Using the Marine Rotifer Brachionus plicatilis as an Endpoint to Evaluate Whether ROS-Dependent Hemolytic Toxicity Is Involved in the Allelopathy Induced by Karenia mikimotoi


Abstract
:1. Introduction
2. Results
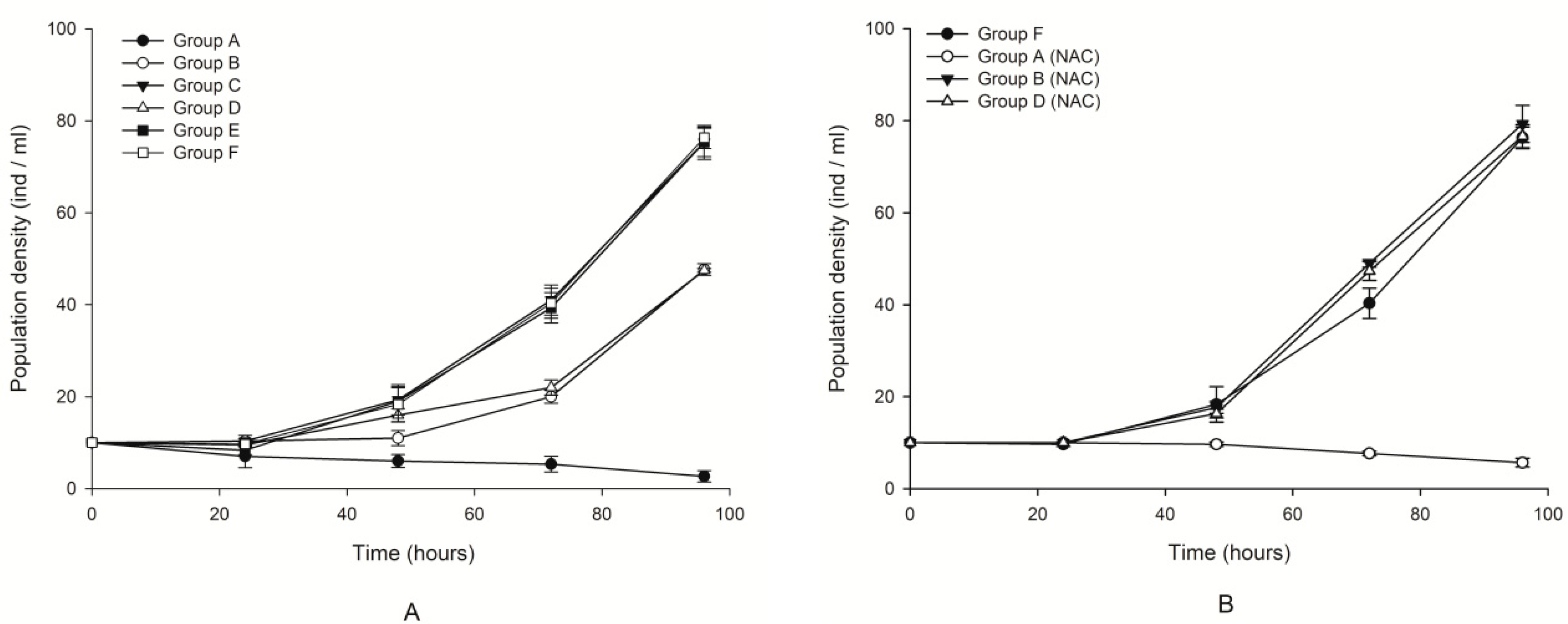
2.1. Allelopathic Effects of K. mikimotoi on B. plicatilis with and without NAC Addition
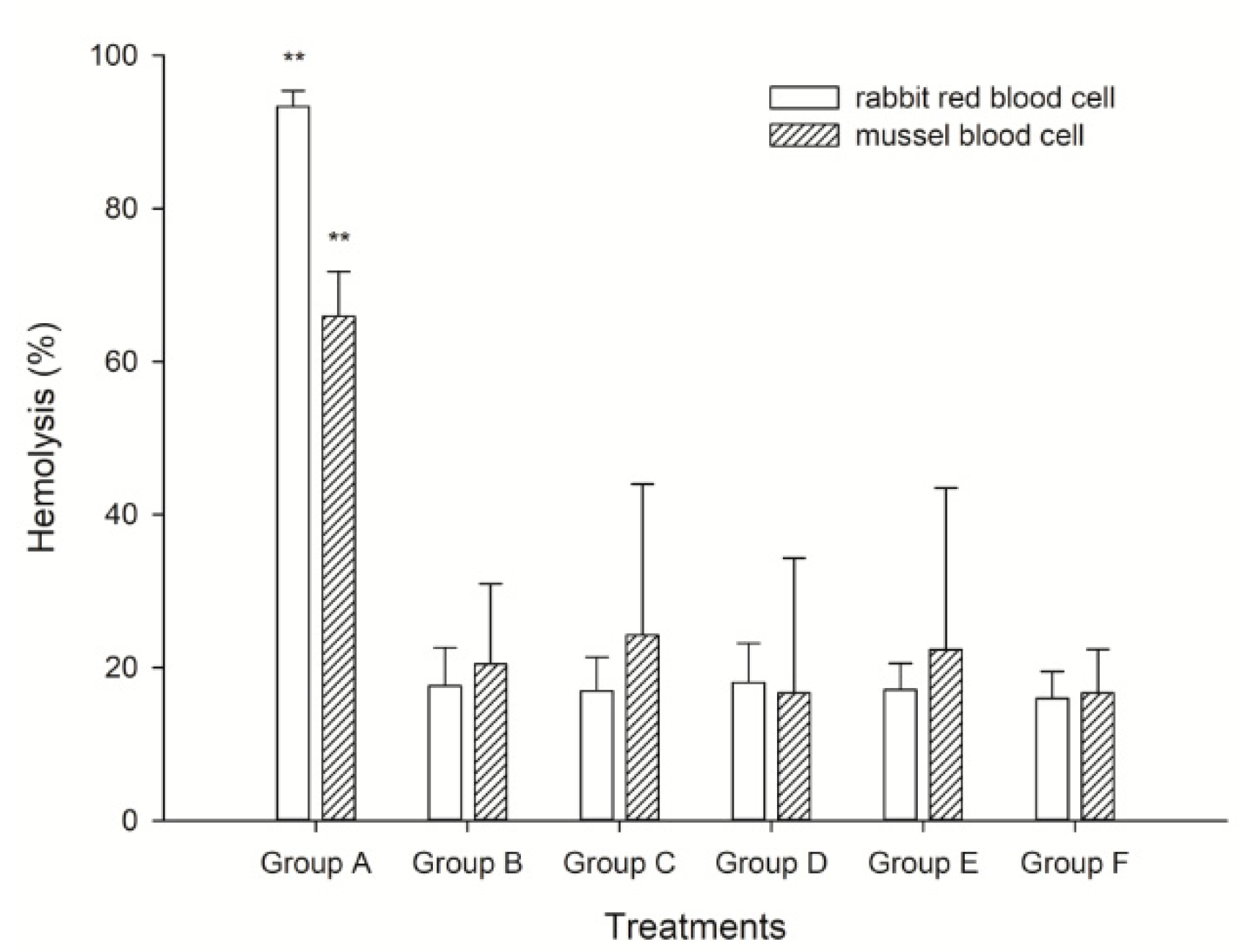
2.2. Hemolytic Activity Analysis on Mussel and Rabbit Blood Cells
3. Discussion
3.1. Is ROS Involved in the Allelopathy Induced by K. mikimotoi on the Rotifer B. plicatilis?
3.2. The Possibility of Applying the Reproductive Changes in B. plicatilis as Biomarkers to Evaluate the Toxicity of HABs
4. Conclusions
5. Materials and Methods
Organism Cultivation
6. Experimental Design
6.1. Allelopathic Effects of K. mikimotoi on the Growth of B. plicatilis
6.2. Analysis of the Hemolytic Activity of K. mikimotoi
6.3. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hallegraeff, G.M.; Bolch, C.J.; Bryan, J.; Koerbin, B. Microalgal spores in ship’s ballast water: A danger to aquaculture. In Toxic Marine Phytoplankton; Graneii, E., Sundstrom, B., Edler, L., Anderson, D.M., Eds.; Elsevier Science: Amsterdam, The Netherlands, 1990; pp. 475–480. [Google Scholar]
- Steidinger, K.A.; Tangen, K. Dinoflagellates. In Identifying Marine Diatoms & Dinoflagellates; Academic: San Diego, CA, USA, 1996; Chapter 3; pp. 387–584. [Google Scholar]
- Chang, F.H. Toxic effects of three closely-related dinoflagellates, Karenia concordia, K. brevisulcata and Karenia mikimotoi (Gymnodiniales, Dinophyceae) on other microalgal species. Harmful Algae 2011, 10, 181–187. [Google Scholar] [CrossRef]
- Dang, L.X.; Li, Y.; Liu, F.; Zhang, Y.; Yang, W.D.; Li, H.Y.; Liu, J.S. Chemical response of the toxic dinoflagellate Karenia mikimotoi against grazing by three species of Zooplankton. J. Eukaryot. Microbiol. 2015, 62, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yan, T.; Ling, J.; Yu, R.; Zhou, M. Detrimental impacts of the dinoflagellate Karenia mikimotoi, in Fujian coastal waters on typical marine organisms. Harmful Algae 2017, 61, 1–12. [Google Scholar] [CrossRef]
- Huo, Y.; Zhang, J.; Xu, S.; Tian, Q.; Zhang, Y.; He, P. Retracted: Effects of seaweed gracilaria verrucosa, on the growth of microalgae: A case study in the laboratory and in an enclosed sea of Hangzhou bay, China. Harmful Algae 2011, 10, 411–418. [Google Scholar] [CrossRef]
- Jenkinson, I.R.; Arzul, G. Mitigation by cysteine compounds of rheotoxicity, cytotoxicity and fish mortality caused by the dinoflagellates, Gymnodinium mikimotoi and G. cf. maguelonnense. In Harmful Algal Blooms 2000; Hallegraeff, G.M., Blackburn, S.I., Bolch, C.J., Lewis, R.J., Eds.; IOC: Paris, France, 2001; pp. 71–84. [Google Scholar]
- Neely, T.; Campbell, L. A modified assay to determine hemolytic toxin variability among Karenia clones isolated from the Gulf of Mexico. Harmful Algae 2006, 5, 592–598. [Google Scholar] [CrossRef]
- Mooney, B.D.; Nichols, P.D.; De Salas, M.F.; Hallegraeff, G.M. Lipid, fatty acid, and sterol composition of eight species of Kareniaceae (dinophyta): Chemotaxonomy and putative lipid phycotoxins. J. Phycol. 2007, 43, 101–111. [Google Scholar] [CrossRef]
- Satake, M.; Shoji, M.; Oshima, Y.; Naoki, H.; Fujita, T.; Yasumoto, T. Gymnocin-A, a cytotoxic polyether from the notorious red tide dinoflagellate, Gymnodinium mikimotoi. Tetrahedron Lett. 2002, 43, 5829–5832. [Google Scholar] [CrossRef]
- Satake, M.; Tanaka, Y.; Ishikura, Y.; Oshima, Y.; Naoki, H.; Yasumoto, T. Gymnocin-B with the largest contiguous polyether rings from the red tide dinoflagellate. Karenia (formerly Gymnodinium) mikimotoi. Tetrahedron Lett. 2005, 46, 3537–3540. [Google Scholar] [CrossRef]
- Yamasaki, Y.; Kim, D.I.; Matsuyama, Y.; Oda, T.; Honjo, T. Production of superoxide anion and hydrogen peroxide by the red tide dinoflagellate Karenia mikimotoi. J. Biosci. Bioeng. 2004, 97, 212–215. [Google Scholar] [CrossRef]
- Gentien, P.; Lunven, M.; Lazure, P.; Youenou, A.; Crassous, M.P. Motility and autotoxicity in Karenia mikimotoi (Dinophyceae). Philos. Trans. R. Soc. B 2007, 362, 1937–1946. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.C.; Xiao, T.Y.; Zhang, T.; Yang, C.H. A summary of research on planktonic algae allelopathy. Chin. J. Microecol. 2009, 21, 768–769. [Google Scholar]
- Inderjit; Duke, S.O. Ecophysiological aspects of allelopathy. Planta 2003, 217, 529–539. [Google Scholar] [PubMed]
- Mulderij, G.; Mooij, W.M.; Smolders, A.J.P.; Van Donk, E. Allelopathic inhibition of phytoplankton by exudates from stratiotes aloides. Aquat. Bot. 2005, 82, 284–296. [Google Scholar] [CrossRef]
- Donk, E.V.; Bund, W.J.V.D. Impact of submerged macrophytes including charophytes on phyto- and zooplankton communities: Allelopathy versus other mechanisms. Aquat. Bot. 2002, 72, 261–274. [Google Scholar] [CrossRef]
- Gentien, P.; Arzul, G. Exotoxin production by Gyrodinium cf. aureolum (Dinophyceae). J. Mar. Biol. Assoc. U. K. 1990, 70, 571–581. [Google Scholar] [CrossRef]
- Colin, S.P.; Dam, H.G. Latitudinal differentiation in the effects of the toxic dinoflagellate Alexandrium spp. on the feeding and reproduction of populations of the copepod Acartia hudsonica. Harmful Algae 2002, 1, 113–125. [Google Scholar] [CrossRef]
- Turner, J.T.; Tester, P.A. Toxic marine phytoplankton, zooplankton grazers, and pelagic food webs. Limnol. Oceanogr. 1997, 42, 1203–1204. [Google Scholar] [CrossRef]
- Liu, S.; Wang, W.X. Feeding and reproductive responses of marine copepods in South China Sea to toxic and nontoxic phytoplankton. Mar. Biol. 2002, 140, 595–603. [Google Scholar]
- Wang, L.; Yan, T.; Yu, R.; Zhou, M. Experimental study on the impact of dinoflagellate Alexandrium species on populations of the rotifer Brachionus plicatilis. Harmful Algae 2005, 4, 371–382. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, W.; Yu, G.; Huang, B.; Gao, P.; Wen, Z. The inhibition effects of microcystis aeruginosa on growth and reproduction of zooplankton. J. Xinyang Teach. Coll. 2004, 17, 437–439. [Google Scholar]
- Estrada, M.; Sala, M.M.; van Lenning, K.; Alcaraz, M.; Felipe, J.; Veldhuis, M.J.W. Biological interactions in enclosed plankton communities including Alexandrium catenella and copepods: Role of phosphorus. J. Exp. Mar. Biol. Ecol. 2008, 355, 1–11. [Google Scholar] [CrossRef]
- Kim, D.; Sato, Y.; Oda, T.; Muramatsu, T.; Matsuyama, Y.; Honjo, T. Specific toxic effect of dinoflagellate Heterocapsa circularisquama on the rotifer Brachionus plicatilis. Biosci. Biotechnol. Biochem. 2000, 64, 2719–2722. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.J. The red tide dinoflagellate Alexandrium tamarense: Effects on behavior and growth of a tintinnid ciliate. Mar. Ecol. Prog. Ser. 1989, 53, 105–116. [Google Scholar] [CrossRef]
- Hansen, P.J. Growth and grazing response of a ciliate feeding on the red tide dinoflagellate Gyrodinium aureolum in monoculture and in mixture with a nontoxic alga. Mar. Ecol. Prog. Ser. 1995, 121, 65–72. [Google Scholar] [CrossRef]
- Yang, C.Z.; Albright, L.J.; Yousif, A.N. Oxygen-radical-mediated effects of the toxic phytoplankter Heterosigma carterae on juvenile rainbow trout Oncorhynchus mykiss. Dis. Aquat. Org. 1995, 23, 101–108. [Google Scholar] [CrossRef]
- Marshall, J.A.; Ross, T.; Pyecroft, S.; Hallegraeff, G. Superoxide production by marine microalgae. Mar. Biol. 2005, 147, 533–540. [Google Scholar] [CrossRef]
- Oda, T.; Ishimatsu, A.; Shimada, M.; Takeshita, S.; Muramatsu, T. Oxygen-radical-mediated toxic effects of the red tide flagellate Chattonella marina on Vibrio alginolyticus. Mar. Biol. 1992, 112, 505–509. [Google Scholar] [CrossRef]
- Oda, T.; Nakamura, A.; Okamoto, T.; Ishimatsu, A.; Muramatsu, T. Lectin-induced enhancement of superoxide anion production by red tide phytoplankton. Mar. Biol. 1998, 131, 383–390. [Google Scholar] [CrossRef]
- Kim, C.S.; Lee, S.G.; Lee, C.K.; Kim, H.G.; Jung, J. Reactive oxygen species as causative agents in the ichthyotoxicity of the red tide dinoflagellate Cochlodinium polykrikoides. J. Plankton Res. 1999, 21, 2105–2115. [Google Scholar] [CrossRef]
- Gentien, P. Bloom dynamics and ecophysiology of the Gymnodinium mikimotoi species complex. In Physiological Ecology of Harmful Algal Blooms; Anderson, D.M., Cembella, A.D., Hallegraeff, G.M., Eds.; Springer: Berlin, Germany, 1998; pp. 155–173. [Google Scholar]
- Mitchell, S.; Rodger, H. Pathology of wild and cultured fish affected by a Karenia mikimotoi bloom in Ireland, 2005. Bull.-Eur. Assoc. Fish Pathol. 2007, 27, 39. [Google Scholar]
- Zhang, J.; Wang, Y.; Tang, X.X.; Fang, K. The toxic effects of polybrominated diphenyl ethers on rotifer Branchionus plicatilis. Adv. Mater. Res. 2013, 807–809, 385–391. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Y.; Sun, K.M.; Tang, X.X. A study of oxidative stress induced by two polybrominated diphenyl ethers in the rotifer Brachionus plicatilis. Mar. Pollut. Bull. 2016, 113, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Tang, X.; Sha, J.; Chen, H.; Sun, T.; Wang, Y. The reproductive toxicity on the rotifer Brachionus plicatilis induced by BDE-47 and studies on the effective mechanism based on antioxidant defense system changes. Chemosphere 2015, 135, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Sha, J.; Wang, Y.; Chen, H.; Wang, M.; Wang, H.; Li, X.; Qi, L.; Tang, X. Using population demographic parameters to assess impacts of two polybrominated diphenyl ethers (BDE-47, BDE-209) on the rotifer Brachionus plicatilis. Ecotoxicol. Environ. Saf. 2015, 119, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, Y.; Uchida, T.; Honjo, T. Toxic effects of the dinoflagellate Heterocapsa circularisquama on clearance rate of the blue mussel Mytilus galloprovincialis. Mar. Ecol. Prog. Ser. 1997, 146, 73–80. [Google Scholar] [CrossRef]
- Cho, Y.; Ozeki, R.; Yotsu-Yamashita, M.; Oshima, Y. Single-cell analysis of paralytic shellfish toxins in Alexandrium tamarense, by HPLC with post-column fluorescent derivatization. Harmful Algae 2013, 25, 47–53. [Google Scholar] [CrossRef]
- Kim, C.H.; Sako, Y.; Ishida, Y. Variation of Toxin Production and Composition in Axenic Cultures of Alexandrium catenella and A. tamarense. Nippon Suisan Gakkaishi 1993, 59, 633–639. [Google Scholar] [CrossRef]
- Kim, D.; Nakamura, A.; Okamoto, T.; Komatsu, N.; Oda, T.; Ishimatsu, A. Toxic potential of the raphidophyte Olisthodiscus luteus: Mediation by reactive oxygen species. J. Plankton Res. 1999, 21, 1017–1027. [Google Scholar] [CrossRef]
- Oda, T.; Nakamura, A.; Shikayama, M.; Kawano, I.; Ishimatsu, A.; Muramatsu, T. Generation of reactive oxygen species by raphidophycean phytoplankton. Biosci. Biotechnol. Biochem. 1997, 61, 1658–1662. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Akaike, T.; Sato, K.; Ishimatsu, A.; Takeshita, S.; Muramatsu, T.; Maeda, H. Hydroxyl radical generation by red tide algae. Arch. Biochem. Biophys. 1992, 294, 38–43. [Google Scholar] [CrossRef]
- Ishimatsu, A.; Oda, T.; Yoshida, M.; Ozaki, M. Oxygen radicals are probably involved in the mortality of yellowtail by Chattonella marina. J. Fish. Sci. 1996, 62, 836–837. [Google Scholar] [CrossRef]
- Kim, D.; Nakashima, T.; Matsuyama, Y.; Niwano, Y.; Yamaguchi, K.; Oda, T. Presence of the distinct systems responsible for superoxide anion and hydrogen peroxide generation in red tide phytoplankton Chattonella marina and Chattonella ovata. J. Plankton Res. 2007, 29, 241–247. [Google Scholar] [CrossRef]
- Kim, D.; Yamasaki, Y.; Yamatogi, T.; Yamaguchi, K.; Matsuyama, Y.; Kang, Y.S.; Lee, Y.; Oda, T. The possibility of reactive oxygen species (ROS)-indipendent toxic effects of Cochlodinium polykrikoides on damselfish (Chromis caerulea). Biosci. Biotechnol. Biochem. 2009, 73, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Sato, Y.; Kim, D.; Muramatsu, T.; Matsuyama, Y.; Honjo, T. Hemolytic activity of Heterocapsa circularisquama (Dinophyceae) and its possible involvement in shellfish toxicity. J. Phycol. 2001, 37, 509–516. [Google Scholar] [CrossRef]
- Sato, Y.; Oda, T.; Muramatsu, T.; Matsuyama, Y.; Honjo, T. Photosensitizing hemolytic toxin in Heterocapsa circularisquama, a newly identified harmful red tide dinoflagellate. Aquat. Toxicol. 2002, 56, 191–196. [Google Scholar] [CrossRef]
- Zou, Y.; Yamasaki, Y.; Matsuyama, Y.; Yamaguchi, K.; Honjo, T.; Oda, T. Possible involvement of hemolytic activity in the contact-dependent lethal effects of the dinoflagellate Karenia mikimotoi on the rotifer Brachionus plicatilis. Harmful Algae 2010, 9, 367–373. [Google Scholar] [CrossRef]
- Williams, P.L.; Dusenbery, D.B. Using the nematode Caenorhabditis elegans to predict mammalian acute lethality to metallic salts. Toxicol. Ind. Health 1988, 4, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.L.; Dusenbery, D.B. A promising indicator of neurobehavioral toxicity using the nematode Caenorhabditis elegans and computer tracking. Toxicol. Ind. Health 1990, 6, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Middendorf, P.J.; Dusenbery, D.B. Fluoroacetic Acid Is a Potent and Specific Inhibitor of Reproduction in the Nematode Caenorhabditis elegans. J. Nematol. 1993, 25, 573–577. [Google Scholar] [PubMed]
- Dhawan, R.; Dusenbery, D.B.; Williams, P.L. Comparison of lethality, reproduction, and behavior as toxicological endpoints in the nematode Caenorhabditis elegans. Toxicol. Environ. Health Part A 1999, 58, 451–462. [Google Scholar]
- Rao, T.R.; Sarma, S.S.S. Demographic parameters of Brachionus patulus, Muller (Rotifera) exposed to sublethal DDT concentrations at low and high food levels. J. Hydrobiol. 1986, 139, 193–200. [Google Scholar]
- Kim, D.; Nakamura, A.; Okamoto, T.; Komatsu, N.; Oda, T.; Iida, T.; Ishimatsu, A.; Muramatsu, T. Mechanism of superoxide anion generation in the toxic red tide phytoplankton Chattonella marina: Possible involvement of NAD(P)H oxidase. Biochim. Biophys. Acta 2000, 1524, 228–232. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Animals; Smith, W.L., Chanley, M.H., Eds.; Plenum Press: New York, NY, USA, 1975; pp. 26–60. [Google Scholar]
- Zhang, X.; Tang, X.; Zhou, B.; Wang, Y. Effects of decabromodiphenyl ether (BDE-209) on inter-specific competition between two species of marine bloom-forming microalgae. PLoS ONE 2013, 8, e56084. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Tang, X.; Zhou, B.; Jiang, Y.; Lv, M.; Zang, Y.; Wang, Y. Is it photosensitization or photodegradation when UV-B irradiation is combined with BDE-47? Evidence from the growth and reproduction changes of rotifer Brachionus plicatilis. Sci. Total Environ. 2018, 628–629, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Radix, P.; Severin, G.; Schramm, K.-W.; Kettrup, A. Reproduction disturbances of Brachionus calyciflorus (rotifer) for the screening of environmental endocrine disrupters. Chemosphere 2002, 47, 1097–1101. [Google Scholar] [CrossRef]
- Hitchcock, D.R.; Mccutcheon, S.C.; Smith, M.C. Using rotifer population demographic parameters to assess impacts of the degradation products from trinitrotoluene phytoremediation. Ecotoxicol. Environ. Saf. 2003, 55, 143–151. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecology: The experimental analysis of distribution and abundance. Q. Rev. Biol. 1985, 48, 133–148. [Google Scholar]
- Pianka, E.R. Evolutionary Ecology; Harper and Row: New York, NY, USA, 1978. [Google Scholar]
- Jiang, Y.; Tang, X.; Sun, T.; Wang, Y. BDE-47 exposure changed the immune function of haemocytes in Mytilus edulis: An explanation based on ROS-mediated pathway. Aquat. Toxicol. 2017, 182, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Caffrey, J.M.; Dasmahapatra, A.; Smith, H.A.; Hede, K.; Frieden, E. The effect of copper ion on glutathione and hemolysis in rabbit erythrocytes. J. Bio. Trace Elem. Res. 1986, 11, 19–26. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Fluids | The Population Growth Rate r |
|---|---|
| group A | −0.3685 ± 0.1493 d |
| group B | 0.3903 ± 0.0066 b |
| group C | 0.5045 ± 0.0122 a |
| group D | 0.3895 ± 0.0021 b |
| group E | 0.5046 ± 0.0104 a |
| group F | 0.5080 ± 0.0078 a |
| group A (+NAC) | −0.1452 ± 0.0397 c |
| group B (+NAC) | 0.5175 ± 0.0125 a |
| group D (+NAC) | 0.5091 ± 0.0081 a |
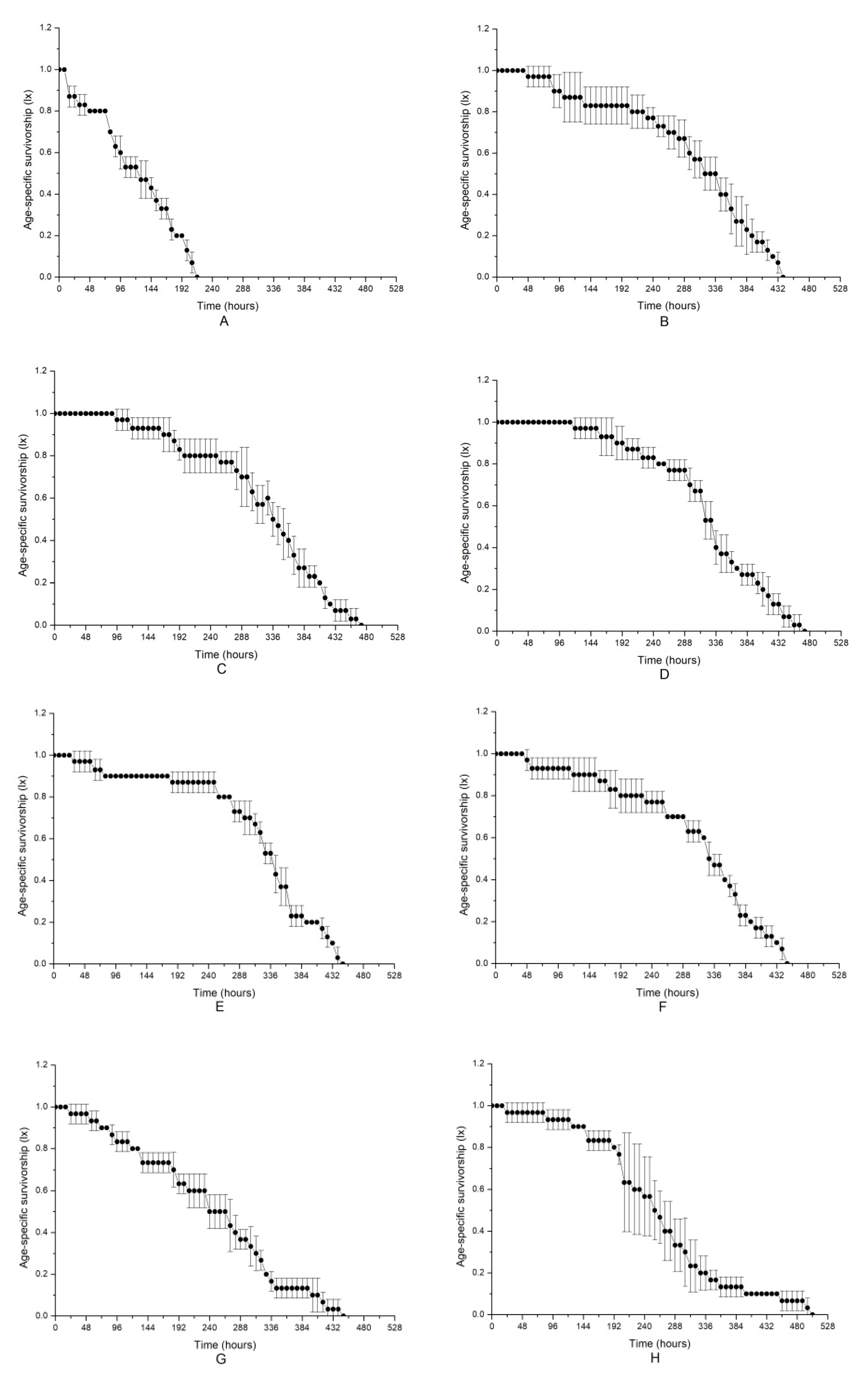
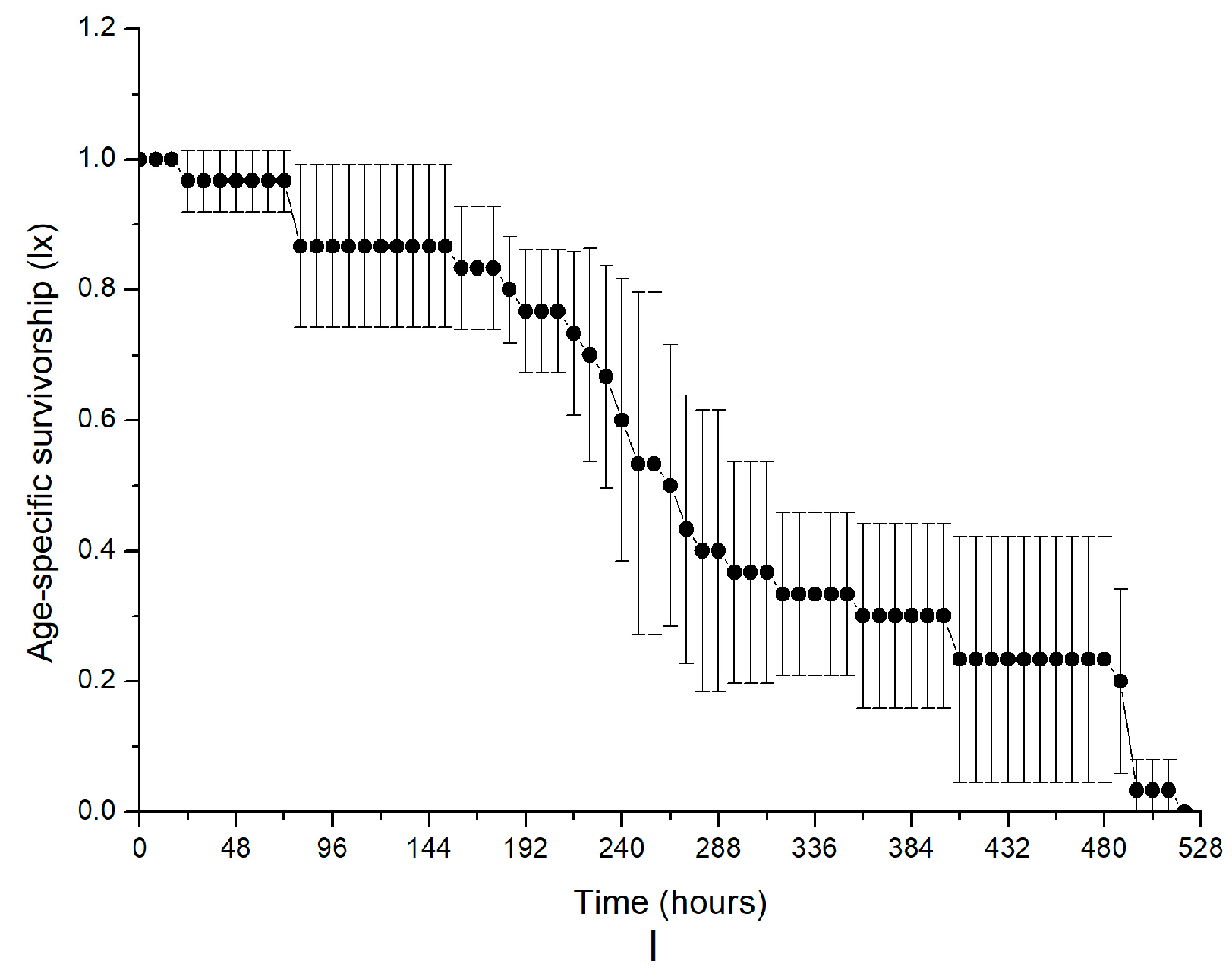
| Parameter | R0 | T | rm | E0 | λ |
|---|---|---|---|---|---|
| Group A | 2.0667 ± 0.3399 b | 5.1130 ± 0.1568 d | 0.1386 ± 0.0322 e | 4.5000 ± 0.2449 c | 1.1492 ± 0.0367 e |
| Group B | 12.6667 ± 2.2005 a | 7.0630 ± 0.2211 ab | 0.3571 ± 0.0167 c | 12.0000 ± 1.2570 ab | 1.4294 ± 0.0237 c |
| Group C | 13.5667 ± 1.6977 a | 7.2136 ± 0.2958 a | 0.3602 ± 0.0036 c | 12.6333 ± 0.7409 a | 1.4336 ± 0.0052 c |
| Group D | 15.1000 ± 2.6470 a | 6.8249 ± 0.3748 ab | 0.3953 ± 0.0144 bc | 13.0333 ± 0.2357 a | 1.4850 ± 0.0214 c |
| Group E | 13.6333 ± 2.9318 a | 6.5161 ± 0.6245 b | 0.4010 ± 0.0456 bc | 12.7333 ± 0.0471 a | 1.4948 ± 0.0672 bc |
| Group F | 15.6333 ± 2.4226 a | 6.4576 ± 0.2978 bc | 0.4239 ± 0.0088 b | 12.2333 ± 0.4497 ab | 1.5280 ± 0.0135 bc |
| Group A(NAC) | 3.3667 ± 0.3771 b | 5.2338 ± 0.3310 cd | 0.2315 ± 0.0231 d | 9.3000 ± 0.2944 b | 1.2609 ± 0.0290 d |
| Group B(NAC) | 13.7000 ± 0.5354 a | 5.3092 ± 0.0751 cd | 0.4929 ± 0.0106 a | 10.4000 ± 0.9899 b | 1.6372 ± 0.0174 a |
| Group D(NAC) | 12.9667 ± 0.9463 a | 5.8012 ± 0.1915 c | 0.4413 ± 0.0038 b | 11.5667 ± 2.4074 ab | 1.5548 ± 0.0059 b |
| Groups | The First Spawning Time | The Last Spawning Time | The Average Number of Eggs | The Average Number of Larvae | Hatchability |
|---|---|---|---|---|---|
| Group A | 84.000 ± 0.001 a | 174.667 ± 11.624 c | 20.667 ± 2.404 b | 26.667 ± 3.712 b | 79.085 ± 10.519 b |
| Group B | 70.667 ± 1.333 b | 308.000 ± 16.000 a | 126.667 ± 15.560 a | 133.000 ± 15.822 a | 95.153 ± 1.850 a |
| Group C | 60.000 ± 4.619 c | 300.000 ± 16.653 ab | 135.667 ± 12.005 a | 142.667 ± 14.518 a | 95.384 ± 1.434 a |
| Group D | 56.000 ± 4.000 cd | 310.667 ± 17.487 a | 151.000 ± 18.717 a | 153.000 ± 19.088 a | 98.724 ± 0.308 a |
| Group E | 53.333 ± 1.333 cd | 284.000 ± 36.661 ab | 136.333 ± 20.731 a | 147.667 ± 17.629 a | 91.834 ± 4.234 a |
| Group F | 52.000 ± 2.309 d | 305.333 ± 26.667 a | 156.333 ± 17.130 a | 161.333 ± 15.836 a | 96.725 ± 2.170 a |
| Group A (NAC) | 60.000 ± 4.000 c | 316.000 ± 20.000 a | 33.667 ± 2.667 b | 38.000 ± 1.155 b | 88.397 ± 4.736 ab |
| Group B (NAC) | 54.667 ± 1.333 cd | 226.667 ± 7.055 bc | 148.667 ± 5.044 a | 156.333 ± 6.119 a | 95.187 ± 2.268 a |
| Group D (NAC) | 52.000 ± 2.309 d | 240.000 ± 16.000 b | 129.667 ± 6.692 a | 139.000 ± 2.082 a | 93.187 ± 3.407 a |
| Group | Definition | Treatment Conditions | Growth Phase |
|---|---|---|---|
| Treatment group | Group A | Whole cell suspension | Plateau phase |
| Group B | Lysed cell suspension | Exponential phase | |
| Group C | Cell-free culture filtrates | Exponential phase | |
| Group D | Lysed cell suspension | Plateau phase | |
| Group E | Cell-free culture filtrates | Plateau phase | |
| Control group | Group F | Sterilized seawater |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Yu, J.; Sun, T.; Liu, C.; Sun, Y.; Wang, Y. Using the Marine Rotifer Brachionus plicatilis as an Endpoint to Evaluate Whether ROS-Dependent Hemolytic Toxicity Is Involved in the Allelopathy Induced by Karenia mikimotoi. Toxins 2018, 10, 439. https://doi.org/10.3390/toxins10110439
Li Y, Yu J, Sun T, Liu C, Sun Y, Wang Y. Using the Marine Rotifer Brachionus plicatilis as an Endpoint to Evaluate Whether ROS-Dependent Hemolytic Toxicity Is Involved in the Allelopathy Induced by Karenia mikimotoi. Toxins. 2018; 10(11):439. https://doi.org/10.3390/toxins10110439
Chicago/Turabian StyleLi, Yuanyuan, Jianfei Yu, Tianli Sun, Chunchen Liu, Yu Sun, and You Wang. 2018. "Using the Marine Rotifer Brachionus plicatilis as an Endpoint to Evaluate Whether ROS-Dependent Hemolytic Toxicity Is Involved in the Allelopathy Induced by Karenia mikimotoi" Toxins 10, no. 11: 439. https://doi.org/10.3390/toxins10110439
APA StyleLi, Y., Yu, J., Sun, T., Liu, C., Sun, Y., & Wang, Y. (2018). Using the Marine Rotifer Brachionus plicatilis as an Endpoint to Evaluate Whether ROS-Dependent Hemolytic Toxicity Is Involved in the Allelopathy Induced by Karenia mikimotoi. Toxins, 10(11), 439. https://doi.org/10.3390/toxins10110439