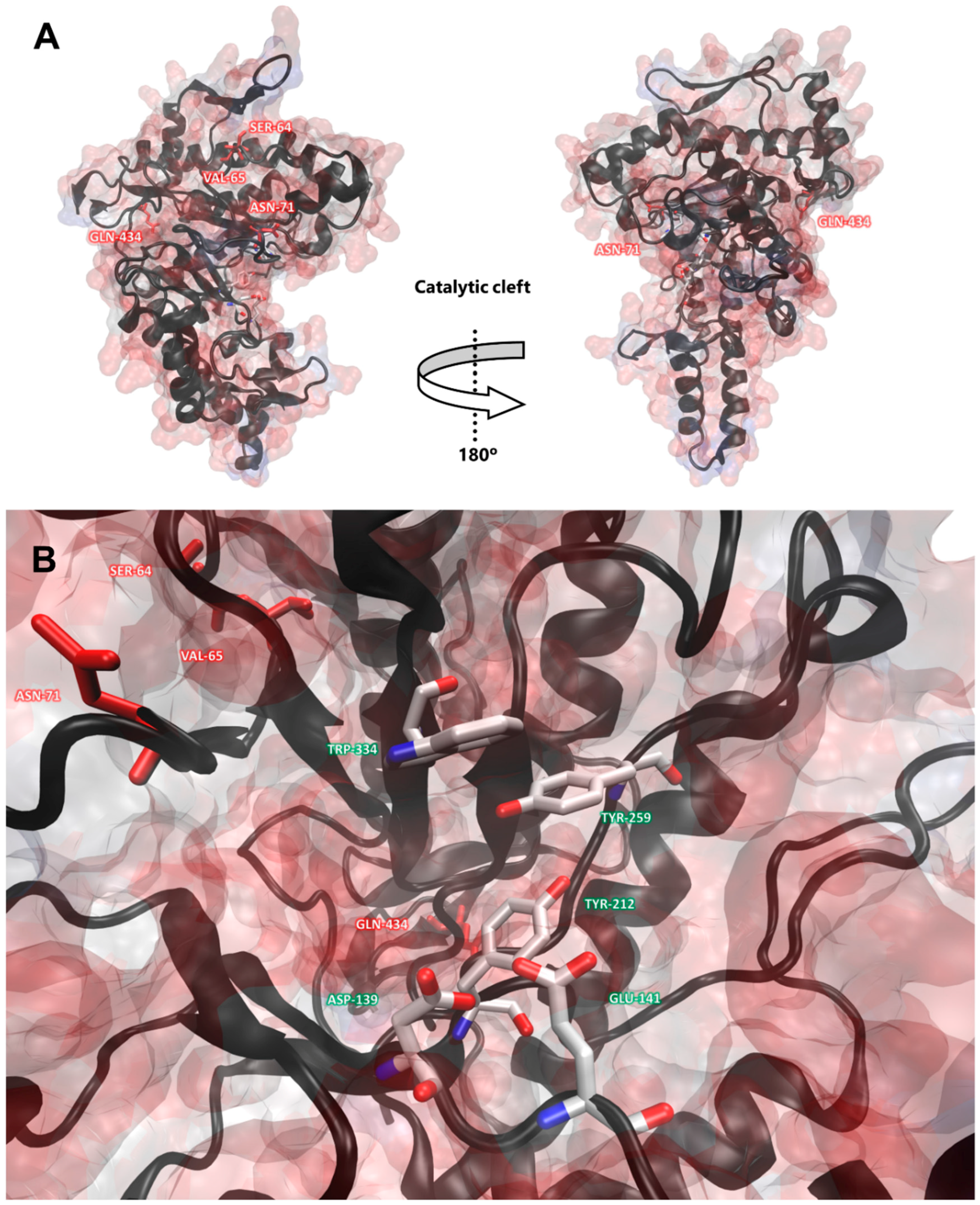
2.5. Hyaluronidase Phylogeny and Selective Pressure
3. Conclusions
4. Materials and Methods
4.1. Fish Collection and Sample Processing
4.2. RNA Extraction and Sequencing
4.3. Data Filtering and De novo Assembly
4.4. Transcript Expression Calculation
4.5. Transcriptome, Functional and Pathway Annotations
4.6. Sequence Alignment and Phylogenetic Analyses
4.7. Selective Pressure Analyses
4.8. Structure Modelling
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Loboda, T.S.; Carvalho, M.R. Systematic revision of the Potamotrygon motoro (Müller & Henle, 1841) species complex in the Paraná-Paraguay basin, with description of two new ocellated species (Chondrichthyes: Myliobatiformes; Potamotrygonidae). Neotrop. Ichthyol. 2013, 11, 693–737. [Google Scholar]
- Carvalho, M.; Lovejoy, N. Morphology and phylogenetic relationships of a remarkable new genus and two new species of Neotropical freshwater stingrays from the Amazon basin (Chondrichthyes: Potamotrygonidae). Zootaxa 2011, 2776, 13–48. [Google Scholar]
- De Carvalho, M.R.; Loboda, T.S.; da Silva, J.P.C.B. A new subfamily, Styracurinae, and new genus, Styracura, for Himantura schmardae (Werner, 1904) and Himantura pacifica (Beebe & Tee-Van, 1941) (Chondrichthyes: Myliobatiformes). Zootaxa 2016, 4175, 21. [Google Scholar] [CrossRef]
- Júlio Júnior, H.F.; Tós, C.D.; Agostinho, Â.A.; Pavanelli, C.S. A massive invasion of fish species after eliminating a natural barrier in the upper rio Paraná basin. Neotrop. Ichthyol. 2009, 7, 709–718. [Google Scholar] [CrossRef]
- Oddone, M.C.; Velasco, G.; Charvet, P. Record of the freshwater stingrays Potamotrygon brachyura and P. motoro (Chondrichthyes, Potamotrygonidae) in the lower Uruguay river, South America. Acta Amaz. 2012, 42, 299–304. [Google Scholar] [CrossRef]
- Barbaro, K.C.; Lira, M.S.; Malta, M.B.; Soares, S.L.; Garrone Neto, D.; Cardoso, J.L.; Santoro, M.L.; Haddad Junior, V. Comparative study on extracts from the tissue covering the stingers of freshwater (Potamotrygon falkneri) and marine (Dasyatis guttata) stingrays. J. Int. Soc. Toxinol. 2007, 50, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Haddad, V., Jr.; Neto, D.G.; de Paula Neto, J.B.; de Luna Marques, F.P.; Barbaro, K.C. Freshwater stingrays: Study of epidemiologic, clinic and therapeutic aspects based on 84 envenomings in humans and some enzymatic activities of the venom. J. Int. Soc. Toxinol. 2004, 43, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Pedroso, C.M.; Jared, C.; Charvet-Almeida, P.; Almeida, M.P.; Garrone Neto, D.; Lira, M.S.; Haddad, V., Jr.; Barbaro, K.C.; Antoniazzi, M.M. Morphological characterization of the venom secretory epidermal cells in the stinger of marine and freshwater stingrays. J. Int. Soc. Toxinol. 2007, 50, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Williamson, J.A.; Burnett, J.W.; Fenner, P.J.; Rifkin, J.F. Venomous and Poisonous Marine Animals: A Medical and Biological Handbook; University of New South Wales Press: Kensington, Australia, 1996. [Google Scholar]
- Junghanss, T.; Bodio, M. Medically important venomous animals: Biology, prevention, first aid, and clinical management. Clin. Infect. Dis. 2006, 43, 1309–1317. [Google Scholar] [CrossRef] [PubMed]
- Barber, G.R.; Swygert, J.S. Necrotizing fasciitis due to Photobacterium damsela in a man lashed by a stingray. N. Engl. J. Med. 2000, 342, 824. [Google Scholar] [CrossRef] [PubMed]
- Domingos, M.O.; Franzolin, M.R.; dos Anjos, M.T.; Franzolin, T.M.; Barbosa Albes, R.C.; de Andrade, G.R.; Lopes, R.J.; Barbaro, K.C. The influence of environmental bacteria in freshwater stingray wound-healing. J. Int. Soc. Toxinol. 2011, 58, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.F.; Girard, R.H.; Rao, D.; Ly, B.T.; Davis, D.P. Stingray envenomation: A retrospective review of clinical presentation and treatment in 119 cases. J. Emerg. Med. 2007, 33, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Diaz, J.H. The evaluation, management, and prevention of stingray injuries in travelers. J. Travel Med. 2008, 15, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Junior, V.H.; Cardoso, J.L.; Neto, D.G. Injuries by marine and freshwater stingrays: History, clinical aspects of the envenomations and current status of a neglected problem in Brazil. J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 16. [Google Scholar] [CrossRef] [PubMed]
- Torrez, P.P.; Quiroga, M.M.; Said, R.; Abati, P.A.; Franca, F.O. Tetanus after envenomations caused by freshwater stingrays. J. Int. Soc. Toxinol. 2015, 97, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Roelants, K.; Norman, J.A. Tentacles of venom: Toxic protein convergence in the Kingdom Animalia. J. Mol. Evol. 2009, 68, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Wuster, W.; Kini, R.M.; Brusic, V.; Khan, A.; Venkataraman, D.; Rooney, A.P. Molecular evolution and phylogeny of elapid snake venom three-finger toxins. J. Mol. Evol. 2003, 57, 110–129. [Google Scholar] [CrossRef] [PubMed]
- Ruder, T.; Sunagar, K.; Undheim, E.A.; Ali, S.A.; Wai, T.C.; Low, D.H.; Jackson, T.N.; King, G.F.; Antunes, A.; Fry, B.G. Molecular phylogeny and evolution of the proteins encoded by coleoid (cuttlefish, octopus, and squid) posterior venom glands. J. Mol. Evol. 2013, 76, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.L.; Wheeler, W.C. Venom Evolution Widespread in Fishes: A Phylogenetic Road Map for the Bioprospecting of Piscine Venoms. J. Hered. 2006, 97, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Undheim, E.A.; Sunagar, K.; Herzig, V.; Kely, L.; Low, D.H.; Jackson, T.N.; Jones, A.; Kurniawan, N.; King, G.F.; Ali, S.A.; et al. A proteomics and transcriptomics investigation of the venom from the barychelid spider Trittame loki (brush-foot trapdoor). Toxins 2013, 5, 2488–2503. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, K.; Undheim, E.A.; Chan, A.H.; Koludarov, I.; Munoz-Gomez, S.A.; Antunes, A.; Fry, B.G. Evolution stings: The origin and diversification of scorpion toxin peptide scaffolds. Toxins 2013, 5, 2456–2487. [Google Scholar] [CrossRef] [PubMed]
- Freitas-de-Sousa, L.A.; Amazonas, D.R.; Sousa, L.F.; Sant’Anna, S.S.; Nishiyama, M.Y.; Serrano, S.M.T.; Junqueira-de-Azevedo, I.L.M.; Chalkidis, H.M.; Moura-da-Silva, A.M.; Mourão, R.H.V. Comparison of venoms from wild and long-term captive Bothrops atrox snakes and characterization of Batroxrhagin, the predominant class PIII metalloproteinase from the venom of this species. Biochimie 2015, 118, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, K.; Undheim, E.A.B.; Scheib, H.; Gren, E.C.K.; Cochran, C.; Person, C.E.; Koludarov, I.; Kelln, W.; Hayes, W.K.; King, G.F.; et al. Intraspecific venom variation in the medically significant Southern Pacific Rattlesnake (Crotalus oreganus helleri): Biodiscovery, clinical and evolutionary implications. J. Proteom. 2014, 99, 68–83. [Google Scholar] [CrossRef] [PubMed]
- Conceição, K.; Konno, K.; Melo, R.L.; Marques, E.E.; Hiruma-Lima, C.A.; Lima, C.; Richardson, M.; Pimenta, D.C.; Lopes-Ferreira, M. Orpotrin: A novel vasoconstrictor peptide from the venom of the Brazilian Stingray Potamotrygon gr. orbignyi. Peptides 2006, 27, 3039–3046. [Google Scholar] [CrossRef] [PubMed]
- Conceição, K.; Santos, J.M.; Bruni, F.M.; Klitzke, C.F.; Marques, E.E.; Borges, M.H.; Melo, R.L.; Fernandez, J.H.; Lopes-Ferreira, M. Characterization of a new bioactive peptide from Potamotrygon gr. orbignyi freshwater stingray venom. Peptides 2009, 30, 2191–2199. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, M.R.; da Silva, N.J.; Ulhoa, C.J. A hyaluronidase from Potamotrygon motoro (freshwater stingrays) venom: Isolation and characterization. J. Int. Soc. Toxinol. 2008, 51, 1060–1067. [Google Scholar] [CrossRef] [PubMed]
- Júnior, N.G.d.O.; Fernandes, G.d.R.; Cardoso, M.H.; Costa, F.F.; Cândido, E.d.S.; Neto, D.G.; Mortari, M.R.; Schwartz, E.F.; Franco, O.L.; de Alencar, S.A. Venom gland transcriptome analyses of two freshwater stingrays (Myliobatiformes: Potamotrygonidae) from Brazil. Sci. Rep. 2016, 6, 21935. [Google Scholar] [CrossRef] [PubMed]
- Baumann, K.; Casewell, N.R.; Ali, S.A.; Jackson, T.N.; Vetter, I.; Dobson, J.S.; Cutmore, S.C.; Nouwens, A.; Lavergne, V.; Fry, B.G. A ray of venom: Combined proteomic and transcriptomic investigation of fish venom composition using barb tissue from the blue-spotted stingray (Crotalus adamanteus). J. Proteom. 2014, 109, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.; Kogan, G.; Jedrzejas, M.J.; Šoltés, L. The many ways to cleave hyaluronan. Biotechnol. Adv. 2007, 25, 537–557. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, D158–D169. [CrossRef] [PubMed]
- Jungo, F.; Bougueleret, L.; Xenarios, I.; Poux, S. The UniProtKB/Swiss-Prot Tox-Prot program: A central hub of integrated venom protein data. J. Int. Soc. Toxinol. 2012, 60, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2016, 44, D67–D72. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Sato, Y.; Furumichi, M.; Tanabe, M. KEGG for integration and interpretation of large-scale molecular data sets. Nucleic Acids Res. 2012, 40, D109–114. [Google Scholar] [CrossRef] [PubMed]
- Ben Bacha, A.; Abid, I.; Horchani, H.; Mejdoub, H. Enzymatic properties of stingray Dasyatis pastinaca group V, IIA and IB phospholipases A(2): A comparative study. Int. J. Biol. Macromol. 2013, 62, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Uechi, G.; Toma, H.; Arakawa, T.; Sato, Y. Molecular characterization on the genome structure of hemolysin toxin isoforms isolated from sea anemone Actineria villosa and Phyllodiscus semoni. J. Int. Soc. Toxinol. 2010, 56, 1470–1476. [Google Scholar] [CrossRef] [PubMed]
- Imperial, J.S.; Kantor, Y.; Watkins, M.; Heralde, F.M., 3rd; Stevenson, B.; Chen, P.; Hansson, K.; Stenflo, J.; Ownby, J.P.; Bouchet, P.; et al. Venomous auger snail Hastula (Impages) hectica (Linnaeus, 1758): Molecular phylogeny, foregut anatomy and comparative toxinology. J. Exp. Zool. Part B Mol. Dev. Evol. 2007, 308, 744–756. [Google Scholar] [CrossRef] [PubMed]
- Ishida, M.; Minagawa, S.; Miyauchi, K.; Shimakura, K.; Nagashima, Y.; Shiomi, K. Amino Acid Sequences of Kunitz-type Protease Inhibitors from the Sea Anemone Actinia Equina. Fish. Sci. 1997, 63, 794–798. [Google Scholar] [CrossRef]
- Ziegman, R.; Alewood, P. Bioactive Components in Fish Venoms. Toxins 2015, 7, 1497. [Google Scholar] [CrossRef] [PubMed]
- Mitra, J.; Bhattacharyya, D. Phosphodiesterase from Daboia russelli russelli venom: Purification, partial characterization and inhibition of platelet aggregation. J. Int. Soc. Toxinol. 2014, 88, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Belli, S.I.; Goding, J.W. Biochemical characterization of human PC-1, an enzyme possessing alkaline phosphodiesterase I and nucleotide pyrophosphatase activities. Eur. J. Biochem. 1994, 226, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Margres, M.J.; McGivern, J.J.; Wray, K.P.; Seavy, M.; Calvin, K.; Rokyta, D.R. Linking the transcriptome and proteome to characterize the venom of the eastern diamondback rattlesnake (Crotalus adamanteus). J. Proteom. 2014, 96, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.R.; Visser, J.C.; Baumann, K.; Dobson, J.; Han, H.; Kuruppu, S.; Morgan, M.; Romilio, A.; Weisbecker, V.; Mardon, K.; et al. The Evolution of Fangs, Venom, and Mimicry Systems in Blenny Fishes. Curr. Biol. 2017, 27, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- Ng, H.C.; Ranganathan, S.; Chua, K.L.; Khoo, H.E. Cloning and molecular characterization of the first aquatic hyaluronidase, SFHYA1, from the venom of stonefish (Synanceja horrida). Gene 2005, 346, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Cherr, G.N.; Yudin, A.I.; Overstreet, J.W. The dual functions of GPI-anchored PH-20: Hyaluronidase and intracellular signaling. J. Int. Soc. Matrix Biol. 2001, 20, 515–525. [Google Scholar] [CrossRef]
- Smith, W.L.; Stern, J.H.; Girard, M.G.; Davis, M.P. Evolution of Venomous Cartilaginous and Ray-Finned Fishes. Integr. Comp. Biol. 2016, 56, 950–961. [Google Scholar] [CrossRef] [PubMed]
- Reitinger, S.; Laschober, G.T.; Fehrer, C.; Greiderer, B.; Lepperdinger, G. Mouse testicular hyaluronidase-like proteins SPAM1 and HYAL5 but not HYALP1 degrade hyaluronan. Biochem. J. 2007, 401, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Pond, S.L.K.; Frost, S.D.W.; Muse, S.V. HyPhy: Hypothesis testing using phylogenies. Bioinformatics 2005, 21, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Kosakovsky Pond, S.L.; Frost, S.D.W. Not So Different After All: A Comparison of Methods for Detecting Amino Acid Sites Under Selection. Mol. Biol. Evol. 2005, 22, 1208–1222. [Google Scholar] [CrossRef] [PubMed]
- Murrell, B.; Moola, S.; Mabona, A.; Weighill, T.; Sheward, D.; Kosakovsky Pond, S.L.; Scheffler, K. FUBAR: A Fast, Unconstrained Bayesian AppRoximation for Inferring Selection. Mol. Biol. Evol. 2013, 30, 1196–1205. [Google Scholar] [CrossRef] [PubMed]
- Murrell, B.; Wertheim, J.O.; Moola, S.; Weighill, T.; Scheffler, K.; Kosakovsky Pond, S.L. Detecting Individual Sites Subject to Episodic Diversifying Selection. PLoS Genet. 2012, 8, e1002764. [Google Scholar] [CrossRef] [PubMed]
- Ridout, K.E.; Dixon, C.J.; Filatov, D.A. Positive Selection Differs between Protein Secondary Structure Elements in Drosophila. Genome Biol. Evol. 2010, 2, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD—Visual Molecular Dynamics. J. Mol. Gr. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Vonk, F.J.; Casewell, N.R.; Henkel, C.V.; Heimberg, A.M.; Jansen, H.J.; McCleary, R.J.R.; Kerkkamp, H.M.E.; Vos, R.A.; Guerreiro, I.; Calvete, J.J.; et al. The king cobra genome reveals dynamic gene evolution and adaptation in the snake venom system. Proc. Natl. Acad. Sci. USA 2013, 110, 20651–20656. [Google Scholar] [CrossRef] [PubMed]
- Kemparaju, K.; Girish, K.S. Snake venom hyaluronidase: A therapeutic target. Cell Biochem. Funct. 2006, 24, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, N.J.; Clementino Ferreira, K.R.; Leite Pinto, R.N.; Aird, S.D. A Severe Accident Caused by an Ocellate River Stingray (Potamotrygon motoro) in Central Brazil: How Well Do We Really Understand Stingray Venom Chemistry, Envenomation, and Therapeutics? Toxins 2015, 7, 2272–2288. [Google Scholar] [CrossRef] [PubMed]
- Monteiro-dos-Santos, J.; Conceicao, K.; Seibert, C.S.; Marques, E.E.; Silva, P.I., Jr.; Soares, A.B.; Lima, C.; Lopes-Ferreira, M. Studies on pharmacological properties of mucus and sting venom of Potamotrygon cf. henlei. Int. Immunopharmacol. 2011, 11, 1368–1377. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef] [PubMed]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B.; et al. TIGR Gene Indices clustering tools (TGICL): A software system for fast clustering of large EST datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Punta, M.; Coggill, P.C.; Eberhardt, R.Y.; Mistry, J.; Tate, J.; Boursnell, C.; Pang, N.; Forslund, K.; Ceric, G.; Clements, J.; et al. The Pfam protein families database. Nucleic Acids Res. 2012, 40, D290–D301. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Fang, L.; Zheng, H.; Zhang, Y.; Chen, J.; Zhang, Z.; Wang, J.; Li, S.; Li, R.; Bolund, L.; et al. WEGO: A web tool for plotting GO annotations. Nucleic Acids Res. 2006, 34, W293–W297. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView Version 4: A Multiplatform Graphical User Interface for Sequence Alignment and Phylogenetic Tree Building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Da Fonseca, R.R.; Antunes, A.; Melo, A.; Ramos, M.J. Structural divergence and adaptive evolution in mammalian cytochromes P450 2C. Gene 2007, 387, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Abascal, F.; Zardoya, R.; Posada, D. ProtTest: Selection of best-fit models of protein evolution. Bioinformatics 2005, 21, 2104–2105. [Google Scholar] [CrossRef] [PubMed]
- Da Fonseca, R.R.; Johnson, W.E.; O’Brien, S.J.; Vasconcelos, V.; Antunes, A. Molecular evolution and the role of oxidative stress in the expansion and functional diversification of cytosolic glutathione transferases. BMC Evol. Biol. 2010, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Anisimova, M.; Nielsen, R.; Yang, Z. Effect of recombination on the accuracy of the likelihood method for detecting positive selection at amino acid sites. Genetics 2003, 164, 1229–1236. [Google Scholar] [PubMed]
- Kosakovsky Pond, S.L.; Posada, D.; Gravenor, M.B.; Woelk, C.H.; Frost, S.D.W. Automated Phylogenetic Detection of Recombination Using a Genetic Algorithm. Mol. Biol. Evol. 2006, 23, 1891–1901. [Google Scholar] [CrossRef] [PubMed]
- Pineda, S.S.; Sollod, B.L.; Wilson, D.; Darling, A.; Sunagar, K.; Undheim, E.A.B.; Kely, L.; Antunes, A.; Fry, B.G.; King, G.F. Diversification of a single ancestral gene into a successful toxin superfamily in highly venomous Australian funnel-web spiders. BMC Genom. 2014, 15, 177. [Google Scholar] [CrossRef] [PubMed]
- Low, D.H.; Sunagar, K.; Undheim, E.A.; Ali, S.A.; Alagon, A.C.; Ruder, T.; Jackson, T.N.; Pineda Gonzalez, S.; King, G.F.; Jones, A.; et al. Dracula’s children: Molecular evolution of vampire bat venom. J. Proteom. 2013, 89, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Undheim, E.A.B.; Ali, S.A.; Jackson, T.N.W.; Debono, J.; Scheib, H.; Ruder, T.; Morgenstern, D.; Cadwallader, L.; Whitehead, D.; et al. Squeezers and leaf-cutters: Differential diversification and degeneration of the venom system in toxicoferan reptiles. Mol. Cell. Proteom. 2013, 12, 1881–1899. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, K.; Fry, B.G.; Jackson, T.N.W.; Casewell, N.R.; Undheim, E.A.B.; Vidal, N.; Ali, S.A.; King, G.F.; Vasudevan, K.; Vasconcelos, V.; et al. Molecular Evolution of Vertebrate Neurotrophins: Co-Option of the Highly Conserved Nerve Growth Factor Gene into the Advanced Snake Venom Arsenalf. PLoS ONE 2013, 8, e81827. [Google Scholar] [CrossRef]
- Pereira, S.R.; Vasconcelos, V.M.; Antunes, A. Computational study of the covalent bonding of microcystins to cysteine residues—A reaction involved in the inhibition of the PPP family of protein phosphatases. FEBS J. 2013, 280, 674–680. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.R.; Vasconcelos, V.M.; Antunes, A. The phosphoprotein phosphatase family of Ser/Thr phosphatases as principal targets of naturally occurring toxins. Crit. Rev. Toxicol. 2011, 41, 83–110. [Google Scholar] [CrossRef] [PubMed]
- Furnham, N.; Holliday, G.L.; de Beer, T.A.; Jacobsen, J.O.; Pearson, W.R.; Thornton, J.M. The Catalytic Site Atlas 2.0: Cataloging catalytic sites and residues identified in enzymes. Nucleic Acids Res. 2014, 42, D485–D489. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process | Value |
|---|---|
| Number of raw reads | 38,674,474 |
| Raw data (bp) | 3,480,702,660 |
| Read length (bp) | 90 |
| Number of high-quality reads | 35,977,224 |
| Average high-quality read length (bp) | 80 |
| Number of contigs | 140,078 |
| Number of contigs ≥ 1 FPKM | 107,129 |
| Number of contigs ≥ 1 FPKM and containing coding sequences | 27,032 |
| Contigs (bp) | 69,861,618 |
| N50 | 690 |
| Average contig length (bp) | 498 |
| Min. contig length (bp) | 174 |
| Max. contig length (bp) | 15,040 |
| Database | Number of Annotated Contigs |
|---|---|
| NCBI | 20,967 (84.77%) |
| SwissProt | 19,851 (80.26%) |
| NCBI and SwissProt | 19,398 (78.43%) |
| NCBI or SwissProt | 21,420 (86.60%) |
| ToxProt | 418 (1.69%) |
| PFAM | 16,823 (68.02%) |
| GO | 19,194 (77.60%) |
| KEGG | 17,572 (71.04%) |
| Potamotrygon motoro | Potamotrygon amandae | Potamotrygon falkneri | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Transcript | Protein | Uniprot Accession | FPKM | Protein | Uniprot Accession | FPKM | Protein | Uniprot Accession | FPKM |
| TR1637 | SE-cephalotoxin | B2DCR8 | 42139.08 | Hyaluronidase | J3S820 | 22201.33 | Hyaluronidase | J3S820 | 9488.18 |
| TR45956 | Hyaluronidase | J3S820 | 7271.53 | Translationally controlled tumor protein homolog | U3EQ60 | 1225.86 | Hemolytic toxin Avt-1 | Q5R231 | 757.19 |
| TR7202 | SE-cephalotoxin | B2DCR8 | 489.97 | Cysteine-rich venom protein latisemin | Q8JI38 | 891.77 | Translationally controlled tumor protein homolog | U3EQ60 | 734.38 |
| TR53378 | DELTA-alicitoxin-Pse1b | P0DL56 | 399.42 | Venom allergen 5 | P81656 | 854.22 | Calglandulin | Q3SB11 | 644.06 |
| TR15069 | Cystatin-2 | J3SE80 | 373.92 | Calglandulin | Q8AY75 | 835.98 | Putative Kunitz-type serine protease inhibitor | B2BS84 | 450.57 |
| TR10238 | Augerpeptide hhe53 | P0CI21 | 324.45 | Cystatin-2 | J3SE80 | 629.15 | Peroxiredoxin-4 | P0CV91 | 397.49 |
| TR3474 | Translationally controlled tumor protein homolog | U3EQ60 | 315.67 | Hemolytic toxin Avt-1 | Q5R231 | 401.11 | Cysteine-rich venom protein 1 | Q8T0W5 | 386.31 |
| TR112682 | Calglandulin | Q3SB11 | 202.31 | Putative Kunitz-type serine protease inhibitor | B2BS84 | 354.6 | Venom allergen 5 | P81656 | 357.2 |
| TR24275 | Putative Kunitz-type serine protease inhibitor | B2BS84 | 198.6 | Peroxiredoxin-4 | P0CV91 | 304.79 | Calglandulin | Q3SB11 | 318.87 |
| TR20805 | Venom allergen 5 | P81656 | 174.48 | Cysteine-rich venom protein 1 | Q8T0W5 | 211.07 | Cysteine-rich venom protein latisemin | Q8JI38 | 299.5 |
| TR20804 | Venom allergen 5.02 | P35782 | 165.85 | Alpha-latroinsectotoxin-Lt1a | Q02989 | 206.6 | Vespryn | Q2XXL4 | 172.92 |
| TR14068 | PI-actitoxin-Aeq3a | P0DMW6 | 158.3 | Kunitz-type serine protease inhibitor bitisilin-3 | Q6T269 | 141.02 | Venom serine protease 34 | Q8MQS8 | 156.83 |
| TR15070 | Cystatin-2 | J3SE80 | 146.47 | Analgesic polypeptide HC3 | C0HJF3 | 121.14 | Calglandulin | Q8AY75 | 150.86 |
| TR86455 | Insulin-like growth factor-binding protein-related protein 1 | G4V4G1 | 142.04 | Kunitz-type serine protease inhibitor kappa-theraphotoxin-Hh1a | P68425 | 117.78 | Alpha-latrocrustotoxin-Lt1a | Q9XZC0 | 150.43 |
| TR110388 | Calglandulin | Q3SB11 | 128.78 | Venom prothrombin activator porpharin-D | Q58L93 | 104.65 | Alpha-latrotoxin-Lh1a | G0LXV8 | 150.2 |
| TR112679 | Calglandulin | Q3SB11 | 126.9 | Zinc metalloproteinase-disintegrin-like BmMP | A8QL49 | 101.89 | Zinc metalloproteinase-disintegrin-like BmMP | A8QL49 | 143.04 |
| TR113284 | Cysteine-rich venom protein 1 | Q8T0W5 | 82.72 | Acidic phospholipase A2 | Q9DF56 | 100.95 | Cystatin-2 | J3SE80 | 140.7 |
| TR67254 | Putative Kunitz-type serine protease inhibitor | B2BS84 | 73.96 | Vespryn | Q2XXL4 | 95.12 | Venom protease | Q7M4I3 | 118.09 |
| TR9752 | Zinc metalloproteinase-disintegrin-like BmMP | A8QL49 | 69.87 | Insulin-like growth factor-binding protein-related protein 1 | G4V4G1 | 93.55 | Alpha-latroinsectotoxin-Lt1a | Q02989 | 114.49 |
| TR1967 | Alpha-latrocrustotoxin-Lt1a | Q9XZC0 | 68.33 | Delta-latroinsectotoxin-Lt1a | Q25338 | 87.69 | Kunitz-type serine protease inhibitor HNTX-852 | P0DJ69 | 108.68 |
| TR119403 | Alpha-latrocrustotoxin-Lt1a | Q9XZC0 | 68.3 | Ohanin | P83234 | 86.84 | Venom prothrombin activator porpharin-D | Q58L93 | 107.52 |
| TR110386 | Calglandulin | Q3SB11 | 67.47 | Venom prothrombin activator vestarin-D2 | A6MFK8 | 71.33 | Analgesic polypeptide HC3 | C0HJF3 | 106.37 |
| TR7292 | DELTA-thalatoxin-Avl1a | Q5R231 | 66.92 | Blarina toxin | Q76B45 | 70.84 | Snake venom metalloprotease inhibitor | A8YPR9 | 86.64 |
| TR53095 | Peroxiredoxin-4 | P0CV91 | 64.91 | Venom protease | Q7M4I3 | 59.88 | Kunitz-type serine protease inhibitor bitisilin-3 | Q6T269 | 81.19 |
| Venom Components | |
|---|---|
| Bony Fish [41] | Marine Stingray [29] |
| Trachynilysin (TLY) | 60S acidic ribosomal protein |
| Stonustoxin (SNTX) | ATP synthase |
| Verrucotoxin (VTX) | Coronin |
| Neoverrucotoxin (neoVTX) | Cystatin |
| Cardioleputin | Cytochrome C |
| Nocitoxin | Ferritin |
| Karatoxin | Galectin |
| Sp-CTx | Ganglioside GM2 activator |
| Plumieribetin | Glutathione S-transferase mu |
| SP-CL 1-5 | Hemoglobin subunit alpha |
| Dracotoxin | Leukocyte elastase inhibitor |
| Trachinine | Nucleoside diphosphate kinase |
| SA-HT | Peroxiredoxin 6 |
| TmC4-47.2 | Transaldolase |
| Nattectin | Type III intermediate filament |
| Toxin-PC | Voltage-dependent anion channel |
| Wap65 | - |
| Natterin | - |
| Hyaluronidase | - |
| Phospholipase A2 [45] | - |
| Proenkephalin [45] | - |
| Neuropeptide Y [45] | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, F.; Huang, Y.; Yang, V.; Mu, X.; Shi, Q.; Antunes, A. Transcriptomic Characterization of the South American Freshwater Stingray Potamotrygon motoro Venom Apparatus. Toxins 2018, 10, 544. https://doi.org/10.3390/toxins10120544
Silva F, Huang Y, Yang V, Mu X, Shi Q, Antunes A. Transcriptomic Characterization of the South American Freshwater Stingray Potamotrygon motoro Venom Apparatus. Toxins. 2018; 10(12):544. https://doi.org/10.3390/toxins10120544
Chicago/Turabian StyleSilva, Filipe, Yu Huang, Vítor Yang, Xidong Mu, Qiong Shi, and Agostinho Antunes. 2018. "Transcriptomic Characterization of the South American Freshwater Stingray Potamotrygon motoro Venom Apparatus" Toxins 10, no. 12: 544. https://doi.org/10.3390/toxins10120544
APA StyleSilva, F., Huang, Y., Yang, V., Mu, X., Shi, Q., & Antunes, A. (2018). Transcriptomic Characterization of the South American Freshwater Stingray Potamotrygon motoro Venom Apparatus. Toxins, 10(12), 544. https://doi.org/10.3390/toxins10120544