Panton-Valentine Leucocidin Proves Direct Neuronal Targeting and Its Early Neuronal and Glial Impacts a Rabbit Retinal Explant Model

 ,
,
Abstract
1. Introduction
2. Results
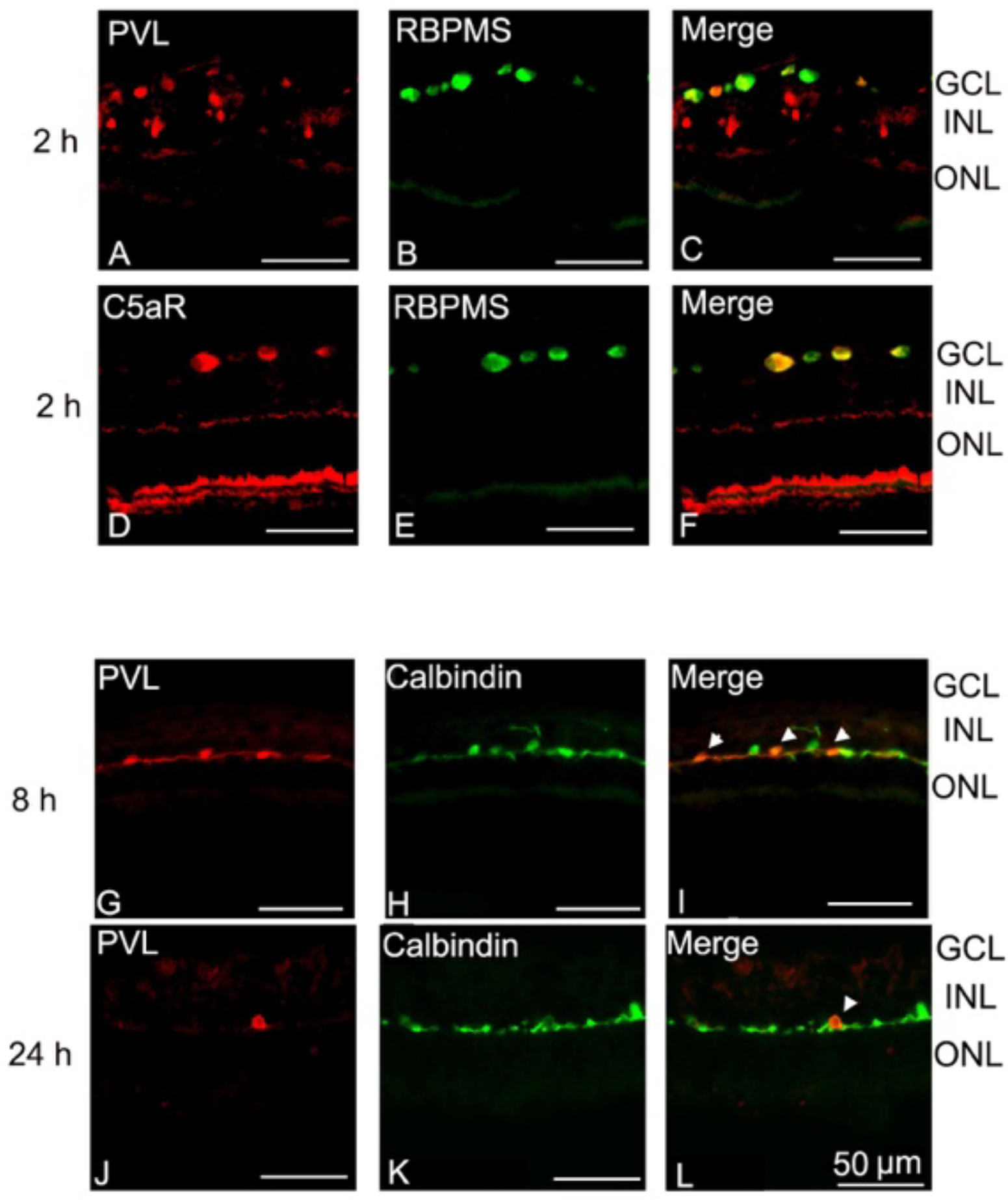
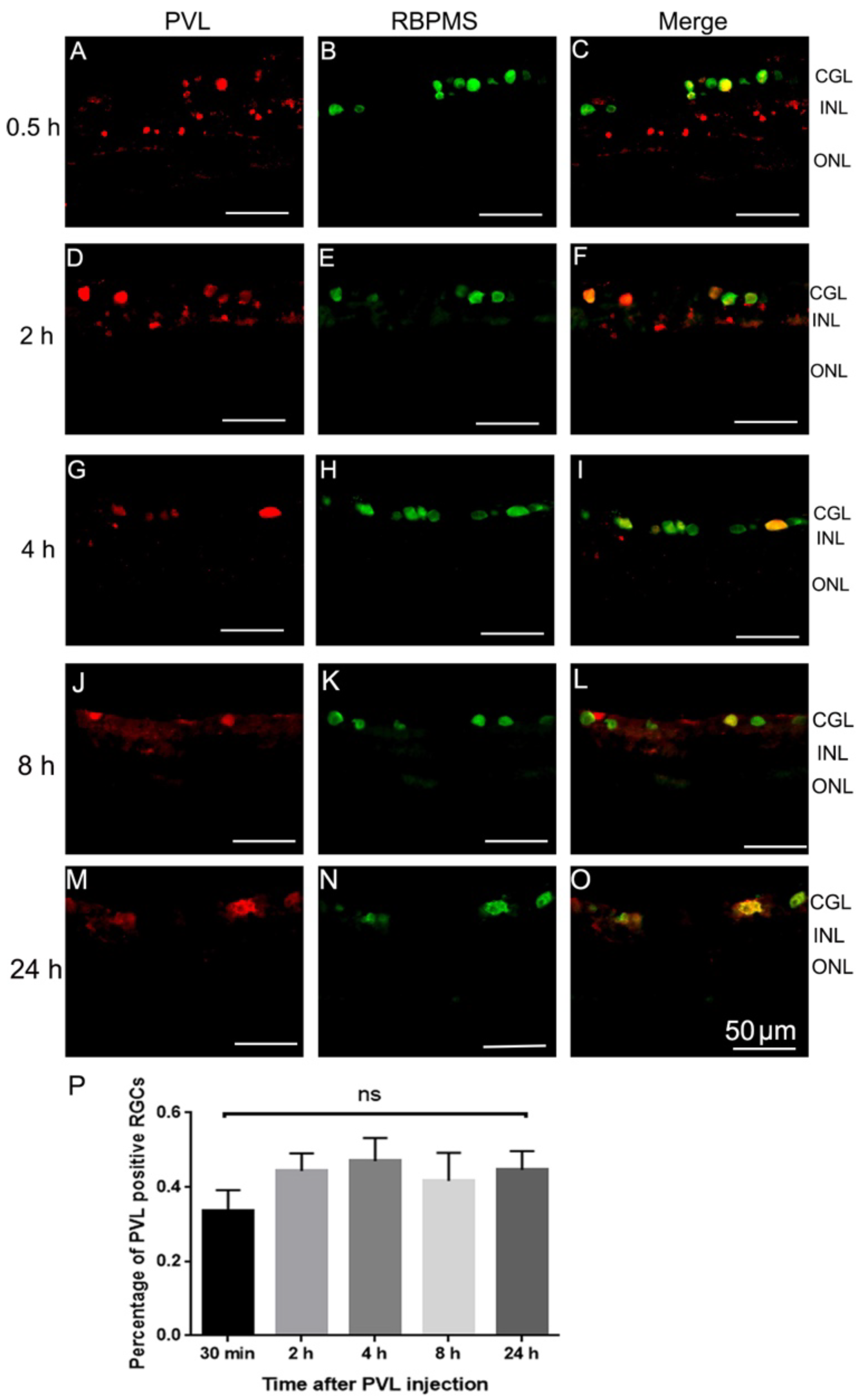
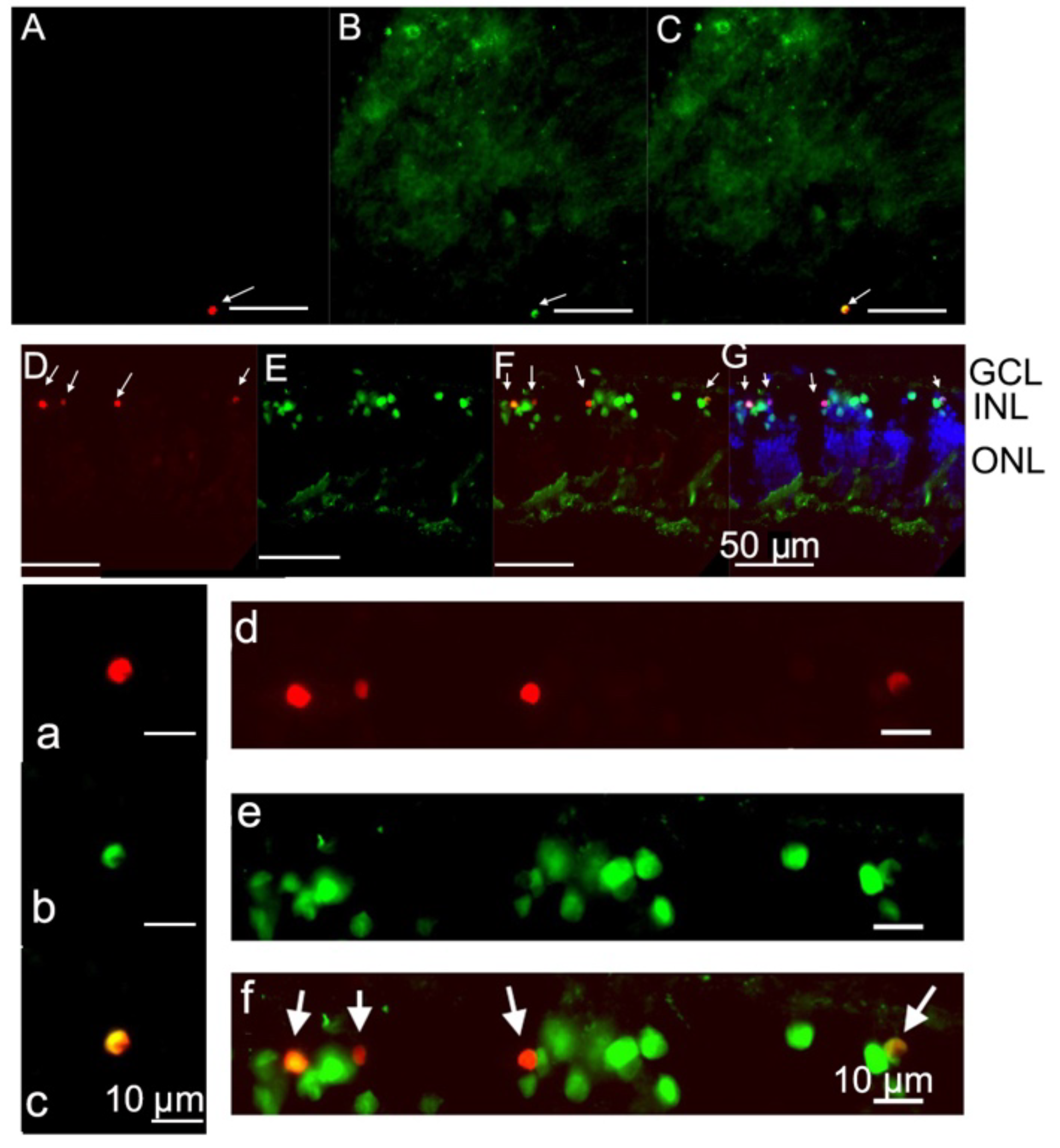
2.1. PVL Co-Localized with RGCs and Horizontal Cells.
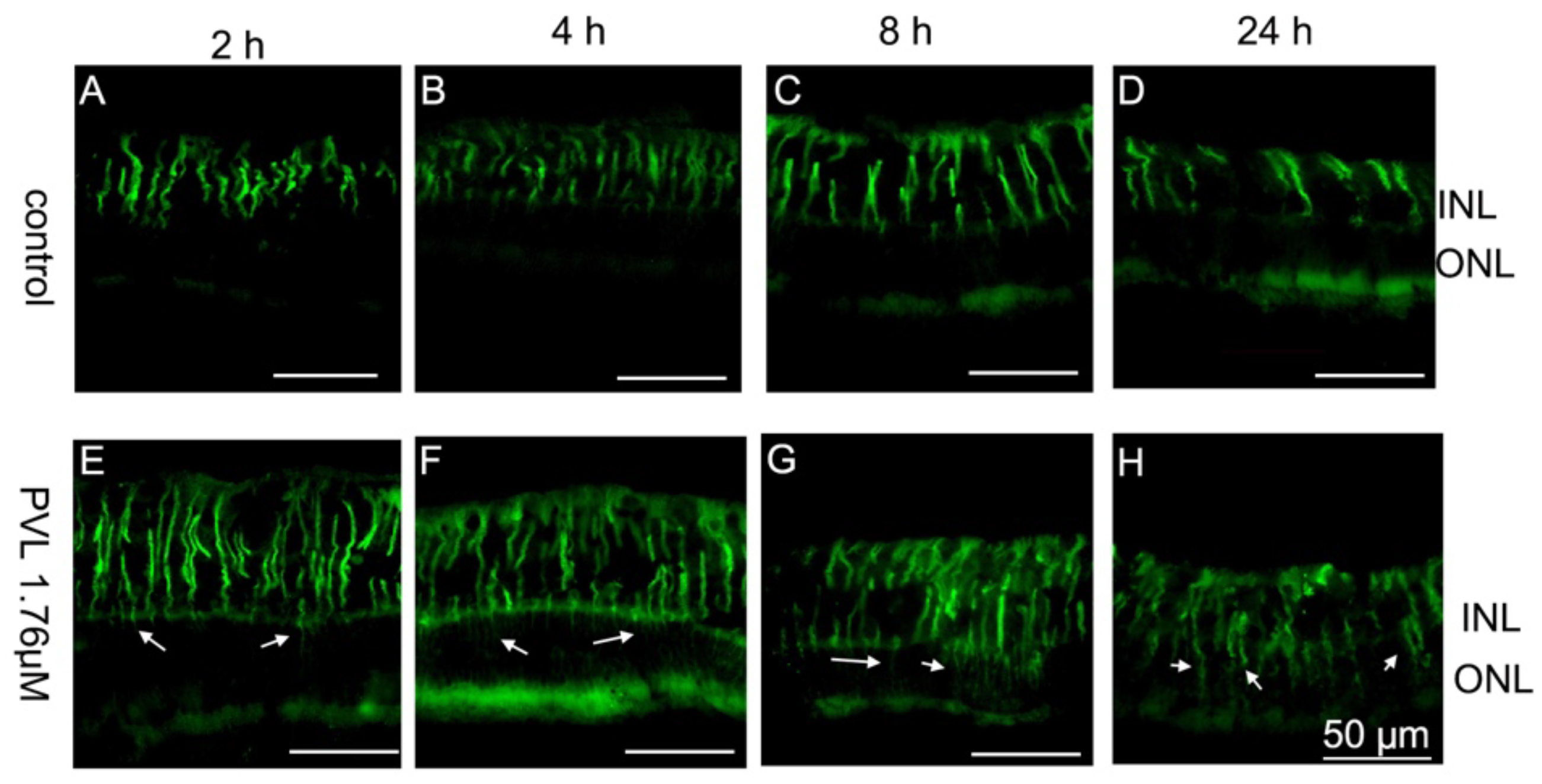
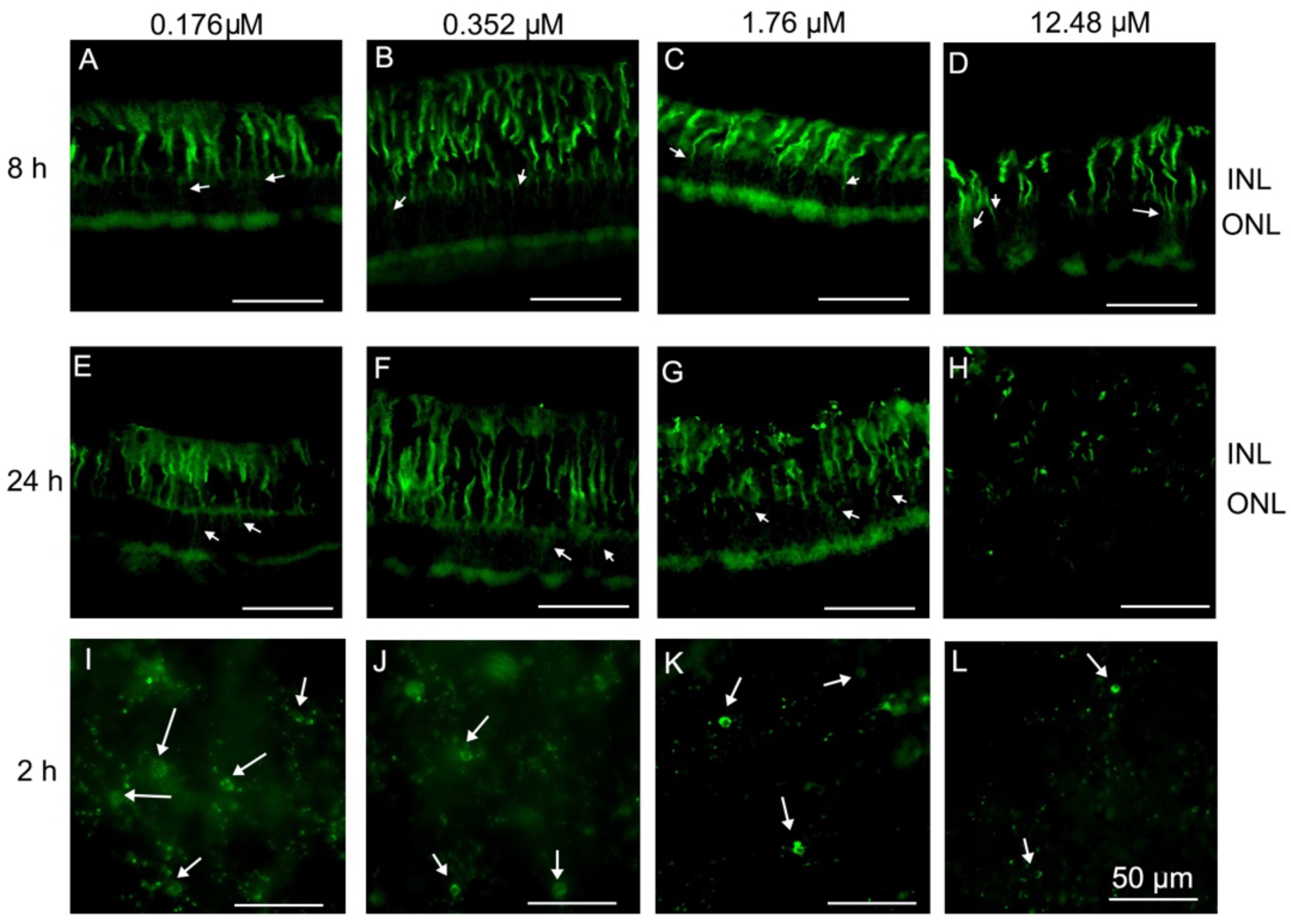
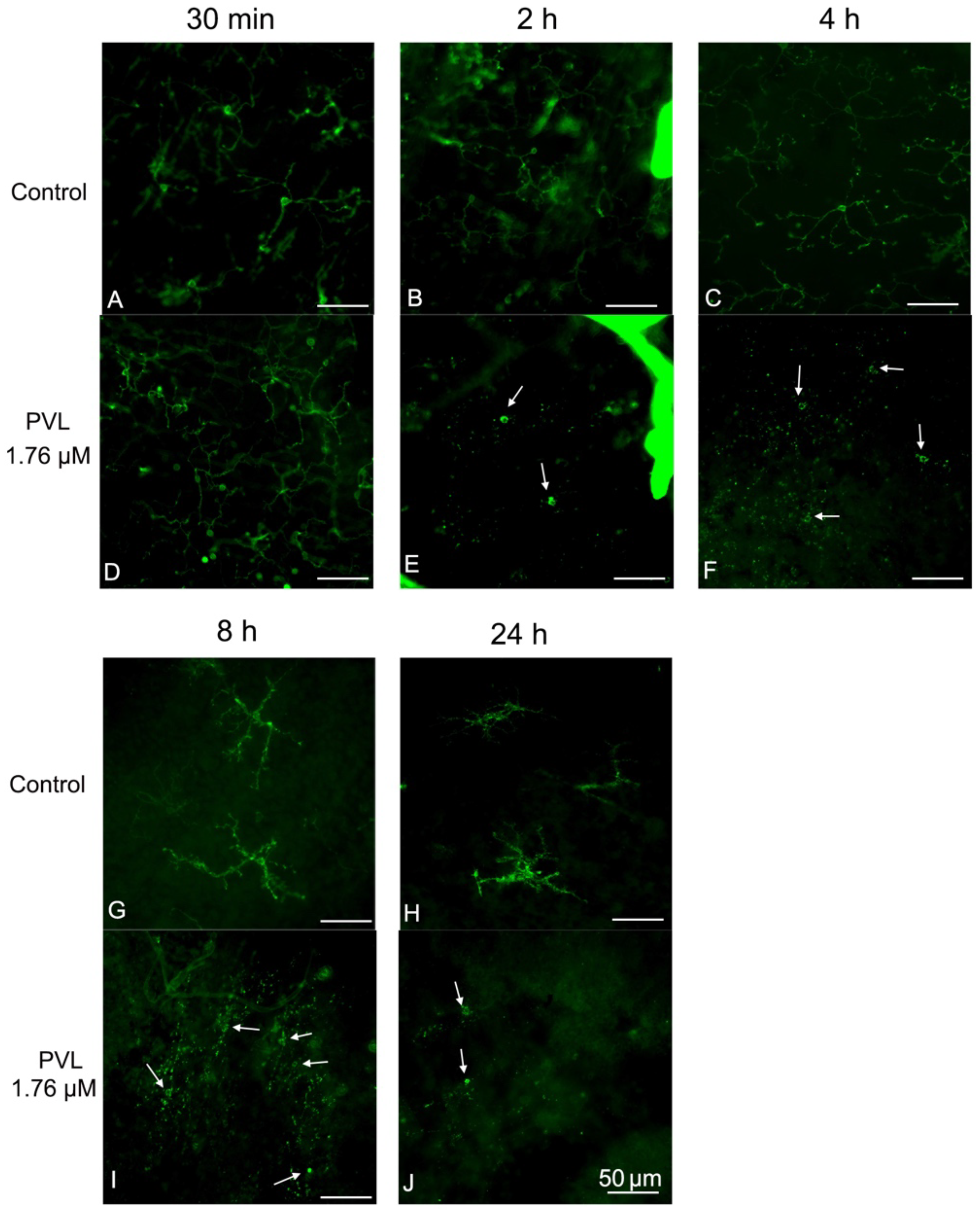
2.2. Müller and Microglial Cells Were Dramatically Activated Early by PVL in a Concentration- and Time-Dependent Manner
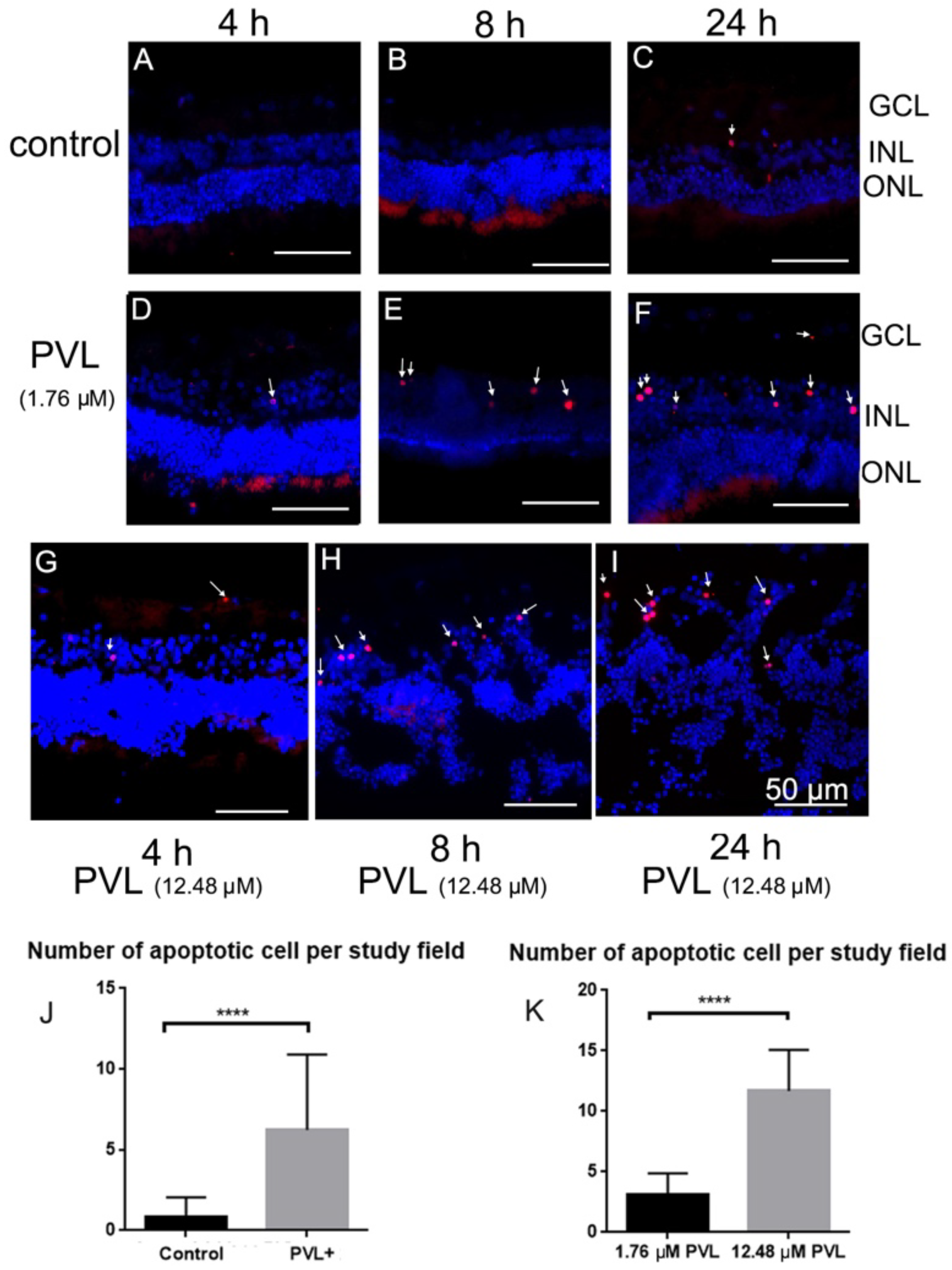
2.3. Some Microglial and Amacrine Cells Underwent Apoptosis in PVL-Treated Explants.
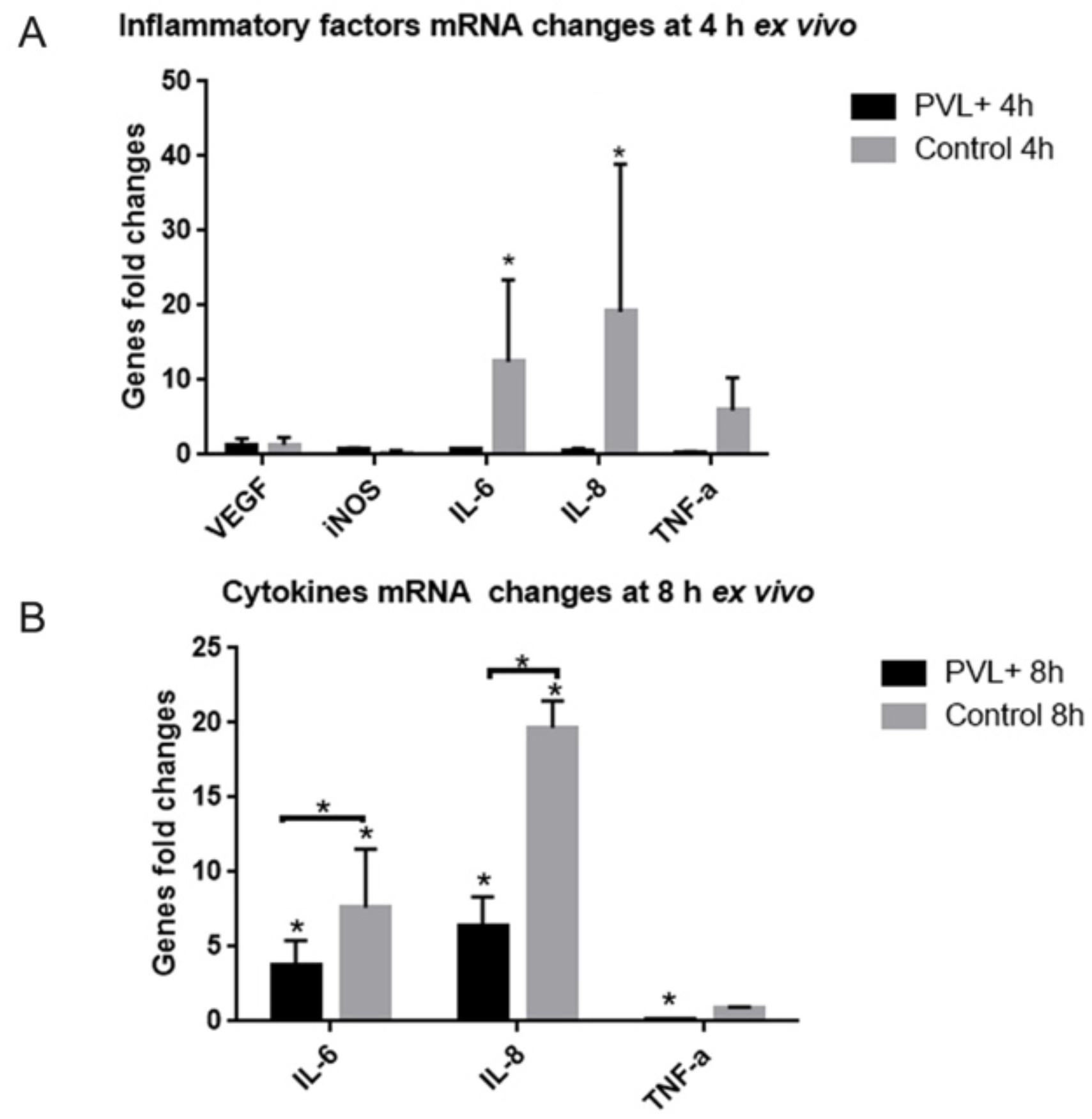
2.4. In PVL-Treated Explants, the Expression of Some Inflammatory Factors Was Not as Increased as in Control Explants
3. Discussion
4. Materials and Methods
4.1. PVL Purification
4.2. Retinal Explant Preparation and Organotypic Culture
4.3. PVL-Treated Explants and Control Explants
4.4. Tissue Processing
4.5. Fluorescent Immunostaining
4.6. Cell Counting
4.7. RNA Extraction
4.8. Real-Time RT-qPCR
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Callegan, M.C.; Engelbert, M.; Jett, B.D.; Gilmore, M.S. Bacterial endophthalmitis: Epidemiology, therapeutics, and bacterium-host interactions. Clin. Microbiol. Rev. 2002, 15, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Callegan, M.C.; Gilmore, M.S.; Gregory, M.; Ramadan, R.T.; Wiskur, B.J.; Moyer, A.L.; Hunt, J.J.; Novosad, B.D. Bacterial endophthalmitis: Therapeutic challenges and host-pathogen interactions. Prog. Retin. Eye Res. 2007, 26, 189–203. [Google Scholar] [CrossRef] [PubMed]
- Vincenot, F.; Saleh, M.; Prévost, G. Les facteurs de virulence de Staphylococcus aureus. Revue Francophone des Lab. 2008, 2008, 61–69. [Google Scholar] [CrossRef]
- Spaan, A.N.; van Strijp, J.A.G.; Torres, V.J. Leukocidins: staphylococcal bi-component pore-forming toxins find their receptors. Nat. Rev. Microbiol. 2017, 15, 435–447. [Google Scholar]
- Rooijakkers, S.H.; Ruyken, M.; van Roon, J.; van Kessel, K.P.; van Strijp, J.A.; van Wamel, W.J. Early expression of SCIN and CHIPS drives instant immune evasion by Staphylococcus aureus. Cell. Microbiol. 2006, 8, 1282–1293. [Google Scholar]
- Spaan, A.N.; Schiepers, A.; de Haas, C.J.; van Hooijdonk, D.D.; Badiou, C.; Contamin, H.; Vandenesch, F.; Lina, G.; Gerard, N.P.; Gerard, C.; et al. Differential Interaction of the Staphylococcal Toxins Panton-Valentine Leukocidin and gamma-Hemolysin CB with Human C5a Receptors. J. Immunol. 2015, 195, 1034–1043. [Google Scholar]
- Rd, A.F.; Torres, V.J. The bicomponent pore-forming leucocidins of Staphylococcus aureus. Microbiol. Mol. Biol. Rev. MMBR 2014, 78, 199–230. [Google Scholar]
- Vandenesch, F.; Naimi, T.; Enright, M.C.; Lina, G.; Nimmo, G.R.; Heffernan, H.; Liassine, N.; Bes, M.; Greenland, T.; Reverdy, M.E.; et al. Community-acquired methicillin-resistant Staphylococcus aureus carrying Panton-Valentine leukocidin genes: Worldwide emergence. Emerg. Infect. Dis. 2003, 9, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Diep, B.A.; Gill, S.R.; Chang, R.F.; Phan, T.H.; Chen, J.H.; Davidson, M.G.; Lin, F.; Lin, J.; Carleton, H.A.; Mongodin, E.F.; et al. Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. Lancet 2006, 367, 731–739. [Google Scholar] [CrossRef]
- Lina, G.; Piemont, Y.; Godail-Gamot, F.; Bes, M.; Peter, M.O.; Gauduchon, V.; Vandenesch, F.; Etienne, J. Involvement of Panton-Valentine leukocidin-producing Staphylococcus aureus in primary skin infections and pneumonia. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 1999, 29, 1128–1132. [Google Scholar] [CrossRef] [PubMed]
- Gillet, Y.; Issartel, B.; Vanhems, P.; Fournet, J.C.; Lina, G.; Bes, M.; Vandenesch, F.; Piemont, Y.; Brousse, N.; Floret, D.; et al. Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet 2002, 359, 753–759. [Google Scholar] [CrossRef]
- Laventie, B.J.; Potrich, C.; Atmanene, C.; Saleh, M.; Joubert, O.; Viero, G.; Bachmeyer, C.; Antonini, V.; Mancini, I.; Cianferani-Sanglier, S.; et al. p-Sulfonato-calix[n]arenes inhibit staphylococcal bicomponent leukotoxins by supramolecular interactions. Biochem. J. 2013, 450, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Laventie, B.J.; Rademaker, H.J.; Saleh, M.; de Boer, E.; Janssens, R.; Bourcier, T.; Subilia, A.; Marcellin, L.; van Haperen, R.; Lebbink, J.H.; et al. Heavy chain-only antibodies and tetravalent bispecific antibody neutralizing Staphylococcus aureus leukotoxins. Proc. Natl. Acad. Sci. USA 2011, 108, 16404–16409. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, J.A.; Speeg-Schatz, C.; Freitas, F.I.; Sahel, J.; Monteil, H.; Prevost, G. Channel-forming leucotoxins from Staphylococcus aureus cause severe inflammatory reactions in a rabbit eye model. J. Med. Microbiol. 1997, 46, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Heitz, P.; Roux, M.; Keller, D.; Bourcier, T.; Sauer, A.; Prevost, G.; Gaucher, D. Panton-Valentine Leukocidin Colocalizes with Retinal Ganglion and Amacrine Cells and Activates Glial Reactions and Microglial Apoptosis. Sci. Rep. 2018, 8, 2953. [Google Scholar] [CrossRef] [PubMed]
- Spaan, A.N.; Henry, T.; Van Rooijen, W.J.; Perret, M.; Badiou, C.; Aerts, P.C.; Kemmink, J.; De Haas, C.J.; Van Kessel, K.P.; Vandenesch, F.; et al. The Staphylococcal Toxin Panton-Valentine Leukocidin Targets Human C5a Receptors. Cell Host Microbe 2013, 13, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Tromp, A.T.; Van Gent, M.; Abrial, P.; Martin, A.; Jansen, J.P.; De Haas, C.J.C.; Van Kessel, K.P.M.; Bardoel, B.W.; Kruse, E.; Bourdonnay, E.; et al. Human CD45 is an F-component-specific receptor for the staphylococcal toxin Panton-Valentine leukocidin. Nat. Microbiol. 2018, 3, 1187. [Google Scholar] [CrossRef] [PubMed]
- Winzeler, A.; Wang, J.T. Purification and culture of retinal ganglion cells from rodents. Cold Spring Harb. Protoc. 2013, 2013, 643–652. [Google Scholar]
- Sawamiphak, S.; Ritter, M.; Acker-Palmer, A. Preparation of retinal explant cultures to study ex vivo tip endothelial cell responses. Nat. Protoc. 2010, 5, 1659–1665. [Google Scholar] [CrossRef] [PubMed]
- Dyer, M.A.; Cepko, C.L. Control of Muller glial cell proliferation and activation following retinal injury. Nat. Neurosci. 2000, 3, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M.; Cardona, A.E. The myeloid cells of the central nervous system parenchyma. Nature 2010, 468, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Jover, E.; Tawk, M.Y.; Laventie, B.J.; Poulain, B.; Prevost, G. Staphylococcal leukotoxins trigger free intracellular Ca(2+) rise in neurones, signalling through acidic stores and activation of store-operated channels. Cell. Microbiol. 2013, 15, 742–758. [Google Scholar] [CrossRef] [PubMed]
- Jackson, T.L.; Antcliff, R.J.; Hillenkamp, J.; Marshall, J. Human retinal molecular weight exclusion limit and estimate of species variation. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2141–2146. [Google Scholar] [CrossRef]
- Duarte, C.B.; Ferreira, I.L.; Santos, P.F.; Carvalho, A.L.; Agostinho, P.M.; Carvalho, A.P. Glutamate in life and death of retinal amacrine cells. Gen. Pharmacol. 1998, 30, 289–295. [Google Scholar] [CrossRef]
- Eberhardt, C.; Amann, B.; Feuchtinger, A.; Hauck, S.M.; Deeg, C.A. Differential expression of inwardly rectifying K+ channels and aquaporins 4 and 5 in autoimmune uveitis indicates misbalance in Muller glial cell-dependent ion and water homeostasis. Glia 2011, 59, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Deeg, C.A.; Amann, B.; Lutz, K.; Hirmer, S.; Lutterberg, K.; Kremmer, E.; Hauck, S.M. Aquaporin 11, a regulator of water efflux at retinal Muller glial cell surface decreases concomitant with immune-mediated gliosis. J. Neuroinflammation 2016, 13, 89. [Google Scholar] [CrossRef] [PubMed]
- Reichenbach, A.; Wurm, A.; Pannicke, T.; Iandiev, I.; Wiedemann, P.; Bringmann, A. Muller cells as players in retinal degeneration and edema. Graefes Arch. Clin. Exp. Ophthalmol. 2007, 245, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Karlstetter, M.; Scholz, R.; Rutar, M.; Wong, W.T.; Provis, J.M.; Langmann, T. Retinal microglia: Just bystander or target for therapy? Prog. Retin. Eye Res. 2015, 45, 30–57. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, U.K.; Kettenmann, H. Microglia: Active sensor and versatile effector cells in the normal and pathologic brain. Nat. Neurosci. 2007, 10, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Broderick, C.; Hoek, R.M.; Forrester, J.V.; Liversidge, J.; Sedgwick, J.D.; Dick, A.D. Constitutive retinal CD200 expression regulates resident microglia and activation state of inflammatory cells during experimental autoimmune uveoretinitis. Am. J. Pathol. 2002, 161, 1669–1677. [Google Scholar] [CrossRef]
- Zhang, Y.K.; Zhao, L.; Wang, X.; Ma, W.X.; Lazere, A.; Qian, H.H.; Zhang, J.; Abu-Asab, M.; Fariss, R.N.; Roger, J.E.; et al. Repopulating retinal microglia restore endogenous organization and function under CX3CL1-CX3CR1 regulation. Sci. Adv. 2018, 4, eaap8492. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, T.J.; Niederkorn, J.Y. A novel role for TGF-beta and IL-10 in the induction of immune privilege. J. Immunol. 1998, 160, 2089–2098. [Google Scholar]
- Andjelic, S.; Lumi, X.; Vereb, Z.; Josifovska, N.; Facsko, A.; Hawlina, M.; Petrovski, G. A simple method for establishing adherent ex vivo explant cultures from human eye pathologies for use in subsequent calcium imaging and inflammatory studies. J. Immunol. Res. 2014, 2014, 232659. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Colin, D.A.; Coraiola, M.; Menestrina, G.; Monteil, H.; Prévost, G. Retrieving biological activity from LukF-PV mutants combined with different S components implies compatibility between the stem domains of these staphylococcal bicomponent leucotoxins. Infect. Immun. 2002, 70, 1310–1318. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PVL Concentration | 4 h | 8 h | 24 h |
|---|---|---|---|
| 0 (Control) | 0 | 0 | 0.81 |
| 0.176 μM | 0 | 0.22 | 2.50 |
| 0.352 μM | 0 | 1.36 | 3.67 |
| 1.76 μM | 2.25 | 2.00 | 5.00 |
| 12.48 μM | 2.00 | 6.00 | 11.67 |
| Results | Retina in Vivo | Retinal Explant Ex Vivo |
|---|---|---|
| PVL Targeted Cells | RGCs, Displaced amacrine cells | RGCs, Some cells in the interne nuclear layer, Horizontal cells |
| Cell Apoptosis | Microglial cells | Microglial cells, A subpopulation of amacrine cells |
| Effects of Glial Cells | Activation of Müller cells; Activation and apoptosis of microglial cells | Activation of Müller cells; Activation and apoptosis of microglial cells |
| Inflammatory Factors | IL-6 increased, IL-8 not increased | IL-6 increased, but not as much as in the control group; IL-8 increased, but not as much as in the control group |
| Target | Antiserum | Source | Concentration |
|---|---|---|---|
| PVL | Rabbit anti-LukS-PV polyclonal | EA-7290, Strasbourg, France | 2 µg/mL |
| C5aR | Rabbit anti-C5aR polyclonal | Abcam, Cambridge, UK | 2 µg/mL |
| Ganglion Cells | Guinea pig anti-RBPMS polyclonal | UCLA Neurobiology, Los Angeles, CA, USA | 2 µg/mL |
| Starburst Amacrine Cells | Goat anti-ChAT polyclonal | Chemicon Merck-Millipore, Temecula, CA, USA | 20 µg/mL |
| Müller Cells | Mouse anti-GFAP polyclonal | Bio-Rad AbD Serotec, Oxfordshire, UK | 2 µg/mL |
| Microglial Cells | FITC-tagged GSAI-B4 | Sigma Aldrich, Saint Louis, MO, USA | 2 µg/mL |
| Amacrine Cells | Rabbit anti-Pax 6 polyclonal | Abcam | 2 µg/mL |
| AII Amacrine Cells | Mouse anti-calretinin monoclonal | Santa Cruz Biotechnology, Heidelberg, Germany | 2 µg/mL |
| Horizontal Cells | Mouse anti-calbindin monoclonal | Santa Cruz Biotechnology | 2 µg/mL |
| Secondary Antibodies | |||
| Anti-Rabbit | Goat and donkey polyclonal Alexa 555nm-conjugated | Life Technologies, Carlsbad, CA, USA | 2 µg/mL |
| Anti-Goat | Donkey polyclonal Alexa 488-conjugated | Molecular Probes, Eugene, OR, USA | 2 µg/mL |
| Anti-Mouse | Donkey polyclonal Alexa 488-conjugated | Abcam | 2 µg/mL |
| Anti-Guinea Pig | Goat polyclonal Alexa 488-conjugated | Abcam | 2 µg/mL |
| TUNEL | DNA strand breaks | Roche Life Science, Indianapolis, IN, USA | - |
| Nuclei | Hoechst 33258 | Molecular ProbesTM, Eugene, OR, USA | 0.1 µg/mL |
| Gene names | Forward Primer | Afterward Primer |
|---|---|---|
| β-actin | 5’-gcgggacatcaaggagaag-3’ | 5’-aggaaggagggctggaaga-3’ |
| IL-6 | 5’-tcaggccaagttcaggagtg-3’ | 5’-atgaagtggatcgtggtcgt-3’ |
| IL-8 | 5’-tggctgtggctctcttgg-3’ | 5’-atttgggatggaaaggtgtg-3’ |
| TNF-α | 5’-cgtagtagcaaacccgcaag-3’ | 5’-tgagtgaggagcacgtagga-3’ |
| VEGF | 5’-cgagaccttggtggacatctt-3’ | 5’-tgcattcacatttgttgtgct-3’ |
| iNOS | 5’-ccaagccctcacctacttcc-3’ | 5’-aactcctccagcacctcca-3’ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Roux, M.J.; Picaud, S.; Keller, D.; Sauer, A.; Heitz, P.; Prévost, G.; Gaucher, D. Panton-Valentine Leucocidin Proves Direct Neuronal Targeting and Its Early Neuronal and Glial Impacts a Rabbit Retinal Explant Model. Toxins 2018, 10, 455. https://doi.org/10.3390/toxins10110455
Liu X, Roux MJ, Picaud S, Keller D, Sauer A, Heitz P, Prévost G, Gaucher D. Panton-Valentine Leucocidin Proves Direct Neuronal Targeting and Its Early Neuronal and Glial Impacts a Rabbit Retinal Explant Model. Toxins. 2018; 10(11):455. https://doi.org/10.3390/toxins10110455
Chicago/Turabian StyleLiu, XuanLi, Michel J Roux, Serge Picaud, Daniel Keller, Arnaud Sauer, Pauline Heitz, Gilles Prévost, and David Gaucher. 2018. "Panton-Valentine Leucocidin Proves Direct Neuronal Targeting and Its Early Neuronal and Glial Impacts a Rabbit Retinal Explant Model" Toxins 10, no. 11: 455. https://doi.org/10.3390/toxins10110455
APA StyleLiu, X., Roux, M. J., Picaud, S., Keller, D., Sauer, A., Heitz, P., Prévost, G., & Gaucher, D. (2018). Panton-Valentine Leucocidin Proves Direct Neuronal Targeting and Its Early Neuronal and Glial Impacts a Rabbit Retinal Explant Model. Toxins, 10(11), 455. https://doi.org/10.3390/toxins10110455