Human Milk and Allergic Diseases: An Unsolved Puzzle

 , , , , , ,
, , , , , , 
Abstract
1. Introduction
2. Breastfeeding and Immunological Outcomes
2.1. Importance of Breastfeeding Duration
2.2. Breastfeeding and Allergic Diseases
2.2.1. Eczema
2.2.2. Food Allergy
2.2.3. Asthma
2.3. Breastfeeding, Thymus and Immunity
3. Human Milk Composition and Allergy
3.1. Human Milk Immunological Composition
3.1.1. Immune Composition and Allergy
3.1.2. Potential for Immunological Composition Alteration via Dietary Interventions
3.2. Human Milk Oligosaccharides
3.2.1. The Fascinating Complexity of Human Milk Oligosaccharides
3.2.2. Shaping the Microbial Balance in Early Life
3.2.3. HMOs Are Directly Involved in Early Life Immune Development
3.3. Human Milk Microbiota
3.4. Human Milk Micronutrients
3.4.1. Vitamin A
3.4.2. B Vitamins
3.4.3. Vitamin D
3.4.4. Iron
3.4.5. Zinc
3.4.6. Summary
3.5. The New Frontier: Human Milk Glycoproteins and Metabolites
3.5.1. Secretory Immunoglobulin A (sIgA)
3.5.2. Lactoferrrin
3.5.3. Low Molecular Weight Metabolites
Milk Fatty Acids
Choline
Lactose
4. Breastfeeding/Human Milk Research Unmet Needs
- ➢
- Large and well-standardised studies of HM composition (integrated data on immune markers, HMOs, PUFAs, microbiome and metabolites), defining lactotypes and assessing variation between women residing in different countries
- ➢
- Application of omics approaches (metabolomics, proteomics, genomics, etc.) to highlight the most important components of HM in relation to allergic diseases
- ➢
- Studies evaluating biological activity of a specific components within HM
- ➢
- Randomised trials of breastfeeding interventions with long-term follow-up for allergic disease development
- ➢
- Randomised trials of early weaning (3–4 months) using different dietary approaches
- ➢
- Large cohort studies which combine assessments of breastfeeding influence on allergy development with the constituent analysis of HM samples
- ➢
- Development of a new intervention strategies for HM composition modification and indirect preventative effect on allergy prevention
- ➢
- Relevance of a geographical location/lifestyle/diet and its’ influence on the composition of human milk should be assessed in more detail and research should account for these important confounders
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Asher, M.I.; Montefort, S.; Bjorksten, B.; Lai, C.K.; Strachan, D.P.; Weiland, S.K.; Williams, H.; ISAAC Phase Three Study Group. Worldwide time trends in the prevalence of symptoms of asthma, allergic rhinoconjunctivitis, and eczema in childhood: Isaac phases one and three repeat multicountry cross-sectional surveys. Lancet 2006, 368, 733–743. [Google Scholar] [CrossRef]
- Williams, H.; Robertson, C.; Stewart, A.; Ait-Khaled, N.; Anabwani, G.; Anderson, R.; Asher, I.; Beasley, R.; Bjorksten, B.; Burr, M.; et al. Worldwide variations in the prevalence of symptoms of atopic eczema in the international study of asthma and allergies in childhood. J. Allergy Clin. Immunol. 1999, 103, 125–138. [Google Scholar] [CrossRef]
- Kung, S.J.; Steenhoff, A.P.; Gray, C. Food allergy in africa: Myth or reality? Clin. Rev. Allergy Immunol. 2014, 46, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Strachan, D.P. Hay fever, hygiene, and household size. BMJ 1989, 299, 1259–1260. [Google Scholar] [CrossRef] [PubMed]
- Strachan, D.P. Family size, infection and atopy: The first decade of the “hygiene hypothesis”. Thorax 2000, 55, S2–S10. [Google Scholar] [CrossRef] [PubMed]
- Sozanska, B.; Blaszczyk, M.; Pearce, N.; Cullinan, P. Atopy and allergic respiratory disease in rural poland before and after accession to the european union. J. Allergy Clin. Immunol. 2014, 133, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, G.; Tsakok, T.; Lack, S.; Lack, G. Prevention of food allergy. J. Allergy Clin. Immunol. 2016, 137, 998–1010. [Google Scholar] [CrossRef] [PubMed]
- Greer, F.R.; Sicherer, S.H.; Burks, A.W.; American Academy of Pediatrics Committee on Nutrition; American Academy of Pediatrics Section on Allergy and Immunology. Effects of early nutritional interventions on the development of atopic disease in infants and children: The role of maternal dietary restriction, breastfeeding, timing of introduction of complementary foods, and hydrolyzed formulas. Pediatrics 2008, 121, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Host, A.; Halken, S.; Muraro, A.; Dreborg, S.; Niggemann, B.; Aalberse, R.; Arshad, S.H.; von Berg, A.; Carlsen, K.H.; Duschen, K.; et al. Dietary prevention of allergic diseases in infants and small children. Pediatr. Allergy Immunol. 2008, 19, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Kjellman, N.I. Prediction and prevention of atopic allergy. Allergy 1998, 53, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Muraro, A.; Halken, S.; Arshad, S.H.; Beyer, K.; Dubois, A.E.; Du Toit, G.; Eigenmann, P.A.; Grimshaw, K.E.; Hoest, A.; Lack, G.; et al. Eaaci food allergy and anaphylaxis guidelines. Primary prevention of food allergy. Allergy 2014, 69, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Prescott, S.L.; Tang, M.L.; Australasian Society of Clinical Immunology and Allergy. The australasian society of clinical immunology and allergy position statement: Summary of allergy prevention in children. Med. J. Aust. 2005, 182, 464–467. [Google Scholar] [PubMed]
- D’Alessandro, A.; Scaloni, A.; Zolla, L. Human milk proteins: An interactomics and updated functional overview. J. Proteome Res. 2010, 9, 3339–3373. [Google Scholar] [CrossRef] [PubMed]
- Holtzman, M.J. Asthma as a chronic disease of the innate and adaptive immune systems responding to viruses and allergens. J. Clin. Investig. 2012, 122, 2741–2748. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Polk, D.B. Probiotics and immune health. Curr. Opin. Gastroenterol. 2011, 27, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Munblit, D.; Verhasselt, V. Allergy prevention by breastfeeding: Possible mechanisms and evidence from human cohorts. Curr. Opin. Allergy Clin. Immunol. 2016, 16, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Lumia, M.; Luukkainen, P.; Kaila, M.; Tapanainen, H.; Takkinen, H.M.; Prasad, M.; Niinisto, S.; Nwaru, B.I.; Kenward, M.G.; Ilonen, J.; et al. Maternal dietary fat and fatty acid intake during lactation and the risk of asthma in the offspring. Acta Paediatr. 2012, 101, e337–e343. [Google Scholar] [CrossRef] [PubMed]
- Joseph, C.L.; Ownby, D.R.; Havstad, S.L.; Woodcroft, K.J.; Wegienka, G.; MacKechnie, H.; Zoratti, E.; Peterson, E.L.; Johnson, C.C. Early complementary feeding and risk of food sensitization in a birth cohort. J. Allergy Clin. Immunol. 2011, 127, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Nwaru, B.I.; Craig, L.C.; Allan, K.; Prabhu, N.; Turner, S.W.; McNeill, G.; Erkkola, M.; Seaton, A.; Devereux, G. Breastfeeding and introduction of complementary foods during infancy in relation to the risk of asthma and atopic diseases up to 10 years. Clin. Exp. Allergy 2013, 43, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Nwaru, B.I.; Takkinen, H.M.; Niemela, O.; Kaila, M.; Erkkola, M.; Ahonen, S.; Haapala, A.M.; Kenward, M.G.; Pekkanen, J.; Lahesmaa, R.; et al. Timing of infant feeding in relation to childhood asthma and allergic diseases. J. Allergy Clin. Immunol. 2013, 131, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Matheson, M.C.; Allen, K.J.; Tang, M.L. Understanding the evidence for and against the role of breastfeeding in allergy prevention. Clin. Exp. Allergy 2012, 42, 827–851. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Becker, A.B.; Guttman, D.S.; Sears, M.R.; Scott, J.A.; Kozyrskyj, A.L.; Canadian Healthy Infant Longitudinal Development Study, I. Gut microbiota diversity and atopic disease: Does breast-feeding play a role? J. Allergy Clin. Immunol. 2013, 131, 247–248. [Google Scholar] [CrossRef] [PubMed]
- Prietl, B.; Treiber, G.; Pieber, T.R.; Amrein, K. Vitamin d and immune function. Nutrients 2013, 5, 2502–2521. [Google Scholar] [CrossRef] [PubMed]
- Victora, C.G.; Horta, B.L.; Loret de Mola, C.; Quevedo, L.; Pinheiro, R.T.; Gigante, D.P.; Goncalves, H.; Barros, F.C. Association between breastfeeding and intelligence, educational attainment, and income at 30 years of age: A prospective birth cohort study from brazil. Lancet Glob. Health 2015, 3, e199–e205. [Google Scholar] [CrossRef]
- Victora, C.G.; Bahl, R.; Barros, A.J.; Franca, G.V.; Horton, S.; Krasevec, J.; Murch, S.; Sankar, M.J.; Walker, N.; Rollins, N.C.; et al. Breastfeeding in the 21st century: Epidemiology, mechanisms, and lifelong effect. Lancet 2016, 387, 475–490. [Google Scholar] [CrossRef]
- Horta, B.; Bahl, R.; Martines, J.; Victoria, C.G. Evidence of the Long-Term Effects of Breastfeeding: Systematic Reviews and Meta-Analysis; WHO: Geneva, Switzerland, 2007. [Google Scholar]
- Akobeng, A.K.; Ramanan, A.V.; Buchan, I.; Heller, R.F. Effect of breast feeding on risk of coeliac disease: A systematic review and meta-analysis of observational studies. Arch. Dis. Child. 2006, 91, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Vriezinga, S.L.; Auricchio, R.; Bravi, E.; Castillejo, G.; Chmielewska, A.; Crespo Escobar, P.; Kolacek, S.; Koletzko, S.; Korponay-Szabo, I.R.; Mummert, E.; et al. Randomized feeding intervention in infants at high risk for celiac disease. N. Engl. J. Med. 2014, 371, 1304–1315. [Google Scholar] [CrossRef] [PubMed]
- Perkin, M.R.; Logan, K.; Tseng, A.; Raji, B.; Ayis, S.; Peacock, J.; Brough, H.; Marrs, T.; Radulovic, S.; Craven, J.; et al. Randomized trial of introduction of allergenic foods in breast-fed infants. N. Engl. J. Med. 2016, 374, 1733–1743. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Strategy for Infant and Young Child Feeding, the Optimal Duration of Exclusive Breastfeeding; World Health Organization: Geneva, Switzerland, 2001. [Google Scholar]
- Agarwal, S.; Karmaus, W.; Davis, S.; Gangur, V. Immune markers in breast milk and fetal and maternal body fluids: A systematic review of perinatal concentrations. J. Hum. Lact. 2011, 27, 171–186. [Google Scholar] [CrossRef] [PubMed]
- Wells, H.G.; Osborne, T.B. The biological reactions of the vegetable proteins: Anaphylaxis. J. Infect. Dis. 1911, 8, 66–124. [Google Scholar] [CrossRef]
- Du Toit, G.; Roberts, G.; Sayre, P.H.; Bahnson, H.T.; Radulovic, S.; Santos, A.F.; Brough, H.A.; Phippard, D.; Basting, M.; Feeney, M.; et al. Randomized trial of peanut consumption in infants at risk for peanut allergy. N. Engl. J. Med. 2015, 372, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Ierodiakonou, D.; Garcia-Larsen, V.; Logan, A.; Groome, A.; Cunha, S.; Chivinge, J.; Robinson, Z.; Geoghegan, N.; Jarrold, K.; Reeves, T.; et al. Timing of allergenic food introduction to the infant diet and risk of allergic or autoimmune disease: A systematic review and meta-analysis. J. Am. Med. Assoc. 2016, 316, 1181–1192. [Google Scholar] [CrossRef] [PubMed]
- Grulee, C.; Sanford, H. The influence of breast and artificial feeding on infantile eczema. J. Pediatr. 1936, 9, 223–225. [Google Scholar] [CrossRef]
- Lowe, A.J.; Thien, F.C.; Stoney, R.M.; Bennett, C.M.; Hosking, C.S.; Hill, D.J.; Carlin, J.B.; Abramson, M.J.; Dharmage, S.C. Associations between fatty acids in colostrum and breast milk and risk of allergic disease. Clin. Exp. Allergy 2008, 38, 1745–1751. [Google Scholar] [CrossRef] [PubMed]
- Wijga, A.H.; van Houwelingen, A.C.; Kerkhof, M.; Tabak, C.; de Jongste, J.C.; Gerritsen, J.; Boshuizen, H.; Brunekreef, B.; Smit, H.A. Breast milk fatty acids and allergic disease in preschool children: The prevention and incidence of asthma and mite allergy birth cohort study. J. Allergy Clin. Immunol. 2006, 117, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Oddy, W.H.; Pal, S.; Kusel, M.M.; Vine, D.; de Klerk, N.H.; Hartmann, P.; Holt, P.G.; Sly, P.D.; Burton, P.R.; Stanley, F.J.; et al. Atopy, eczema and breast milk fatty acids in a high-risk cohort of children followed from birth to 5 yr. Pediatr. Allergy Immunol. 2006, 17, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Oken, E.; Bogdanovich, N.; Matush, L.; Sevkovskaya, Z.; Chalmers, B.; Hodnett, E.D.; Vilchuck, K.; Kramer, M.S.; Martin, R.M. Cohort profile: The promotion of breastfeeding intervention trial (probit). Int. J. Epidemiol. 2013, 43, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Flohr, C.; Nagel, G.; Weinmayr, G.; Kleiner, A.; Strachan, D.P.; Williams, H.C.; ISAAC Phase Three Study Group. Lack of evidence for a protective effect of prolonged breastfeeding on childhood eczema: Lessons from the international study of asthma and allergies in childhood (isaac) phase two. Br. J. Dermatol. 2011, 165, 1280–1289. [Google Scholar] [CrossRef] [PubMed]
- Giwercman, C.; Halkjaer, L.B.; Jensen, S.M.; Bonnelykke, K.; Lauritzen, L.; Bisgaard, H. Increased risk of eczema but reduced risk of early wheezy disorder from exclusive breast-feeding in high-risk infants. J. Allergy Clin. Immunol. 2010, 125, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.T.; Wu, C.C.; Ou, C.Y.; Chang, J.C.; Liu, C.A.; Wang, C.L.; Chuang, H.; Kuo, H.C.; Hsu, T.Y.; Chen, C.P.; et al. A prospective birth cohort study of different risk factors for development of allergic diseases in offspring of non-atopic parents. Oncotarget 2017, 8, 10858–10870. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Rha, Y.H.; Oh, I.H.; Choi, Y.S.; Kim, Y.E.; Choi, S.H. Does breast-feeding relate to development of atopic dermatitis in young korean children? Based on the fourth and fifth korea national health and nutrition examination survey 2007–2012. Allergy Asthma Immunol. Res. 2017, 9, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Robinson, D.C.; Williams, H.; Pearce, A.; Law, C.; Hope, S. Do early-life exposures explain why more advantaged children get eczema? Findings from the u.K. Millennium cohort study. Br. J. Dermatol. 2016, 174, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Draaisma, E.; Garcia-Marcos, L.; Mallol, J.; Sole, D.; Perez-Fernandez, V.; Brand, P.L.; Group, E.S. A multinational study to compare prevalence of atopic dermatitis in the first year of life. Pediatr. Allergy Immunol. 2015, 26, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, N.; Odenwald, H.; Kukkonen, A.K.; Kuitunen, M.; Savilahti, E.; Kunz, C. Fut2-dependent breast milk oligosaccharides and allergy at 2 and 5 years of age in infants with high hereditary allergy risk. Eur. J. Nutr. 2017, 56, 1293–1301. [Google Scholar] [CrossRef] [PubMed]
- Jelding-Dannemand, E.; Malby Schoos, A.M.; Bisgaard, H. Breast-feeding does not protect against allergic sensitization in early childhood and allergy-associated disease at age 7 years. J. Allergy Clin. Immunol. 2015, 136, 1302–1308. [Google Scholar] [CrossRef] [PubMed]
- Elbert, N.J.; van Meel, E.R.; den Dekker, H.T.; de Jong, N.W.; Nijsten, T.E.C.; Jaddoe, V.W.V.; de Jongste, J.C.; Pasmans, S.; Duijts, L. Duration and exclusiveness of breastfeeding and risk of childhood atopic diseases. Allergy 2017. [Google Scholar] [CrossRef] [PubMed]
- Bion, V.; Lockett, G.A.; Soto-Ramirez, N.; Zhang, H.; Venter, C.; Karmaus, W.; Holloway, J.W.; Arshad, S.H. Evaluating the efficacy of breastfeeding guidelines on long-term outcomes for allergic disease. Allergy 2016, 71, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Elliott, L.; Henderson, J.; Northstone, K.; Chiu, G.Y.; Dunson, D.; London, S.J. Prospective study of breast-feeding in relation to wheeze, atopy, and bronchial hyperresponsiveness in the avon longitudinal study of parents and children (alspac). J. Allergy Clin. Immunol. 2008, 122, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Friedman, N.J.; Zeiger, R.S. The role of breast-feeding in the development of allergies and asthma. J. Allergy Clin. Immunol. 2005, 115, 1238–1248. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Kang, M.J.; Kwon, J.W.; Park, K.S.; Hong, S.J. Breastfeeding might have protective effects on atopy in children with the cd14c-159t ct/cc genotype. Allergy Asthma Immunol. Res. 2013, 5, 239–241. [Google Scholar] [CrossRef] [PubMed]
- Sakihara, T.; Sugiura, S.; Ito, K. The ingestion of cow’s milk formula in the first 3 months of life prevents the development of cow’s milk allergy. Asia Pac. Allergy 2016, 6, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Onizawa, Y.; Noguchi, E.; Okada, M.; Sumazaki, R.; Hayashi, D. The association of the delayed introduction of cow’s milk with ige-mediated cow’s milk allergies. J. Allergy Clin. Immunol. Pract. 2016, 4, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Warstedt, K.; Furuhjelm, C.; Falth-Magnusson, K.; Fageras, M.; Duchen, K. High levels of omega-3 fatty acids in milk from omega-3 fatty acid-supplemented mothers are related to less immunoglobulin e-associated disease in infancy. Acta Paediatr. 2016, 105, 1337–1347. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.L.; Lai, S.H.; Yeh, K.W.; Huang, Y.L.; Yao, T.C.; Tsai, M.H.; Hua, M.C.; Huang, J.L.; Study, P.C. Exclusive breastfeeding is associated with reduced cow’s milk sensitization in early childhood. Pediatr. Allergy Immunol. 2014, 25, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Tran, M.M.; Lefebvre, D.L.; Dai, D.; Dharma, C.; Subbarao, P.; Lou, W.; Azad, M.B.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; et al. Timing of food introduction and development of food sensitization in a prospective birth cohort. Pediatr. Allergy Immunol. 2017, 28, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Jepsen, A.A.; Chawes, B.L.; Carson, C.G.; Schoos, A.M.; Thysen, A.H.; Waage, J.; Brix, S.; Bisgaard, H. High breast milk il-1beta level is associated with reduced risk of childhood eczema. Clin. Exp. Allergy 2016, 46, 1344–1354. [Google Scholar] [CrossRef] [PubMed]
- Van Meel, E.R.; de Jong, M.; Elbert, N.J.; den Dekker, H.T.; Reiss, I.K.; de Jongste, J.C.; Jaddoe, V.W.V.; Duijts, L. Duration and exclusiveness of breastfeeding and school-age lung function and asthma. Ann. Allergy Asthma Immunol. 2017, 119, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Logan, C.A.; Brandt, S.; Wabitsch, M.; Brenner, H.; Wiens, F.; Stahl, B.; Marosvolgyi, T.; Decsi, T.; Rothenbacher, D.; Genuneit, J. New approach shows no association between maternal milk fatty acid composition and childhood wheeze or asthma. Allergy 2017, 72, 1374–1383. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Vehling, L.; Lu, Z.; Dai, D.; Subbarao, P.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Lefebvre, D.L.; Sears, M.R.; et al. Breastfeeding, maternal asthma and wheezing in the first year of life: A longitudinal birth cohort study. Eur. Respir. J. 2017, 49, 1602019. [Google Scholar] [CrossRef] [PubMed]
- North, M.L.; Brook, J.R.; Lee, E.Y.; Omana, V.; Daniel, N.M.; Steacy, L.M.; Evans, G.J.; Diamond, M.L.; Ellis, A.K. The kingston allergy birth cohort: Exploring parentally reported respiratory outcomes through the lens of the exposome. Ann. Allergy Asthma Immunol. 2017, 118, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Kashanian, M.; Mohtashami, S.S.; Bemanian, M.H.; Moosavi, S.A.J.; Moradi Lakeh, M. Evaluation of the associations between childhood asthma and prenatal and perinatal factors. Int. J. Gynaecol. Obstet. 2017, 137, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Du, R.; Zeiger, A.M.; McGarry, M.E.; Hu, D.; Thakur, N.; Pino-Yanes, M.; Galanter, J.M.; Farber, H.J.; Eng, C.; et al. Breastfeeding associated with higher lung function in african american youths with asthma. J. Asthma 2016. [Google Scholar] [CrossRef] [PubMed]
- Arif, A.A.; Racine, E.F. Does longer duration of breastfeeding prevent childhood asthma in low-income families? J. Asthma 2016, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Den Dekker, H.T.; Sonnenschein-van der Voort, A.M.; Jaddoe, V.W.; Reiss, I.K.; de Jongste, J.C.; Duijts, L. Breastfeeding and asthma outcomes at the age of 6 years. The generation r study. Pediatr. Allergy Immunol. 2016, 27, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Kull, I.; Wickman, M.; Lilja, G.; Nordvall, S.L.; Pershagen, G. Breast feeding and allergic diseases in infants-a prospective birth cohort study. Arch. Dis. Child. 2002, 87, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Matheson, M.C.; Erbas, B.; Balasuriya, A.; Jenkins, M.A.; Wharton, C.L.; Tang, M.L.; Abramson, M.J.; Walters, E.H.; Hopper, J.L.; Dharmage, S.C. Breast-feeding and atopic disease: A cohort study from childhood to middle age. J. Allergy Clin. Immunol. 2007, 120, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.W.; Dakin, C.J.; O’Callaghan, M.J. Breastfeeding does not increase the risk of asthma at 14 years. Pediatrics 2006, 117, e787–e792. [Google Scholar] [CrossRef] [PubMed]
- Gdalevich, M.; Mimouni, D.; David, M.; Mimouni, M. Breast-feeding and the onset of atopic dermatitis in childhood: A systematic review and meta-analysis of prospective studies. J. Am. Acad. Dermatol. 2001, 45, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Gdalevich, M.; Mimouni, D.; Mimouni, M. Breast-feeding and the risk of bronchial asthma in childhood: A systematic review with meta-analysis of prospective studies. J. Pediatr. 2001, 139, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Horta, B.; Victora, C. Long-Term Effects of Breastfeeding: A Systematic Review; WHO: Geneva, Switzerland, 2013. [Google Scholar]
- Dogaru, C.M.; Nyffenegger, D.; Pescatore, A.M.; Spycher, B.D.; Kuehni, C.E. Breastfeeding and childhood asthma: Systematic review and meta-analysis. Am. J. Epidemiol. 2014, 179, 1153–1167. [Google Scholar] [CrossRef] [PubMed]
- Ip, S.; Chung, M.; Raman, G.; Chew, P.; Magula, N.; DeVine, D.; Trikalinos, T.; Lau, J. Breastfeeding and maternal and infant health outcomes in developed countries. Evid. Rep. Technol. Assess. 2007, 153, 1–186. [Google Scholar]
- Lodge, C.J.; Tan, D.J.; Lau, M.X.; Dai, X.; Tham, R.; Lowe, A.J.; Bowatte, G.; Allen, K.J.; Dharmage, S.C. Breastfeeding and asthma and allergies: A systematic review and meta-analysis. Acta Paediatr. 2015, 104, 38–53. [Google Scholar] [CrossRef] [PubMed]
- Kramer, M.S.; Chalmers, B.; Hodnett, E.D.; Sevkovskaya, Z.; Dzikovich, I.; Shapiro, S.; Collet, J.P.; Vanilovich, I.; Mezen, I.; Ducruet, T.; et al. Promotion of breastfeeding intervention trial (probit): A randomized trial in the republic of belarus. J. Am. Med. Assoc. 2001, 285, 413–420. [Google Scholar] [CrossRef]
- Kramer, M.S.; Matush, L.; Vanilovich, I.; Platt, R.; Bogdanovich, N.; Sevkovskaya, Z.; Dzikovich, I.; Shishko, G.; Mazer, B.; Promotion of Breastfeeding Intervention Trial Study Group. Effect of prolonged and exclusive breast feeding on risk of allergy and asthma: Cluster randomised trial. BMJ 2007, 335, 815. [Google Scholar] [CrossRef] [PubMed]
- Tomicic, S.; Johansson, G.; Voor, T.; Bjorksten, B.; Bottcher, M.F.; Jenmalm, M.C. Breast milk cytokine and iga composition differ in estonian and swedish mothers-relationship to microbial pressure and infant allergy. Pediatr. Res. 2010, 68, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Bjorksten, B.; Ait-Khaled, N.; Innes Asher, M.; Clayton, T.O.; Robertson, C.; ISAAC Phase Three Study Group. Global analysis of breast feeding and risk of symptoms of asthma, rhinoconjunctivitis and eczema in 6–7 year old children: Isaac phase three. Allergol. Immunopathol. 2011, 39, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Saarinen, U.M.; Kajosaari, M. Breastfeeding as prophylaxis against atopic disease: Prospective follow-up study until 17 years old. Lancet 1995, 346, 1065–1069. [Google Scholar] [CrossRef]
- Kull, I.; Melen, E.; Alm, J.; Hallberg, J.; Svartengren, M.; van Hage, M.; Pershagen, G.; Wickman, M.; Bergstrom, A. Breast-feeding in relation to asthma, lung function, and sensitization in young schoolchildren. J. Allergy Clin. Immunol. 2010, 125, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Lucas, A.; Brooke, O.G.; Morley, R.; Cole, T.J.; Bamford, M.F. Early diet of preterm infants and development of allergic or atopic disease: Randomised prospective study. BMJ 1990, 300, 837–840. [Google Scholar] [CrossRef] [PubMed]
- Pesonen, M.; Kallio, M.J.; Ranki, A.; Siimes, M.A. Prolonged exclusive breastfeeding is associated with increased atopic dermatitis: A prospective follow-up study of unselected healthy newborns from birth to age 20 years. Clin. Exp. Allergy 2006, 36, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Mihrshahi, S.; Ampon, R.; Webb, K.; Almqvist, C.; Kemp, A.S.; Hector, D.; Marks, G.B.; Team, C. The association between infant feeding practices and subsequent atopy among children with a family history of asthma. Clin. Exp. Allergy 2007, 37, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Suzue, J.; Kaji, H. Breastfeeding reduces the severity of respiratory syncytial virus infection among young infants: A multi-center prospective study. Pediatr. Int. 2009, 51, 812–816. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.L.; Griggs, K.M.; Forsyth, K.D.; Bersten, A.D. Lower interleukin-8 levels in airway aspirates from breastfed infants with acute bronchiolitis. Pediatr. Allergy Immunol. 2010, 21, e691–e696. [Google Scholar] [CrossRef] [PubMed]
- Brew, B.K.; Allen, C.W.; Toelle, B.G.; Marks, G.B. Systematic review and meta-analysis investigating breast feeding and childhood wheezing illness. Paediatr. Perinat. Epidemiol. 2011, 25, 507–518. [Google Scholar] [CrossRef] [PubMed]
- De Benedictis, F.M.; Bush, A. Infantile wheeze: Rethinking dogma. Arch. Dis. Child. 2017, 102, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Scholtens, S.; Wijga, A.H.; Brunekreef, B.; Kerkhof, M.; Hoekstra, M.O.; Gerritsen, J.; Aalberse, R.; de Jongste, J.C.; Smit, H.A. Breast feeding, parental allergy and asthma in children followed for 8 years. The piama birth cohort study. Thorax 2009, 64, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Nagel, G.; Buchele, G.; Weinmayr, G.; Bjorksten, B.; Chen, Y.Z.; Wang, H.; Nystad, W.; Saraclar, Y.; Braback, L.; Batlles-Garrido, J.; et al. Effect of breastfeeding on asthma, lung function and bronchial hyperreactivity in isaac phase ii. Eur. Respir. J. 2009, 33, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Klein, L.; Kyewski, B.; Allen, P.M.; Hogquist, K.A. Positive and negative selection of the t cell repertoire: What thymocytes see (and don’t see). Nat. Rev. Immunol. 2014, 14, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S. Naturally arising foxp3-expressing cd25+cd4+ regulatory t cells in immunological tolerance to self and non-self. Nat. Immunol. 2005, 6, 345–352. [Google Scholar] [CrossRef] [PubMed]
- McLean-Tooke, A.; Spickett, G.P.; Gennery, A.R. Immunodeficiency and autoimmunity in 22q11.2 deletion syndrome. Scand. J. Immunol. 2007, 66, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Shanley, D.P.; Aw, D.; Manley, N.R.; Palmer, D.B. An evolutionary perspective on the mechanisms of immunosenescence. Trends Immunol. 2009, 30, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Lynch, H.E.; Goldberg, G.L.; Chidgey, A.; Van den Brink, M.R.; Boyd, R.; Sempowski, G.D. Thymic involution and immune reconstitution. Trends Immunol. 2009, 30, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, N.; Eviston, D.P.; Quinton, A.E.; Benzie, R.J.; Kirby, A.C.; Peek, M.J.; Nanan, R.K. Smaller fetal thymuses in pre-eclampsia: A prospective cross-sectional study. Ultrasound Obstet. Gynecol. 2011, 37, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Gerpe, L.; Rey-Mendez, M. Evolution of the thymus size in response to physiological and random events throughout life. Microsc. Res. Tech. 2003, 62, 464–476. [Google Scholar] [CrossRef] [PubMed]
- Hasselbalch, H.; Jeppesen, D.L.; Engelmann, M.D.; Michaelsen, K.F.; Nielsen, M.B. Decreased thymus size in formula-fed infants compared with breastfed infants. Acta Paediatr. 1996, 85, 1029–1032. [Google Scholar] [CrossRef] [PubMed]
- Hasselbalch, H.; Engelmann, M.D.; Ersboll, A.K.; Jeppesen, D.L.; Fleischer-Michaelsen, K. Breast-feeding influences thymic size in late infancy. Eur. J. Pediatr. 1999, 158, 964–967. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, D.L.; Hasselbalch, H.; Lisse, I.M.; Ersboll, A.K.; Engelmann, M.D. T-lymphocyte subsets, thymic size and breastfeeding in infancy. Pediatr. Allergy Immunol. 2004, 15, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Eysteinsdottir, J.H.; Freysdottir, J.; Haraldsson, A.; Stefansdottir, J.; Skaftadottir, I.; Helgason, H.; Ogmundsdottir, H.M. The influence of partial or total thymectomy during open heart surgery in infants on the immune function later in life. Clin. Exp. Immunol. 2004, 136, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Ngom, P.T.; Collinson, A.C.; Pido-Lopez, J.; Henson, S.M.; Prentice, A.M.; Aspinall, R. Improved thymic function in exclusively breastfed infants is associated with higher interleukin 7 concentrations in their mothers’ breast milk. Am. J. Clin. Nutr. 2004, 80, 722–728. [Google Scholar] [PubMed]
- Candeias, S.; Muegge, K.; Durum, S.K. Il-7 receptor and vdj recombination: Trophic versus mechanistic actions. Immunity 1997, 6, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Pannaraj, P.S.; Li, F.; Cerini, C.; Bender, J.M.; Yang, S.; Rollie, A.; Adisetiyo, H.; Zabih, S.; Lincez, P.J.; Bittinger, K.; et al. Association between breast milk bacterial communities and establishment and development of the infant gut microbiome. JAMA Pediatr. 2017, 171, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Peroni, D.G.; Pescollderungg, L.; Piacentini, G.L.; Rigotti, E.; Maselli, M.; Watschinger, K.; Piazza, M.; Pigozzi, R.; Boner, A.L. Immune regulatory cytokines in the milk of lactating women from farming and urban environments. Pediatr. Allergy Immunol. 2010, 21, 977–982. [Google Scholar] [CrossRef] [PubMed]
- Amoudruz, P.; Holmlund, U.; Schollin, J.; Sverremark-Ekstrom, E.; Montgomery, S.M. Maternal country of birth and previous pregnancies are associated with breast milk characteristics. Pediatr. Allergy Immunol. 2009, 20, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Holmlund, U.; Amoudruz, P.; Johansson, M.A.; Haileselassie, Y.; Ongoiba, A.; Kayentao, K.; Traore, B.; Doumbo, S.; Schollin, J.; Doumbo, O.; et al. Maternal country of origin, breast milk characteristics and potential influences on immunity in offspring. Clin. Exp. Immunol. 2010, 162, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Wang, G.; Liu, X.; Kumar, R.; Tsai, H.J.; Arguelles, L.; Hao, K.; Pearson, C.; Ortiz, K.; Bonzagni, A.; et al. Gene polymorphisms, breast-feeding, and development of food sensitization in early childhood. J. Allergy Clin. Immunol. 2011, 128, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Kramer, M.S.; Kakuma, R. Optimal duration of exclusive breastfeeding. Cochrane Database Syst. Rev. 2012, 8, CD003517. [Google Scholar]
- Koletzko, B.; Lien, E.; Agostoni, C.; Bohles, H.; Campoy, C.; Cetin, I.; Decsi, T.; Dudenhausen, J.W.; Dupont, C.; Forsyth, S.; et al. The roles of long-chain polyunsaturated fatty acids in pregnancy, lactation and infancy: Review of current knowledge and consensus recommendations. J. Perinat. Med. 2008, 36, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Hunt, K.M.; Foster, J.A.; Forney, L.J.; Schutte, U.M.; Beck, D.L.; Abdo, Z.; Fox, L.K.; Williams, J.E.; McGuire, M.K.; McGuire, M.A. Characterization of the diversity and temporal stability of bacterial communities in human milk. PLoS ONE 2011, 6, e21313. [Google Scholar] [CrossRef] [PubMed]
- Takahata, Y.; Takada, H.; Nomura, A.; Ohshima, K.; Nakayama, H.; Tsuda, T.; Nakano, H.; Hara, T. Interleukin-18 in human milk. Pediatr. Res. 2001, 50, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Ustundag, B.; Yilmaz, E.; Dogan, Y.; Akarsu, S.; Canatan, H.; Halifeoglu, I.; Cikim, G.; Aygun, A.D. Levels of cytokines (il-1beta, il-2, il-6, il-8, tnf-alpha) and trace elements (zn, cu) in breast milk from mothers of preterm and term infants. Mediat. Inflamm. 2005, 2005, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Rigotti, E.; Piacentini, G.L.; Ress, M.; Pigozzi, R.; Boner, A.L.; Peroni, D.G. Transforming growth factor-beta and interleukin-10 in breast milk and development of atopic diseases in infants. Clin. Exp. Allergy 2006, 36, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Munblit, D.; Treneva, M.; Peroni, D.G.; Colicino, S.; Chow, L.; Dissanayeke, S.; Abrol, P.; Sheth, S.; Pampura, A.; Boner, A.L.; et al. Colostrum and mature human milk of women from london, moscow, and verona: Determinants of immune composition. Nutrients 2016, 8, 695. [Google Scholar] [CrossRef] [PubMed]
- Minniti, F.; Comberiati, P.; Munblit, D.; Piacentini, G.L.; Antoniazzi, E.; Zanoni, L.; Boner, A.L.; Peroni, D.G. Breast-milk characteristics protecting against allergy. Endocr. Metab. Immune Disord. Drug Targets 2014, 14, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Munblit, D.; Boyle, R.J.; Warner, J.O. Factors affecting breast milk composition and potential consequences for development of the allergic phenotype. Clin. Exp. Allergy 2015, 45, 583–601. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, J.S.; Bryan, D.L.; Neumann, M.A.; Makrides, M.; Gibson, R.A. Transforming growth factor beta in human milk does not change in response to modest intakes of docosahexaenoic acid. Lipids 2001, 36, 1179–1181. [Google Scholar] [CrossRef] [PubMed]
- Dunstan, J.A.; Roper, J.; Mitoulas, L.; Hartmann, P.E.; Simmer, K.; Prescott, S.L. The effect of supplementation with fish oil during pregnancy on breast milk immunoglobulin a, soluble cd14, cytokine levels and fatty acid composition. Clin. Exp. Allergy 2004, 34, 1237–1242. [Google Scholar] [CrossRef] [PubMed]
- Urwin, H.J.; Miles, E.A.; Noakes, P.S.; Kremmyda, L.S.; Vlachava, M.; Diaper, N.D.; Perez-Cano, F.J.; Godfrey, K.M.; Calder, P.C.; Yaqoob, P. Salmon consumption during pregnancy alters fatty acid composition and secretory iga concentration in human breast milk. J. Nutr. 2012, 142, 1603–1610. [Google Scholar] [CrossRef] [PubMed]
- Bottcher, M.F.; Abrahamsson, T.R.; Fredriksson, M.; Jakobsson, T.; Bjorksten, B. Low breast milk tgf-beta2 is induced by lactobacillus reuteri supplementation and associates with reduced risk of sensitization during infancy. Pediatr. Allergy Immunol. 2008, 19, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Prescott, S.L.; Wickens, K.; Westcott, L.; Jung, W.; Currie, H.; Black, P.N.; Stanley, T.V.; Mitchell, E.A.; Fitzharris, P.; Siebers, R.; et al. Supplementation with lactobacillus rhamnosus or bifidobacterium lactis probiotics in pregnancy increases cord blood interferon-gamma and breast milk transforming growth factor-beta and immunoglobin a detection. Clin. Exp. Allergy 2008, 38, 1606–1614. [Google Scholar] [CrossRef] [PubMed]
- Boyle, R.J.; Ismail, I.H.; Kivivuori, S.; Licciardi, P.V.; Robins-Browne, R.M.; Mah, L.J.; Axelrad, C.; Moore, S.; Donath, S.; Carlin, J.B.; et al. Lactobacillus gg treatment during pregnancy for the prevention of eczema: A randomized controlled trial. Allergy 2011, 66, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Hoppu, U.; Isolauri, E.; Laakso, P.; Matomaki, J.; Laitinen, K. Probiotics and dietary counselling targeting maternal dietary fat intake modifies breast milk fatty acids and cytokines. Eur. J. Nutr. 2012, 51, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Kuitunen, M.; Kukkonen, A.K.; Savilahti, E. Impact of maternal allergy and use of probiotics during pregnancy on breast milk cytokines and food antibodies and development of allergy in children until 5 years. Int. Arch. Allergy Immunol. 2012, 159, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Savilahti, E.M.; Kukkonen, A.K.; Kuitunen, M.; Savilahti, E. Soluble cd14, alpha-and beta-defensins in breast milk: Association with the emergence of allergy in a high-risk population. Innate Immun. 2015, 21, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Linnamaa, P.; Nieminen, K.; Koulu, L.; Tuomasjukka, S.; Kallio, H.; Yang, B.; Tahvonen, R.; Savolainen, J. Black currant seed oil supplementation of mothers enhances ifn-gamma and suppresses il-4 production in breast milk. Pediatr. Allergy Immunol. 2013, 24, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Nikniaz, L.; Ostadrahimi, A.; Mahdavi, R.; Hejazi, M.A.; Salekdeh, G.H. Effects of synbiotic supplementation on breast milk levels of iga, tgf-beta1, and tgf-beta2. J. Hum. Lact. 2013, 29, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Kalliomaki, M.; Ouwehand, A.; Arvilommi, H.; Kero, P.; Isolauri, E. Transforming growth factor-beta in breast milk: A potential regulator of atopic disease at an early age. J. Allergy Clin. Immunol. 1999, 104, 1251–1257. [Google Scholar] [CrossRef]
- Jones, C.A.; Holloway, J.A.; Popplewell, E.J.; Diaper, N.D.; Holloway, J.W.; Vance, G.H.; Warner, J.A.; Warner, J.O. Reduced soluble cd14 levels in amniotic fluid and breast milk are associated with the subsequent development of atopy, eczema, or both. J. Allergy Clin. Immunol. 2002, 109, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Bottcher, M.F.; Jenmalm, M.C.; Bjorksten, B. Cytokine, chemokine and secretory iga levels in human milk in relation to atopic disease and iga production in infants. Pediatr. Allergy Immunol. 2003, 14, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Oddy, W.H.; Halonen, M.; Martinez, F.D.; Lohman, I.C.; Stern, D.A.; Kurzius-Spencer, M.; Guerra, S.; Wright, A.L. Tgf-beta in human milk is associated with wheeze in infancy. J. Allergy Clin. Immunol. 2003, 112, 723–728. [Google Scholar] [CrossRef]
- Savilahti, E.; Siltanen, M.; Kajosaari, M.; Vaarala, O.; Saarinen, K.M. Iga antibodies, tgf-beta1 and -beta2, and soluble cd14 in the colostrum and development of atopy by age 4. Pediatr. Res. 2005, 58, 1300–1305. [Google Scholar] [CrossRef] [PubMed]
- Snijders, B.E.; Damoiseaux, J.G.; Penders, J.; Kummeling, I.; Stelma, F.F.; van Ree, R.; van den Brandt, P.A.; Thijs, C. Cytokines and soluble cd14 in breast milk in relation with atopic manifestations in mother and infant (koala study). Clin. Exp. Allergy 2006, 36, 1609–1615. [Google Scholar] [CrossRef] [PubMed]
- Soto-Ramirez, N.; Karmaus, W.; Yousefi, M.; Zhang, H.; Liu, J.; Gangur, V. Maternal immune markers in serum during gestation and in breast milk and the risk of asthma-like symptoms at ages 6 and 12 months: A longitudinal study. Allergy Asthma Clin. Immunol. 2012, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Ismail, I.H.; Licciardi, P.V.; Oppedisano, F.; Boyle, R.J.; Tang, M.L. Relationship between breast milk scd14, tgf-beta1 and total iga in the first month and development of eczema during infancy. Pediatr. Allergy Immunol. 2013, 24, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Orivuori, L.; Loss, G.; Roduit, C.; Dalphin, J.C.; Depner, M.; Genuneit, J.; Lauener, R.; Pekkanen, J.; Pfefferle, P.; Riedler, J.; et al. Soluble immunoglobulin a in breast milk is inversely associated with atopic dermatitis at early age: The pasture cohort study. Clin. Exp. Allergy 2014, 44, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Munblit, D.; Treneva, M.; Peroni, D.G.; Colicino, S.; Chow, L.Y.; Dissanayeke, S.; Pampura, A.; Boner, A.L.; Geddes, D.T.; Boyle, R.J.; et al. Immune components in human milk are associated with early infant immunological health outcomes: A prospective three-country analysis. Nutrients 2017, 9, 532. [Google Scholar] [CrossRef] [PubMed]
- Oddy, W.H.; Rosales, F. A systematic review of the importance of milk tgf-beta on immunological outcomes in the infant and young child. Pediatr. Allergy Immunol. 2010, 21, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, J.; Sasahara, A.; Yoshida, T.; Sira, M.M.; Futatani, T.; Kanegane, H.; Miyawaki, T. Role of transforming growth factor-beta in breast milk for initiation of iga production in newborn infants. Early Hum. Dev. 2004, 77, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Ulevitch, R.J.; Tobias, P.S. Receptor-dependent mechanisms of cell stimulation by bacterial endotoxin. Annu. Rev. Immunol. 1995, 13, 437–457. [Google Scholar] [CrossRef] [PubMed]
- Pugin, J.; Heumann, I.D.; Tomasz, A.; Kravchenko, V.V.; Akamatsu, Y.; Nishijima, M.; Glauser, M.P.; Tobias, P.S.; Ulevitch, R.J. Cd14 is a pattern recognition receptor. Immunity 1994, 1, 509–516. [Google Scholar] [CrossRef]
- Holt, P.G.; Sly, P.D.; Bjorksten, B. Atopic versus infectious diseases in childhood: A question of balance? Pediatr. Allergy Immunol. 1997, 8, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Labeta, M.O.; Vidal, K.; Nores, J.E.; Arias, M.; Vita, N.; Morgan, B.P.; Guillemot, J.C.; Loyaux, D.; Ferrara, P.; Schmid, D.; et al. Innate recognition of bacteria in human milk is mediated by a milk-derived highly expressed pattern recognition receptor, soluble cd14. J. Exp. Med. 2000, 191, 1807–1812. [Google Scholar] [CrossRef] [PubMed]
- Vidal, K.; Labeta, M.O.; Schiffrin, E.J.; Donnet-Hughes, A. Soluble cd14 in human breast milk and its role in innate immune responses. Acta Odontol. Scand. 2001, 59, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Jarvinen, K.M.; Suarez-Farinas, M.; Savilahti, E.; Sampson, H.A.; Berin, M.C. Immune factors in breast milk related to infant milk allergy are independent of maternal atopy. J. Allergy Clin. Immunol. 2015, 135, 1390–1393. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.; Kuhn, L.; Ghosh, M.K.; Kankasa, C.; Semrau, K.; Sinkala, M.; Mwiya, M.; Thea, D.M.; Aldrovandi, G.M. Low and undetectable breast milk interleukin-7 concentrations are associated with reduced risk of postnatal hiv transmission. J. Acquir. Immune Defic. Syndr. 2007, 46, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Castellote, C.; Casillas, R.; Ramirez-Santana, C.; Perez-Cano, F.J.; Castell, M.; Moretones, M.G.; Lopez-Sabater, M.C.; Franch, A. Premature delivery influences the immunological composition of colostrum and transitional and mature human milk. J. Nutr. 2011, 141, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Zuccotti, G.; Meneghin, F.; Aceti, A.; Barone, G.; Callegari, M.L.; Di Mauro, A.; Fantini, M.P.; Gori, D.; Indrio, F.; Maggio, L.; et al. Probiotics for prevention of atopic diseases in infants: Systematic review and meta-analysis. Allergy 2015, 70, 1356–1371. [Google Scholar] [CrossRef] [PubMed]
- Kondo, N.; Suda, Y.; Nakao, A.; Oh-Oka, K.; Suzuki, K.; Ishimaru, K.; Sato, M.; Tanaka, T.; Nagai, A.; Yamagata, Z. Maternal psychosocial factors determining the concentrations of transforming growth factor-beta in breast milk. Pediatr. Allergy Immunol. 2011, 22, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Rautava, S.; Kalliomaki, M.; Isolauri, E. Probiotics during pregnancy and breast-feeding might confer immunomodulatory protection against atopic disease in the infant. J. Allergy Clin. Immunol. 2002, 109, 119–121. [Google Scholar] [CrossRef] [PubMed]
- Osborn, D.A.; Sinn, J.K. Prebiotics in infants for prevention of allergy. Cochrane Database Syst. Rev. 2013, CD006474. [Google Scholar] [CrossRef]
- Dotterud, C.K.; Storro, O.; Johnsen, R.; Oien, T. Probiotics in pregnant women to prevent allergic disease: A randomized, double-blind trial. Br. J. Dermatol. 2010, 163, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Stahl, B.; Folkerts, G.; Garssen, J.; van’t-Land, B. The immunological benefits for complex oligosaccharides in human milk. In Nutrition, Immunity & Infection; CRC Press: Boca Raton, FL, USA, 2017; in press. [Google Scholar]
- Thurl, S.; Munzert, M.; Henker, J.; Boehm, G.; Muller-Werner, B.; Jelinek, J.; Stahl, B. Variation of human milk oligosaccharides in relation to milk groups and lactational periods. Br. J. Nutr. 2010, 104, 1261–1271. [Google Scholar] [CrossRef] [PubMed]
- Erney, R.M.; Malone, W.T.; Skelding, M.B.; Marcon, A.A.; Kleman-Leyer, K.M.; O’Ryan, M.L.; Ruiz-Palacios, G.; Hilty, M.D.; Pickering, L.K.; Prieto, P.A. Variability of human milk neutral oligosaccharides in a diverse population. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, P.; Warren, C.D.; Altaye, M.; Morrow, A.L.; Ruiz-Palacios, G.; Pickering, L.K.; Newburg, D.S. Fucosylated human milk oligosaccharides vary between individuals and over the course of lactation. Glycobiology 2001, 11, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Morrow, A.L.; Meinzen-Derr, J.; Huang, P.; Schibler, K.R.; Cahill, T.; Keddache, M.; Kallapur, S.G.; Newburg, D.S.; Tabangin, M.; Warner, B.B.; et al. Fucosyltransferase 2 non-secretor and low secretor status predicts severe outcomes in premature infants. J. Pediatr. 2011, 158, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Smilowitz, J.T.; O’Sullivan, A.; Barile, D.; German, J.B.; Lonnerdal, B.; Slupsky, C.M. The human milk metabolome reveals diverse oligosaccharide profiles. J. Nutr. 2013, 143, 1709–1718. [Google Scholar] [CrossRef] [PubMed]
- Spevacek, A.R.; Smilowitz, J.T.; Chin, E.L.; Underwood, M.A.; German, J.B.; Slupsky, C.M. Infant maturity at birth reveals minor differences in the maternal milk metabolome in the first month of lactation. J. Nutr. 2015, 145, 1698–1708. [Google Scholar] [CrossRef] [PubMed]
- Verhasselt, V.; Milcent, V.; Cazareth, J.; Kanda, A.; Fleury, S.; Dombrowicz, D.; Glaichenhaus, N.; Julia, V. Breast milk-mediated transfer of an antigen induces tolerance and protection from allergic asthma. Nat. Med. 2008, 14, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.A.; Iyengar, R.S. Breast milk, microbiota, and intestinal immune homeostasis. Pediatr. Res. 2015, 77, 220–228. [Google Scholar] [PubMed]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. Inducible foxp3+ regulatory t-cell development by a commensal bacterium of the intestinal microbiota. Proc. Natl. Acad Sci. USA 2010, 107, 12204–12209. [Google Scholar] [CrossRef] [PubMed]
- Schijf, M.A.; Kruijsen, D.; Bastiaans, J.; Coenjaerts, F.E.; Garssen, J.; van Bleek, G.M.; van’t Land, B. Specific dietary oligosaccharides increase th1 responses in a mouse respiratory syncytial virus infection model. J. Virol. 2012, 86, 11472–11482. [Google Scholar] [CrossRef] [PubMed]
- Diesner, S.C.; Bergmayr, C.; Pfitzner, B.; Assmann, V.; Krishnamurthy, D.; Starkl, P.; Endesfelder, D.; Rothballer, M.; Welzl, G.; Rattei, T.; et al. A distinct microbiota composition is associated with protection from food allergy in an oral mouse immunization model. Clin. Immunol. 2016, 173, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Hua, X.; Goedert, J.J.; Pu, A.; Yu, G.; Shi, J. Allergy associations with the adult fecal microbiota: Analysis of the american gut project. EBioMedicine 2016, 3, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Noval Rivas, M.; Burton, O.T.; Wise, P.; Zhang, Y.-Q.; Hobson, S.A.; Garcia Lloret, M.; Chehoud, C.; Kuczynski, J.; DeSantis, T.; Warrington, J.; et al. A microbiota signature associated with experimental food allergy promotes allergic sensitization and anaphylaxis. J. Allergy Clin. Immunol. 2013, 131, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Ruokolainen, L.; Paalanen, L.; Karkman, A.; Laatikainen, T.; von Hertzen, L.; Vlasoff, T.; Markelova, O.; Masyuk, V.; Auvinen, P.; Paulin, L.; et al. Significant disparities in allergy prevalence and microbiota between the young people in finnish and russian karelia. Clin. Exp. Allergy 2017, 47, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-L.; Chen, X.; Zheng, P.-Y.; Luo, Y.; Lu, G.-F.; Liu, Z.-Q.; Huang, H.; Yang, P.-C. Oral bifidobacterium modulates intestinal immune inflammation in mice with food allergy. J. Gastroenterol. Hepatol. 2010, 25, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef] [PubMed]
- An, D.; Oh, S.F.; Olszak, T.; Neves, J.F.; Avci, F.Y. Sphingolipids from a symbiotic microbe regulate homeostasis of host intestinal natural killer t cells. Cell 2014, 156, 123–133. [Google Scholar] [CrossRef] [PubMed]
- El Aidy, S.; Hooiveld, G.; Tremaroli, V.; Backhed, F.; Kleerebezem, M. The gut microbiota and mucosal homeostasis. Gut Microbes 2014, 4, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Olszak, T.; An, D.; Zeissig, S.; Vera, M.P.; Richter, J.; Franke, A.; Glickman, J.N.; Siebert, R.; Baron, R.M.; Kasper, D.L.; et al. Microbial exposure during early life has persistent effects on natural killer t cell function. Science 2012, 336, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Marcobal, A.; Barboza, M.; Froehlich, J.W.; Block, D.E.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Consumption of human milk oligosaccharides by gut-related microbes. J. Agric. Food Chem. 2010, 58, 5334–5340. [Google Scholar] [CrossRef] [PubMed]
- De Leoz, M.L.A.; Kalanetra, K.M.; Bokulich, N.A.; Strum, J.S.; Underwood, M.A.; German, J.B.; Mills, D.A.; Lebrilla, C.B. Human milk glycomics and gut microbial genomics in infant feces show a correlation between human milk oligosaccharides and gut microbiota: A proof-of-concept study. J. Proteome Res. 2015, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Garrido, D.; Barile, D.; Mills, D.A. A molecular basis for bifidobacterial enrichment in the infant gastrointestinal tract. Adv. Nutr. 2012, 3, 415S–421S. [Google Scholar] [CrossRef] [PubMed]
- LoCascio, R.G.; Ninonuevo, M.R.; Freeman, S.L.; Sela, D.A.; Grimm, R.; Lebrilla, C.B.; Mills, D.A.; German, J.B. Glycoprofiling of bifidobacterial consumption of human milk oligosaccharides demonstrates strain specific, preferential consumption of small chain glycans secreted in early human lactation. J. Agric. Food Chem. 2007, 55, 8914–8919. [Google Scholar] [CrossRef] [PubMed]
- Marcobal, A.; Sonnenburg, J.L. Human milk oligosaccharide consumption by intestinal microbiota. Clin. Microbiol. Infect. 2012, 18, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.E.; Ni onuevo, M.; Mills, D.A.; Lebrilla, C.B.; German, J.B. In vitro fermentability of human milk oligosaccharides by several strains of bifidobacteria. Mol. Nutr. Food Res. 2007, 51, 1398–1405. [Google Scholar] [CrossRef] [PubMed]
- Boudry, G.; Hamilton, M.K.; Chichlowski, M.; Wickramasinghe, S.; Barile, D.; Kalanetra, K.M.; Mills, D.A.; Raybould, H.E. Bovine milk oligosaccharides decrease gut permeability and improve inflammation and microbial dysbiosis in diet-induced obese mice. J. Dairy Sci. 2017, 100, 2471–2481. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-Y.; Yang, Z.-Y.; Dai, W.-K.; Huang, J.-Q.; Li, Y.-H.; Zhang, J.; Qiu, C.-Z.; Wei, C.; Zhou, Q.; Sun, X.; et al. Protective effect of Bifidobacterium infantis cgmcc313-2 on ovalbumin-induced airway asthma and β-lactoglobulin-induced intestinal food allergy mouse models. World J. Gastroenterol. 2017, 23, 2149–2158. [Google Scholar] [CrossRef] [PubMed]
- Schouten, B.; van Esch, B.C.; Hofman, G.A.; van Doorn, S.A.; Knol, J.; Nauta, A.J.; Garssen, J.; Willemsen, L.E.; Knippels, L.M. Cow milk allergy symptoms are reduced in mice fed dietary synbiotics during oral sensitization with whey. J. Nutr. 2009, 139, 1398–1403. [Google Scholar] [CrossRef] [PubMed]
- Lewis, Z.T.; Totten, S.M.; Smilowitz, J.T.; Popovic, M.; Parker, E.; Lemay, D.G.; Van Tassell, M.L.; Miller, M.J.; Jin, Y.-S.; German, J.B.; et al. Maternal fucosyltransferase 2 status affects the gut bifidobacterial communities of breastfed infants. Microbiome 2015, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, D.L.; Doster, R.S.; Weitkamp, J.-H.; Aronoff, D.; Gaddy, J.A.; Townsend, S.D. Human milk oligosaccharides exhibit antimicrobial and anti-biofilm properties against group b streptococcus. ACS Infect. Dis. 2017, 3, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Bode, L. The functional biology of human milk oligosaccharides. Early Hum. Dev. 2015, 91, 619–622. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.E.; Autran, C.A.; Szyszka, A.; Escajadillo, T.; Huang, M.; Godula, K.; Prudden, A.R.; Boons, G.-J.; Lewis, A.L.; Doran, K.S.; et al. Human milk oligosaccharides inhibit growth of group B streptococcus. J. Biol. Chem. 2017. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, A.J.; Koplin, J.J.; Lowe, A.J.; Tang, M.L.; Matheson, M.C.; Robinson, M.; Peters, R.; Dharmage, S.C.; Allen, K.J. Formula and breast feeding in infant food allergy: A population-based study. J. Paediatr. Child Health 2016, 52, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Hoyt, A.E.W.; Medico, T.; Commins, S.P. Breast milk and food allergy: Connections and current recommendations. Pediatr. Clin. N. Am. 2015, 62, 1493–1507. [Google Scholar] [CrossRef] [PubMed]
- Chua, M.C.; Ben-Amor, K.; Lay, C.; Neo, A.G.E.; Chiang, W.C.; Rao, R.; Chew, C.; Chaithongwongwatthana, S.; Khemapech, N.; Knol, J.; et al. Effect of synbiotic on the gut microbiota of caesarean delivered infants: A randomized, double-blind, multicenter study. J. Pediatr. Gastroenterol. Nutr. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ley, D.; Desseyn, J.L.; Mischke, M.; Knol, J.; Turck, D.; Gottrand, F. Early-life origin of intestinal inflammatory disorders. Nutr. Rev. 2017, 75, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Naarding, M.A.; Ludwig, I.S.; Groot, F.; Berkhout, B.; Geijtenbeek, T.B.; Pollakis, G.; Paxton, W.A. Lewis x component in human milk binds dc-sign and inhibits hiv-1 transfer to cd4+ t lymphocytes. J. Clin. Investig. 2005, 115, 3256–3264. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Hiller, J.; van Bergenhenegouwen, J.; Knippels, L.M.; Garssen, J.; Traidl-Hoffmann, C. In vitro evidence for immune-modulatory properties of non-digestible oligosaccharides: Direct effect on human monocyte derived dendritic cells. PLoS ONE 2015, 10, e0132304. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Lawlor, N.T.; Newburg, D.S. Human milk components modulate toll-like receptor-mediated inflammation. Adv. Nutr. 2016, 7, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Gringhuis, S.I.; Kaptein, T.M.; Wevers, B.A.; Mesman, A.W.; Geijtenbeek, T.B. Fucose-specific dc-sign signalling directs t helper cell type-2 responses via ikkepsilon- and cyld-dependent bcl3 activation. Nat. Commun. 2014, 5, 3898. [Google Scholar] [CrossRef] [PubMed]
- Noll, A.J.; Yu, Y.; Lasanajak, Y.; Duska-McEwen, G.; Buck, R.H.; Smith, D.F.; Cummings, R.D. Human dc-sign binds specific human milk glycans. Biochem. J. 2016, 473, 1343–1353. [Google Scholar] [CrossRef] [PubMed]
- De Kivit, S.; Kostadinova, A.I.; Kerperien, J.; Morgan, M.E.; Muruzabal, V.A.; Hofman, G.A.; Knippels, L.M.J.; Kraneveld, A.D.; Garssen, J.; Willemsen, L.E.M. Dietary, nondigestible oligosaccharides and bifidobacterium breve m-16v suppress allergic inflammation in intestine via targeting dendritic cell maturation. J. Leukoc. Biol. 2017, 102, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Marriage, B.J.; Buck, R.H.; Goehring, K.C.; Oliver, J.S.; Williams, J.A. Infants fed a lower calorie formula with 2’fl show growth and 2’fl uptake like breast-fed infants. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Arslanoglu, S.; Moro, G.E.; Boehm, G.; Wienz, F.; Stahl, B.; Bertino, E. Early neutral prebiotic oligosaccharide supplementation reduces the incidence of some allergic manifestations in the first 5 years of life. J. Biol. Regul. Homeost. Agents 2012, 26, 49–59. [Google Scholar] [PubMed]
- Moro, G.; Arslanoglu, S.; Stahl, B.; Jelinek, J.; Wahn, U.; Boehm, G. A mixture of prebiotic oligosaccharides reduces the incidence of atopic dermatitis during the first six months of age. Arch. Dis. Child. 2006, 91, 814–819. [Google Scholar] [CrossRef] [PubMed]
- Bollrath, J.; Powrie, F. Immunology. Feed your tregs more fiber. Science 2013, 341, 463–464. [Google Scholar] [CrossRef] [PubMed]
- Bendiks, M.; Kopp, M.V. The relationship between advances in understanding the microbiome and the maturing hygiene hypothesis. Curr. Allergy Asthma Rep. 2013, 13, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.M.; Murphy, K.; Stanton, C.; Ross, R.P.; Kober, O.I.; Juge, N.; Avershina, E.; Rudi, K.; Narbad, A.; Jenmalm, M.C.; et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb. Ecol. Health Dis. 2015, 26, 26050. [Google Scholar] [CrossRef] [PubMed]
- Bridgman, S.L.; Kozyrskyj, A.L.; Scott, J.A.; Becker, A.B.; Azad, M.B. Gut microbiota and allergic disease in children. Ann. Allergy Asthma Immunol. 2016, 116, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Guaraldi, F.; Salvatori, G. Effect of breast and formula feeding on gut microbiota shaping in newborns. Front. Cell. Infect. Microbiol. 2012, 2, 94. [Google Scholar] [CrossRef] [PubMed]
- Perez, P.F.; Dore, J.; Leclerc, M.; Levenez, F.; Benyacoub, J.; Serrant, P.; Segura-Roggero, I.; Schiffrin, E.J.; Donnet-Hughes, A. Bacterial imprinting of the neonatal immune system: Lessons from maternal cells? Pediatrics 2007, 119, e724–e732. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Langa, S.; Reviriego, C.; Jiménez, E.; Marín, M.L.; Olivares, M.; Boza, J.; Jiménez, J.; Fernández, L.; Xaus, J.; et al. The commensal microflora of human milk: New perspectives for food bacteriotherapy and probiotics. Trends Food Sci. Technol. 2004, 15, 121–127. [Google Scholar] [CrossRef]
- McGuire, M.K.; McGuire, M.A. Human milk: Mother nature’s prototypical probiotic food? Adv. Nutr. 2015, 6, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Abrahamsson, T.R.; Sinkiewicz, G.; Jakobsson, T.; Fredrikson, M.; Bjorksten, B. Probiotic lactobacilli in breast milk and infant stool in relation to oral intake during the first year of life. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Rubio, R.; Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E.; Mira, A. The human milk microbiome changes over lactation and is shaped by maternal weight and mode of delivery. Am. J. Clin. Nutr. 2012, 96, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Boix-Amoros, A.; Collado, M.C.; Mira, A. Relationship between milk microbiota, bacterial load, macronutrients, and human cells during lactation. Front. Microbiol. 2016, 7, 492. [Google Scholar] [CrossRef] [PubMed]
- Fitzstevens, J.L.; Smith, K.C.; Hagadorn, J.I.; Caimano, M.J.; Matson, A.P.; Brownell, E.A. Systematic review of the human milk microbiota. Nutr. Clin. Pract. 2017, 32, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gallego, C.; Garcia-Mantrana, I.; Salminen, S.; Collado, M.C. The human milk microbiome and factors influencing its composition and activity. Semin. Fetal Neonatal Med. 2016, 21, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Drago, L.; Toscano, M.; De Grandi, R.; Grossi, E.; Padovani, E.M.; Peroni, D.G. Microbiota network and mathematic microbe mutualism in colostrum and mature milk collected in two different geographic areas: Italy versus burundi. ISME J. 2017, 11, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Dave, V.; Street, K.; Francis, S.; Bradman, A.; Riley, L.; Eskenazi, B.; Holland, N. Bacterial microbiome of breast milk and child saliva from low-income mexican-american women and children. Pediatr. Res. 2016, 79, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Soto, A.; Martin, V.; Jimenez, E.; Mader, I.; Rodriguez, J.M.; Fernandez, L. Lactobacilli and bifidobacteria in human breast milk: Influence of antibiotherapy and other host and clinical factors. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, C.; Angelini, M.; Gloor, G.B.; Reid, G. Human milk microbiota profiles in relation to birthing method, gestation and infant gender. Microbiome 2016, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Contreras, G.A.; Rodriguez, J.M. Mastitis: Comparative etiology and epidemiology. J. Mammary Gland Biol. Neoplasia 2011, 16, 339–356. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, E.; de Andres, J.; Manrique, M.; Pareja-Tobes, P.; Tobes, R.; Martinez-Blanch, J.F.; Codoner, F.M.; Ramon, D.; Fernandez, L.; Rodriguez, J.M. Metagenomic analysis of milk of healthy and mastitis-suffering women. J. Human Lact. 2015, 31, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.M. The origin of human milk bacteria: Is there a bacterial entero-mammary pathway during late pregnancy and lactation? Adv. Nutr. 2014, 5, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Xuan, C.; Shamonki, J.M.; Chung, A.; Dinome, M.L.; Chung, M.; Sieling, P.A.; Lee, D.J. Microbial dysbiosis is associated with human breast cancer. PLoS ONE 2014, 9, e83744. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, C.; Cummins, J.; Brackstone, M.; Macklaim, J.M.; Gloor, G.B.; Baban, C.K.; Scott, L.; O’Hanlon, D.M.; Burton, J.P.; Francis, K.P.; et al. Microbiota of human breast tissue. Appl. Environ. Microbiol. 2014, 80, 3007–3014. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Maldonado-Barragan, A.; Moles, L.; Rodriguez-Banos, M.; Campo, R.D.; Fernandez, L.; Rodriguez, J.M.; Jimenez, E. Sharing of bacterial strains between breast milk and infant feces. J. Hum. Lact. 2012, 28, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Ropero, M.P.; Martin, R.; Sierra, S.; Lara-Villoslada, F.; Rodriguez, J.M.; Xaus, J.; Olivares, M. Two lactobacillus strains, isolated from breast milk, differently modulate the immune response. J. Appl. Microbiol. 2007, 102, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Gronlund, M.M.; Gueimonde, M.; Laitinen, K.; Kociubinski, G.; Gronroos, T.; Salminen, S.; Isolauri, E. Maternal breast-milk and intestinal bifidobacteria guide the compositional development of the bifidobacterium microbiota in infants at risk of allergic disease. Clin. Exp. Allergy 2007, 37, 1764–1772. [Google Scholar] [CrossRef] [PubMed]
- Dzidic, M.; Abrahamsson, T.R.; Artacho, A.; Bjorksten, B.; Collado, M.C.; Mira, A.; Jenmalm, M.C. Aberrant iga responses to the gut microbiota during infancy precede asthma and allergy development. J. Allergy Clin. Immunol. 2017, 139, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Waidyatillake, N.T.; Stoney, R.; Thien, F.; Lodge, C.J.; Simpson, J.A.; Allen, K.J.; Abramson, M.J.; Erbas, B.; Svanes, C.; Dharmage, S.C.; et al. Breast milk polyunsaturated fatty acids: Associations with adolescent allergic disease and lung function. Allergy 2017, 72, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Seppo, A.E.; Autran, C.A.; Bode, L.; Jarvinen, K.M. Human milk oligosaccharides and development of cow’s milk allergy in infants. J. Allergy Clin. Immunol. 2017, 139, 708–711. [Google Scholar] [CrossRef] [PubMed]
- Baiz, N.; Macchiaverni, P.; Tulic, M.K.; Rekima, A.; Annesi-Maesano, I.; Verhasselt, V.; EDEN Mother-Child Cohort Study Group. Early oral exposure to house dust mite allergen through breast milk: A potential risk factor for allergic sensitization and respiratory allergies in children. J. Allergy Clin. Immunol. 2017, 139, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Braun-Fahrlander, C.; von Mutius, E. Can farm milk consumption prevent allergic diseases? Clin. Exp. Allergy 2011, 41, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Barthow, C.; Wickens, K.; Stanley, T.; Mitchell, E.A.; Maude, R.; Abels, P.; Purdie, G.; Murphy, R.; Stone, P.; Kang, J.; et al. The probiotics in pregnancy study (pip study): Rationale and design of a double-blind randomised controlled trial to improve maternal health during pregnancy and prevent infant eczema and allergy. BMC Pregnancy Childbirth 2016, 16, 133. [Google Scholar] [CrossRef] [PubMed]
- Gueimonde, M.; Sakata, S.; Kalliomaki, M.; Isolauri, E.; Benno, Y.; Salminen, S. Effect of maternal consumption of lactobacillus gg on transfer and establishment of fecal bifidobacterial microbiota in neonates. J. Pediatr. Gastroenterol. Nutr. 2006, 42, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Mastromarino, P.; Capobianco, D.; Miccheli, A.; Pratico, G.; Campagna, G.; Laforgia, N.; Capursi, T.; Baldassarre, M.E. Administration of a multistrain probiotic product (vsl#3) to women in the perinatal period differentially affects breast milk beneficial microbiota in relation to mode of delivery. Pharmacol. Res. 2015, 95–96, 63–70. [Google Scholar] [PubMed]
- Niers, L.; Martin, R.; Rijkers, G.; Sengers, F.; Timmerman, H.; van Uden, N.; Smidt, H.; Kimpen, J.; Hoekstra, M. The effects of selected probiotic strains on the development of eczema (the panda study). Allergy 2009, 64, 1349–1358. [Google Scholar] [CrossRef] [PubMed]
- Rautava, S.; Kainonen, E.; Salminen, S.; Isolauri, E. Maternal probiotic supplementation during pregnancy and breast-feeding reduces the risk of eczema in the infant. J. Allergy Clin. Immunol. 2012, 130, 1355–1360. [Google Scholar] [CrossRef] [PubMed]
- Kramer, M.S.; Kakuma, R. The optimal duration of exclusive breastfeeding: A systematic review. Adv. Exp. Med. Biol. 2004, 554, 63–77. [Google Scholar] [PubMed]
- Allen, L.H. Current information gaps in micronutrient research, programs and policy: How can we fill them? World Rev. Nutr. Diet. 2016, 115, 109–117. [Google Scholar] [PubMed]
- Hampel, D.; Allen, L.H. Analyzing b-vitamins in human milk: Methodological approaches. Crit. Rev. Food Sci. Nutr. 2016, 56, 494–511. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J. The developmental origins of chronic adult disease. Acta Paediatr. 2004, 93, 26–33. [Google Scholar] [CrossRef]
- Jensen, R.G. Handbook of Milk Composition; Academic Press Inc.: San Deigo, CA, USA, 1995. [Google Scholar]
- Kamao, M.; Tsugawa, N.; Suhara, Y.; Wada, A.; Mori, T.; Murata, K.; Nishino, R.; Ukita, T.; Uenishi, K.; Tanaka, K.; et al. Quantification of fat-soluble vitamins in human breast milk by liquid chromatography-tandem mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2007, 859, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Bolisetty, S.; Gupta, J.M.; Graham, G.G.; Salonikas, C.; Naidoo, D. Vitamin k in preterm breastmilk with maternal supplementation. Acta Paediatr. 1998, 87, 960–962. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.S.; Dimenstein, R.; Ribeiro, K.D. Vitamin e concentration in human milk and associated factors: A literature review. J. Pediatr. 2014, 90, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Jung, B.M.; Lee, B.N.; Kim, Y.J.; Jung, J.A.; Chang, N. Retinol, alpha-tocopherol, and selected minerals in breast milk of lactating women with full-term infants in south korea. Nutr. Res. Pract. 2017, 11, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Furukawa, M.; Asoh, M.; Kanno, T.; Kojima, T.; Yonekubo, A. Fat-soluble and water-soluble vitamin contents of breast milk from japanese women. J. Nutr. Sci. Vitaminol. 2005, 51, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Picciano, M.F. Clinical Nutrition of the Essential Trace Elements and Minerals; Springer Science and Business Media: New York, NY, USA, 2000. [Google Scholar]
- Dawodu, A.; Tsang, R.C. Maternal vitamin d status: Effect on milk vitamin d content and vitamin d status of breastfeeding infants. Adv. Nutr. 2012, 3, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.H. Multiple micronutrients in pregnancy and lactation: An overview. Am. J. Clin. Nutr. 2005, 81, 1206S–1212S. [Google Scholar] [PubMed]
- Hampel, D.; Shahab-Ferdows, S.; Adair, L.S.; Bentley, M.E.; Flax, V.L.; Jamieson, D.J.; Ellington, S.R.; Tegha, G.; Chasela, C.S.; Kamwendo, D.; et al. Thiamin and riboflavin in human milk: Effects of lipid-based nutrient supplementation and stage of lactation on vitamer secretion and contributions to total vitamin content. PLoS ONE 2016, 11, e0149479. [Google Scholar] [CrossRef] [PubMed]
- Treneva, M.; Munblit, D.; Pampura, A. Moscow infants: Atopic dermatitis, skin reactions to the dietary intake at 1-year of age followed to 2-years of age and sensitization at the age of 1 year. Allergy 2015, 70, 306. [Google Scholar]
- Page, R.; Robichaud, A.; Arbuckle, T.E.; Fraser, W.D.; MacFarlane, A.J. Total folate and unmetabolized folic acid in the breast milk of a cross-section of canadian women. Am. J. Clin. Nutr. 2017, 105, 1101–1109. [Google Scholar] [CrossRef] [PubMed]
- Houghton, L.A.; Yang, J.; O’Connor, D.L. Unmetabolized folic acid and total folate concentrations in breast milk are unaffected by low-dose folate supplements. Am. J. Clin. Nutr. 2009, 89, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Hampel, D.; Shahab-Ferdows, S.; Islam, M.M.; Peerson, J.M.; Allen, L.H. Vitamin concentrations in human milk vary with time within feed, circadian rhythm, and single-dose supplementation. J. Nutr. 2017, 147, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.M.; Chantry, C.J.; Young, S.L.; Achando, B.S.; Allen, L.H.; Arnold, B.F.; Colford, J.M., Jr.; Dentz, H.N.; Hampel, D.; Kiprotich, M.C.; et al. Vitamin b-12 concentrations in breast milk are low and are not associated with reported household hunger, recent animal-source food, or vitamin b-12 intake in women in rural kenya. J. Nutr. 2016, 146, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Deegan, K.L.; Jones, K.M.; Zuleta, C.; Ramirez-Zea, M.; Lildballe, D.L.; Nexo, E.; Allen, L.H. Breast milk vitamin b-12 concentrations in guatemalan women are correlated with maternal but not infant vitamin b-12 status at 12 months postpartum. J. Nutr. 2012, 142, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Maas, C.; Franz, A.R.; Shunova, A.; Mathes, M.; Bleeker, C.; Poets, C.F.; Schleicher, E.; Bernhard, W. Choline and polyunsaturated fatty acids in preterm infants’ maternal milk. Eur. J. Nutr. 2017, 56, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Gaxiola-Robles, R.; Labrada-Martagon, V.; Celis de la Rosa Ade, J.; Acosta-Vargas, B.; Mendez-Rodriguez, L.C.; Zenteno-Savin, T. Interaction between mercury (hg), arsenic (as) and selenium (se) affects the activity of glutathione s-transferase in breast milk; possible relationship with fish and sellfish intake. Nutr. Hosp. 2014, 30, 436–446. [Google Scholar] [PubMed]
- Shearer, T.R.; Hadjimarkos, D.M. Geographic distribution of selenium in human milk. Arch. Environ. Health 1975, 30, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Park, J.H.; Kim, E.A.; Lee-Kim, Y.C. Longitudinal study on trace mineral compositions (selenium, zinc, copper, manganese) in korean human preterm milk. J. Korean Med. Sci. 2012, 27, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Djurovic, D.; Milisavljevic, B.; Mugosa, B.; Lugonja, N.; Miletic, S.; Spasic, S.; Vrvic, M. Zinc concentrations in human milk and infant serum during the first six months of lactation. J. Trace Elem. Med. Biol. 2017, 41, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Chen, T.; Lu, W.; Wu, S.; Zhu, J. Breast milk macro- and micronutrient composition in lactating mothers from suburban and urban shanghai. J. Paediatr. Child Health 2010, 46, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, A.; O’Leary, P.; James, I.; Skeaff, S.; Sherriff, J. Assessment of breast milk iodine concentrations in lactating women in western australia. Nutrients 2016, 8, 699. [Google Scholar] [CrossRef] [PubMed]
- Henjum, S.; Kjellevold, M.; Ulak, M.; Chandyo, R.K.; Shrestha, P.S.; Froyland, L.; Strydom, E.E.; Dhansay, M.A.; Strand, T.A. Iodine concentration in breastmilk and urine among lactating women of bhaktapur, nepal. Nutrients 2016, 8, 255. [Google Scholar] [CrossRef] [PubMed]
- Osei, J.; Andersson, M.; Reijden, O.V.; Dold, S.; Smuts, C.M.; Baumgartner, J. Breast-milk iodine concentrations, iodine status, and thyroid function of breastfed infants aged 2–4 months and their mothers residing in a south african township. J. Clin. Res. Pediatr. Endocrinol. 2016, 8, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Domellof, M.; Braegger, C.; Campoy, C.; Colomb, V.; Decsi, T.; Fewtrell, M.; Hojsak, I.; Mihatsch, W.; Molgaard, C.; Shamir, R.; et al. Iron requirements of infants and toddlers. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Haskell, M.J.; Brown, K.H. Maternal vitamin a nutriture and the vitamin a content of human milk. J. Mammary Gland Biol. Neoplasia 1999, 4, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Turfkruyer, M.; Rekima, A.; Macchiaverni, P.; Le Bourhis, L.; Muncan, V.; van den Brink, G.R.; Tulic, M.K.; Verhasselt, V. Oral tolerance is inefficient in neonatal mice due to a physiological vitamin a deficiency. Mucosal Immunol. 2016, 9, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Nurmatov, U.; Devereux, G.; Sheikh, A. Nutrients and foods for the primary prevention of asthma and allergy: Systematic review and meta-analysis. J. Allergy Clin. Immunol. 2011, 127, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Aage, S.; Kiraly, N.; Da Costa, K.; Byberg, S.; Bjerregaard-Andersen, M.; Fisker, A.B.; Aaby, P.; Benn, C.S. Neonatal vitamin a supplementation associated with increased atopy in girls. Allergy 2015, 70, 985–994. [Google Scholar] [CrossRef] [PubMed]
- Duggan, C.; Srinivasan, K.; Thomas, T.; Samuel, T.; Rajendran, R.; Muthayya, S.; Finkelstein, J.L.; Lukose, A.; Fawzi, W.; Allen, L.H.; et al. Vitamin b-12 supplementation during pregnancy and early lactation increases maternal, breast milk, and infant measures of vitamin b-12 status. J. Nutr. 2014, 144, 758–764. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.H. B vitamins in breast milk: Relative importance of maternal status and intake, and effects on infant status and function. Adv. Nutr. 2012, 3, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Sasaki, S.; Tanaka, K.; Hirota, Y. Maternal b vitamin intake during pregnancy and wheeze and eczemea in japanese infants aged 16–24 months: The osaka maternal and child health study. Pediatr. Allergy Immunol. 2011, 22, 67–74. [Google Scholar]
- Ala-Houhala, M.; Koskinen, T.; Parviainen, M.T.; Visakorpi, J.K. 25-hydroxyvitamin d and vitamin d in human milk: Effects of supplementation and season. Am. J. Clin. Nutr. 1988, 48, 1057–1060. [Google Scholar] [PubMed]
- Greer, F.R.; Hollis, B.W.; Cripps, D.J.; Tsang, R.C. Effects of maternal ultraviolet b irradiation on vitamin d content of human milk. J. Pediatr. 1984, 105, 431–433. [Google Scholar] [CrossRef]
- Specker, B.L.; Tsang, R.C.; Hollis, B.W. Effect of race and diet on human-milk vitamin d and 25-hydroxyvitamin d. Am. J. Dis. Child. 1985, 139, 1134–1137. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, M.T.; Blumberg, D.L.; Hludzinski, J.; Kay, S. Nutritional rickets in suburbia. J. Am. Coll. Nutr. 1998, 17, 637–641. [Google Scholar] [CrossRef] [PubMed]
- Thiele, D.K.; Senti, J.L.; Anderson, C.M. Maternal vitamin d supplementation to meet the needs of the breastfed infant: A systematic review. J. Hum. Lact. 2013, 29, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.L.; Greer, F.R. Prevention of rickets and vitamin d deficiency in infants, children, and adolescents. Pediatrics 2008, 122, 1142–1152. [Google Scholar] [CrossRef] [PubMed]
- Weisse, K.; Winkler, S.; Hirche, F.; Herberth, G.; Hinz, D.; Bauer, M.; Roder, S.; Rolle-Kampczyk, U.; von Bergen, M.; Olek, S.; et al. Maternal and newborn vitamin d status and its impact on food allergy development in the german lina cohort study. Allergy 2013, 68, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Gale, C.R.; Robinson, S.M.; Harvey, N.C.; Javaid, M.K.; Jiang, B.; Martyn, C.N.; Godfrey, K.M.; Cooper, C.; Princess Anne Hospital Study Group. Maternal vitamin d status during pregnancy and child outcomes. Eur. J. Clin. Nutr. 2008, 62, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Erkkola, M.; Kaila, M.; Nwaru, B.I.; Kronberg-Kippila, C.; Ahonen, S.; Nevalainen, J.; Veijola, R.; Pekkanen, J.; Ilonen, J.; Simell, O.; et al. Maternal vitamin d intake during pregnancy is inversely associated with asthma and allergic rhinitis in 5-year-old children. Clin. Exp. Allergy 2009, 39, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.J.; Ponsonby, A.; Gurrin, L.C.; Wake, M.; Vuillermin, P.; Martin, P.; Matheson, M.L.A.; Robinson, M.; Tey, D.; Osborne, N.J.; et al. Vitamin d insufficiency is associated with challenge-proven food allergy in infants. J. Allergy Clin. Immunol. 2013, 131, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Hon, K.L.; Kong, A.P.; Pong, H.N.; Wong, G.W.; Leung, T.F. Vitamin d deficiency is associated with diagnosis and severity of childhood atopic dermatitis. Pediatr. Allergy Immunol. 2014, 25, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Samochocki, Z.; Bogaczewicz, J.; Jeziorkowska, R.; Sysa-Jedrzejowska, A.; Glinska, O.; Karczmarewicz, E.; McCauliffe, D.P.; Wozniacka, A. Vitamin d effects in atopic dermatitis. J. Am. Acad. Dermatol. 2013, 69, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Litonjua, A.A.; Carey, V.J.; Laranjo, N.; Harshfield, B.J.; McElrath, T.F.; O’Connor, G.T.; Sandel, M.; Iverson, R.E., Jr.; Lee-Paritz, A.; Strunk, R.C.; et al. Effect of prenatal supplementation with vitamin d on asthma or recurrent wheezing in offspring by age 3 years: The vdaart randomized clinical trial. J. Am. Med. Assoc. 2016, 315, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Goldring, S.T.; Griffiths, C.J.; Martineau, A.R.; Robinson, S.; Yu, C.; Poulton, S.; Kirkby, J.C.; Stocks, J.; Hooper, R.; Shaheen, S.O.; et al. Prenatal vitamin d supplementation and child respiratory health: A randomised controlled trial. PLoS ONE 2013, 8, e66627. [Google Scholar] [CrossRef] [PubMed]
- Chawes, B.L.; Bonnelykke, K.; Stokholm, J.; Vissing, N.H.; Bjarnadottir, E.; Schoos, A.M.; Wolsk, H.M.; Pedersen, T.M.; Vinding, R.K.; Thorsteinsdottir, S.; et al. Effect of vitamin d3 supplementation during pregnancy on risk of persistent wheeze in the offspring: A randomized clinical trial. J. Am. Med. Assoc. 2016, 315, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Tuokkola, J.; Luukkainen, P.; Kaila, M.; Takkinen, H.M.; Niinisto, S.; Veijola, R.; Virta, L.J.; Knip, M.; Simell, O.; Ilonen, J.; et al. Maternal dietary folate, folic acid and vitamin d intakes during pregnancy and lactation and the risk of cows’ milk allergy in the offspring. Br. J. Nutr. 2016, 116, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Cuello-Garcia, C.A.; Fiocchi, A.; Pawankar, R.; Yepes-Nuñez, J.J.; Morgano, G.P.; Zhang, Y.; Ahn, K.; Al-Hammadi, S.; Agarwal, A.; Gandhi, S.; et al. World allergy organization-mcmaster university guidelines for allergic disease prevention (glad-p): Vitamin d. World Allergy Organ. J. 2016, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Hernell, O.; Fewtrell, M.S.; Georgieff, M.K.; Krebs, N.F.; Lonnerdal, B. Summary of current recommendations on iron provision and monitoring of iron status for breastfed and formula-fed infants in resource-rich and resource-constrained countries. J. Pediatr. 2015, 167, S40–S47. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K.; Wewerka, S.W.; Nelson, C.A.; Deregnier, R.A. Iron status at 9 months of infants with low iron stores at birth. J. Pediatr. 2002, 141, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Fewtrell, M.; Bronsky, J.; Campoy, C.; Domellof, M.; Embleton, N.; Fidler Mis, N.; Hojsak, I.; Hulst, J.M.; Indrio, F.; Lapillonne, A.; et al. Complementary feeding: A position paper by the european society for paediatric gastroenterology, hepatology, and nutrition (espghan) committee on nutrition. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Krebs, N.F.; Sherlock, L.G.; Westcott, J.; Culbertson, D.; Hambidge, K.M.; Feazel, L.M.; Robertson, C.E.; Frank, D.N. Effects of different complementary feeding regimens on iron status and enteric microbiota in breastfed infants. J. Pediatr. 2013, 163, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Friel, J.K.; Aziz, K.; Andrews, W.L.; Harding, S.V.; Courage, M.L.; Adams, R.J. A double-masked, randomized control trial of iron supplementation in early infancy in healthy term breast-fed infants. J. Pediatr. 2003, 143, 582–586. [Google Scholar] [CrossRef]
- Baker, R.D.; Greer, F.R. Diagnosis and prevention of iron deficiency and iron-deficiency anemia in infants and young children (0–3 years of age). Pediatrics 2010, 126, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- WHO. The Global Prevalence of Anaemia in 2011; WHO: Geneva, Switzerland, 2015. [Google Scholar]
- Toyran, M.; Kaymak, M.; Vezir, E.; Harmanci, K.; Kaya, A.; Ginis, T.; Kose, G.; Kocabas, C.N. Trace element levels in children with atopic dermatitis. J. Investig. Allergol. Clin. Immunol. 2012, 22, 341–344. [Google Scholar] [PubMed]
- Gibson, R.S. A historical review of progress in the assessment of dietary zinc intake as an indicator of population zinc status. Adv. Nutr. 2012, 3, 772–782. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.H.; Rivera, J.A.; Bhutta, Z.; Gibson, R.S.; King, J.C.; Lonnerdal, B.; Ruel, M.T.; Sandtrom, B.; Wasantwisut, E.; Hotz, C. International zinc nutrition consultative group (izincg) technical document #1. Assessment of the risk of zinc deficiency in populations and options for its control. Food Nutr. Bull. 2004, 25, S99–S203. [Google Scholar] [PubMed]
- Krebs, N.F.; Miller, L.V.; Michael Hambidge, K. Zinc deficiency in infants and children: A review of its complex and synergistic interactions. Paediatr. Int. Child. Health 2014, 34, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Kent, J.C.; Mitoulas, L.R.; Cregan, M.D.; Ramsay, D.T.; Doherty, D.A.; Hartmann, P.E. Volume and frequency of breastfeedings and fat content of breast milk throughout the day. Pediatrics 2006, 117, e387–e395. [Google Scholar] [CrossRef] [PubMed]
- Moco, S.; Collino, S.; Rezzi, S.; Martin, F.P. Metabolomics perspectives in pediatric research. Pediatr. Res. 2013, 73, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Sundekilde, U.K.; Downey, E.; O’Mahony, J.A.; O’Shea, C.A.; Ryan, C.A.; Kelly, A.L.; Bertram, H.C. The effect of gestational and lactational age on the human milk metabolome. Nutrients 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Andreas, N.J.; Hyde, M.J.; Gomez-Romero, M.; Lopez-Gonzalvez, M.A.; Villasenor, A.; Wijeyesekera, A.; Barbas, C.; Modi, N.; Holmes, E.; Garcia-Perez, I. Multiplatform characterization of dynamic changes in breast milk during lactation. Electrophoresis 2015. [Google Scholar] [CrossRef] [PubMed]
- Villasenor, A.; Garcia-Perez, I.; Garcia, A.; Posma, J.M.; Fernandez-Lopez, M.; Nicholas, A.J.; Modi, N.; Holmes, E.; Barbas, C. Breast milk metabolome characterization in a single-phase extraction, multiplatform analytical approach. Anal. Chem. 2014, 86, 8245–8252. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Domellof, M.; Zivkovic, A.M.; Larsson, G.; Ohman, A.; Nording, M.L. Nmr-based metabolite profiling of human milk: A pilot study of methods for investigating compositional changes during lactation. Biochem. Biophys. Res. Commun. 2016, 469, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Palmas, F.; Fattuoni, C.; Noto, A.; Barberini, L.; Dessi, A.; Fanos, V. The choice of amniotic fluid in metabolomics for the monitoring of fetus health. Expert Rev. Mol. Diagn. 2016, 16, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Jantscher-Krenn, E.; Zherebtsov, M.; Nissan, C.; Goth, K.; Guner, Y.S.; Naidu, N.; Choudhury, B.; Grishin, A.V.; Ford, H.R.; Bode, L. The human milk oligosaccharide disialyllacto-n-tetraose prevents necrotising enterocolitis in neonatal rats. Gut 2012, 61, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Autran, C.A.; Kellman, B.P.; Kim, J.H.; Asztalos, E.; Blood, A.B.; Spence, E.C.; Patel, A.L.; Hou, J.; Lewis, N.E.; Bode, L. Human milk oligosaccharide composition predicts risk of necrotising enterocolitis in preterm infants. Gut 2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, M.; Wu, S.; Lebrilla, C.B.; Chapkin, R.S.; Ivanov, I.; Donovan, S.M. Fecal microbiota composition of breast-fed infants is correlated with human milk oligosaccharides consumed. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Mosconi, E.; Rekima, A.; Seitz-Polski, B.; Kanda, A.; Fleury, S.; Tissandie, E.; Monteiro, R.; Dombrowicz, D.D.; Julia, V.; Glaichenhaus, N.; et al. Breast milk immune complexes are potent inducers of oral tolerance in neonates and prevent asthma development. Mucosal Immunol. 2010, 3, 461–474. [Google Scholar] [CrossRef] [PubMed]
- Verhasselt, V. Neonatal tolerance under breastfeeding influence: The presence of allergen and transforming growth factor-beta in breast milk protects the progeny from allergic asthma. J. Pediatr. 2010, 156, S16–S20. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Kozyrskyj, A.L. Gut microbial metabolism defines host metabolism: An emerging perspective in obesity and allergic inflammation. Obes. Rev. 2017, 18, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Julia, V.; Macia, L.; Dombrowicz, D. The impact of diet on asthma and allergic diseases. Nat. Rev. Immunol. 2015, 15, 308–322. [Google Scholar] [CrossRef] [PubMed]
- Corthesy, B. Multi-faceted functions of secretory iga at mucosal surfaces. Front. Immunol. 2013, 4, 185. [Google Scholar] [CrossRef] [PubMed]
- Hennet, T.; Borsig, L. Breastfed at tiffany’s. Trends Biochem. Sci. 2016, 41, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Brandtzaeg, P. The mucosal immune system and its integration with the mammary glands. J. Pediatr. 2010, 156, S8–S15. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, K.; Hida, M.; Kohgo, T.; Fukunaga, Y. Changes in salivary and fecal secretory iga in infants under different feeding regimens. Pediatr. Int. 2009, 51, 342–345. [Google Scholar] [CrossRef] [PubMed]
- Urwin, H.J.; Miles, E.A.; Noakes, P.S.; Kremmyda, L.S.; Vlachava, M.; Diaper, N.D.; Godfrey, K.M.; Calder, P.C.; Vulevic, J.; Yaqoob, P. Effect of salmon consumption during pregnancy on maternal and infant faecal microbiota, secretory iga and calprotectin. Br. J. Nutr. 2014, 111, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Kukkonen, K.; Kuitunen, M.; Haahtela, T.; Korpela, R.; Poussa, T.; Savilahti, E. High intestinal iga associates with reduced risk of ige-associated allergic diseases. Pediatr. Allergy Immunol. 2010, 21, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Koutras, A.K.; Vigorita, V.J. Fecal secretory immunoglobulin a in breast milk versus formula feeding in early infancy. J. Pediatr. Gastroenterol. Nutr. 1989, 9, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Munblit, D.; Sheth, S.; Abrol, P.; Treneva, M.; Peroni, D.G.; Chow, L.Y.; Boner, A.L.; Pampura, A.; Warner, J.O.; Boyle, R.J. Exposures influencing total iga level in colostrum. J. Dev. Orig. Health Dis. 2016, 7, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Breakey, A.A.; Hinde, K.; Valeggia, C.R.; Sinofsky, A.; Ellison, P.T. Illness in breastfeeding infants relates to concentration of lactoferrin and secretory immunoglobulin a in mother’s milk. Evol. Med. Public Health 2015, 2015, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Bridgman, S.L.; Konya, T.; Azad, M.B.; Sears, M.R.; Becker, A.B.; Turvey, S.E.; Mandhane, P.J.; Subbarao, P.; Investigators, C.S.; Scott, J.A.; et al. Infant gut immunity: A preliminary study of iga associations with breastfeeding. J. Dev. Orig. Health Dis. 2016, 7, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Bridgman, S.L.; Konya, T.; Azad, M.B.; Guttman, D.S.; Sears, M.R.; Becker, A.B.; Turvey, S.E.; Mandhane, P.J.; Subbarao, P.; Investigators, C.S.; et al. High fecal iga is associated with reduced clostridium difficile colonization in infants. Microbes Infect. Inst. Pasteur 2016, 18, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Adlerberth, I.; Wold, A.E. Establishment of the gut microbiota in western infants. Acta Paediatr. 2009, 98, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Penders, J.; Thijs, C.; van den Brandt, P.A.; Kummeling, I.; Snijders, B.; Stelma, F.; Adams, H.; van Ree, R.; Stobberingh, E.E. Gut microbiota composition and development of atopic manifestations in infancy: The koala birth cohort study. Gut 2007, 56, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Mastromarino, P.; Capobianco, D.; Campagna, G.; Laforgia, N.; Drimaco, P.; Dileone, A.; Baldassarre, M.E. Correlation between lactoferrin and beneficial microbiota in breast milk and infant’s feces. Biometals 2014, 27, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Cacho, N.T.; Lawrence, R.M. Innate immunity and breast milk. Front. Immunol. 2017, 8, 584. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.A.; Sjogren, Y.M.; Persson, J.O.; Nilsson, C.; Sverremark-Ekstrom, E. Early colonization with a group of lactobacilli decreases the risk for allergy at five years of age despite allergic heredity. PLoS ONE 2011, 6, e23031. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Lai, C.T.; Hartmann, P.; Oddy, W.H.; Kusel, M.M.; Sly, P.D.; Holt, P.G. Anti-infective proteins in breast milk and asthma-associated phenotypes during early childhood. Pediatr. Allergy Immunol. 2014, 25, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Yates, C.M.; Calder, P.C.; Ed Rainger, G. Pharmacology and therapeutics of omega-3 polyunsaturated fatty acids in chronic inflammatory disease. Pharmacol. Ther. 2014, 141, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Miyata, J.; Arita, M. Role of omega-3 fatty acids and their metabolites in asthma and allergic diseases. Allergol. Int. 2015, 64, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Koltsida, O.; Karamnov, S.; Pyrillou, K.; Vickery, T.; Chairakaki, A.D.; Tamvakopoulos, C.; Sideras, P.; Serhan, C.N.; Andreakos, E. Toll-like receptor 7 stimulates production of specialized pro-resolving lipid mediators and promotes resolution of airway inflammation. EMBO Mol. Med. 2013, 5, 762–775. [Google Scholar] [CrossRef] [PubMed]
- Thorburn, A.N.; McKenzie, C.I.; Shen, S.; Stanley, D.; Macia, L.; Mason, L.J.; Roberts, L.K.; Wong, C.H.; Shim, R.; Robert, R.; et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat. Commun. 2015, 6, 7320. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, H.S.; Holtug, K.; Ynggard, C.; Mortensen, P.B. Faecal concentrations and production rates of short chain fatty acids in normal neonates. Acta Paediatr. Scand. 1988, 77, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Bridgman, S.L.; Azad, M.B.; Field, C.J.; Haqq, A.M.; Becker, A.B.; Mandhane, P.J.; Subbarao, P.; Turvey, S.E.; Sears, M.R.; Scott, J.A.; et al. Fecal short-chain fatty acid variations by breastfeeding status in infants at 4 months: Differences in relative versus absolute concentrations. Front. Nutr. 2017, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Vuillermin, P.J.; Goverse, G.; Vinuesa, C.G.; Mebius, R.E.; Macia, L.; Mackay, C.R. Dietary fiber and bacterial scfa enhance oral tolerance and protect against food allergy through diverse cellular pathways. Cell Rep. 2016, 15, 2809–2824. [Google Scholar] [CrossRef] [PubMed]
- Stefka, A.T.; Feehley, T.; Tripathi, P.; Qiu, J.; McCoy, K.; Mazmanian, S.K.; Tjota, M.Y.; Seo, G.Y.; Cao, S.; Theriault, B.R.; et al. Commensal bacteria protect against food allergen sensitization. Proc. Natl. Acad. Sci. USA 2014, 111, 13145–13150. [Google Scholar] [CrossRef] [PubMed]
- Albright, C.D.; Tsai, A.Y.; Friedrich, C.B.; Mar, M.H.; Zeisel, S.H. Choline availability alters embryonic development of the hippocampus and septum in the rat. Brain Res. Dev. Brain Res. 1999, 113, 13–20. [Google Scholar] [CrossRef]
- Ilcol, Y.O.; Ozbek, R.; Hamurtekin, E.; Ulus, I.H. Choline status in newborns, infants, children, breast-feeding women, breast-fed infants and human breast milk. J. Nutr. Biochem. 2005, 16, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Ozarda, Y.; Cansev, M.; Ulus, I.H. Breast milk choline contents are associated with inflammatory status of breastfeeding women. J. Hum. Lact. 2014, 30, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Marnell, L.; Mold, C.; Du Clos, T.W. C-reactive protein: Ligands, receptors and role in inflammation. Clin. Immunol. 2005, 117, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Kushner, I.; Rzewnicki, D.; Samols, D. What does minor elevation of c-reactive protein signify? Am. J. Med. 2006, 119, 166. [Google Scholar] [CrossRef] [PubMed]
- Detopoulou, P.; Panagiotakos, D.B.; Antonopoulou, S.; Pitsavos, C.; Stefanadis, C. Dietary choline and betaine intakes in relation to concentrations of inflammatory markers in healthy adults: The attica study. Am. J. Clin. Nutr. 2008, 87, 424–430. [Google Scholar] [PubMed]
- Bager, P.; Wohlfahrt, J.; Westergaard, T. Caesarean delivery and risk of atopy and allergic disease: Meta-analyses. Clin. Exp. Allergy 2008, 38, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Cederlund, A.; Kai-Larsen, Y.; Printz, G.; Yoshio, H.; Alvelius, G.; Lagercrantz, H.; Stromberg, R.; Jornvall, H.; Gudmundsson, G.H.; Agerberth, B. Lactose in human breast milk an inducer of innate immunity with implications for a role in intestinal homeostasis. PLoS ONE 2013, 8, e53876. [Google Scholar] [CrossRef] [PubMed]
- Marincola, F.C.; Noto, A.; Caboni, P.; Reali, A.; Barberini, L.; Lussu, M.; Murgia, F.; Santoru, M.L.; Atzori, L.; Fanos, V. A metabolomic study of preterm human and formula milk by high resolution nmr and gc/ms analysis: Preliminary results. J. Matern. Fetal Neonatal Med. 2012, 25, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Skovbjerg, H.; Sjostrom, H.; Noren, O. Purification and characterisation of amphiphilic lactase/phlorizin hydrolase from human small intestine. Eur. J. Biochem. 1981, 114, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Hove, H.; Norgaard, H.; Mortensen, P.B. Lactic acid bacteria and the human gastrointestinal tract. Eur. J. Clin. Nutr. 1999, 53, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.M.; Sitarik, A.R.; Havstad, S.L.; Fujimura, K.E.; Wegienka, G.; Cassidy-Bushrow, A.E.; Kim, H.; Zoratti, E.M.; Lukacs, N.W.; Boushey, H.A.; et al. Joint effects of pregnancy, sociocultural, and environmental factors on early life gut microbiome structure and diversity. Sci. Rep. 2016, 6, 31775. [Google Scholar] [CrossRef] [PubMed]
- Madan, J.C.; Hoen, A.G.; Lundgren, S.N.; Farzan, S.F.; Cottingham, K.L.; Morrison, H.G.; Sogin, M.L.; Li, H.; Moore, J.H.; Karagas, M.R. Association of cesarean delivery and formula supplementation with the intestinal microbiome of 6-week-old infants. JAMA Pediatr. 2016, 170, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Konya, T.; Persaud, R.R.; Guttman, D.S.; Chari, R.S.; Field, C.J.; Sears, M.R.; Mandhane, P.J.; Turvey, S.E.; Subbarao, P.; et al. Impact of maternal intrapartum antibiotics, method of birth and breastfeeding on gut microbiota during the first year of life: A prospective cohort study. Int. J. Obstet. Gynaecol. 2016, 123, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Kramer, M.S. Does breast feeding help protect against atopic disease? Biology, methodology, and a golden jubilee of controversy. J. Pediatr. 1988, 112, 181–190. [Google Scholar] [CrossRef]


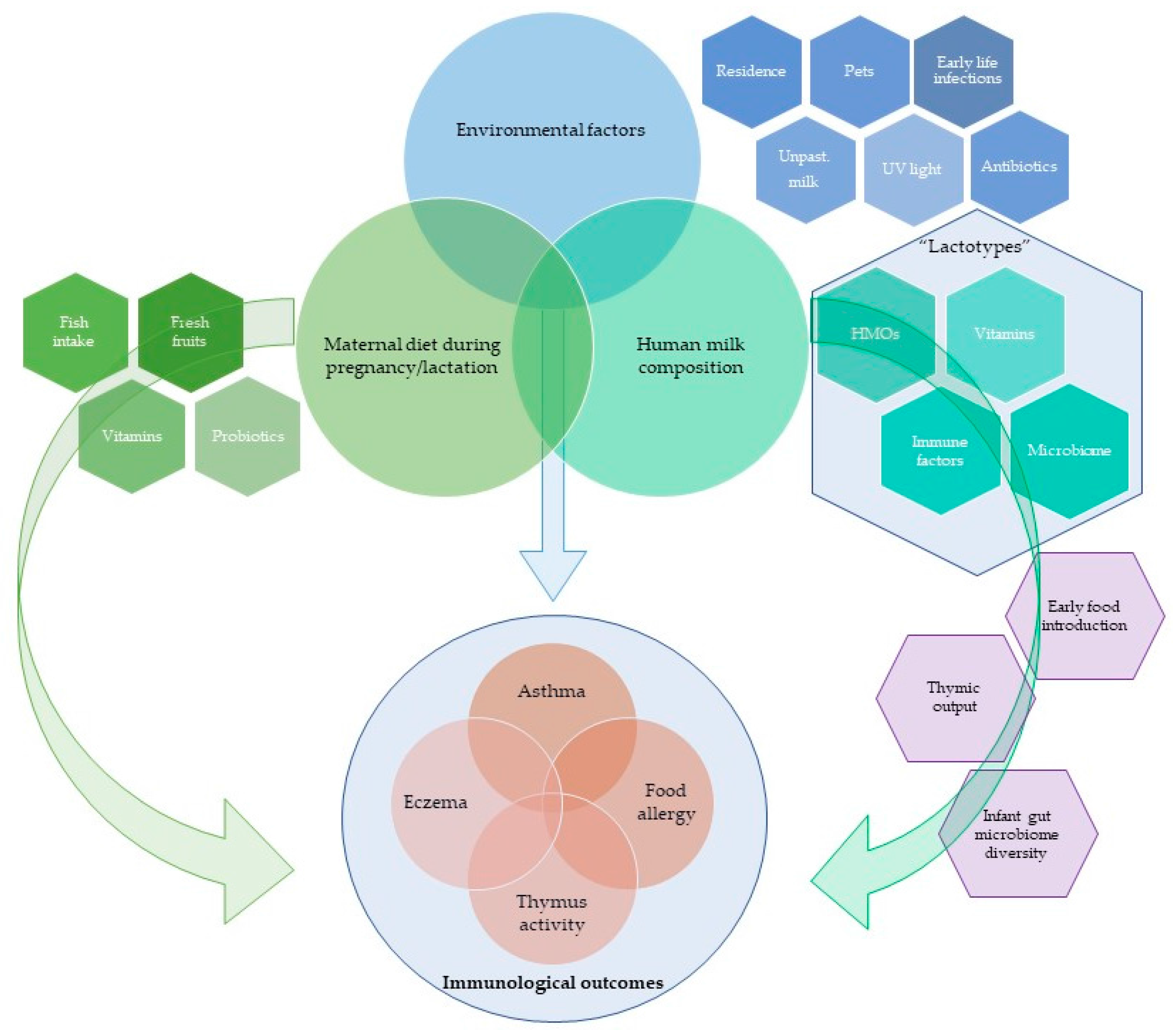{kind=link}
| Study | Intervention | Time of HM Collection Postpartum | HM Composition Changes |
|---|---|---|---|
| Fish Oil and Fresh Fish | |||
| Hawkes 2001 [119] | Fish oil supplementation | 5 weeks | no significant influence on TGF-β1 and TGF-β2 |
| Dunstan 2004 [120] | Fish oil supplementation | 3 days | no significant influence on IgA and sCD14 levels |
| Urwin 2012 [121] | Farmed salmon supplementation | 1, 5 and 28 days | no significant influence on TGF-β1, TGF-β2 and sCD14 |
| Probiotics | |||
| Bottcher 2008 [122] | Probiotic supplementation (L. reuteri) | 3 days and 1 month | ↓ TGF-β2 and ↑ IL-10 (borderline significance) in 3 day samples no difference in IgA, SIgA, TGF-β1, TNF, sCD14 in 1 month samples |
| Prescott 2008 [123] | Probiotic supplementation (L. rhamnosus or B. lactis) | 7 days | ↑ TGF-β1 in HM from B. lactis group no significant influence on IL6, IL10, IL13, IFN-γ, TNF-α, sCD14, total IgA |
| Boyle 2011 [124] | Probiotic supplementation (L. rhamnosus) | 7 and 28 days | ↓ sCD14 and IgA levels in HM from L. rhamnosus GG group no significant influence of on TGF-β1 |
| Hoppu 2012 [125] | Diet and Probiotic supplementation (L. rhamnosus and B. lactis) | colostrum (after birth) and 1 month | ↑ IL-2, IL-4, IL10 TNF-α and total n-3 fatty acids in probiotic group no significant influence on IFN-γ and IL6 |
| Kuitunen 2012 [126] | Probiotic supplementation (A combination of 2 species of L. rhamnosus, B. breve and P. freudenreichii) | 0–3 days and 3 months | ↑ IL-10 and ↓ casein IgA antibodies in probiotics group |
| Savilahti 2015 [127] | Probiotic supplementation (A combination of 2 species of L. rhamnosus, B. breve and P. freudenreichii) | 0–3 days and 3 months | no significant influence on sCD14, HBD2 and HNP1–3 |
| Other Interventions | |||
| Linnamaa 2013 [128] | Blackcurrant seed oil | after delivery and 3 months | ↑ IFN-γ and ↓ IL-4 in blackcurrant seed oil group no significant influence on IL-5, IL-10, IL-12 and TNF levels |
| Nikniaz 2013 [129] | Synbiotic | 3 and 4 months | ↑ IgA and TGF-β2 in synbiotic group no significant influence on TGF-β1 |
| Study | Allergic Outcomes Assessed | Relationship between Human Milk Composition and Outcomes | |
|---|---|---|---|
| Human Milk Composition Factors | Outcome of Influence | ||
| Kalliomaki 1999 [130] | Eczema (up to 12 months) | ↑ TGF-β1 and TGF-β2 (colostrum) | higher post weaning-onset atopic disease |
| Jones 2002 [131] | Eczema (up to 6 months) | ↓ sCD14 (3 months HM) | higher eczema incidence at 6 months of age |
| Bottcher 2003 [132] | Allergic sensitisation (up to 2 years) Salivary IgA (up to 2 years) Eczema (up to 2 years) | IL-4, IL-5, IL-6, IL-8, IL-10, IL-13, IL-16, IFN-γ, TGF-β1, TGF-β2, RANTES, eotaxin or SIgA (colostrum and 1 month HM) | no significant influence on atopy and/or allergy |
| Oddy 2003 [133] | Asthma-like symptoms (up to 12 months) | ↑ TGF-β1 (2 weeks HM) TNF-α, sCD14 and IL10 (2 weeks HM) | lower risk of wheeze in infancy no significant association with infant wheeze |
| Savilahti 2005 [134] | Allergic sensitisation (up to 4 years) Eczema (up to 4 years) | ↓ IgA casein antibodies and sCD14 (colostrum) | higher incidence of atopy development |
| Snijders 2006 [135] | Eczema (up to 12 months) Allergic sensitisation (up to 2 years) Wheezing (up to 2 years) | TGF-β1, IL-10, IL-12 and sCD14 (1 month HM) | no significant influence on any of the atopic manifestations |
| Bottcher 2008 [122] | Allergic sensitisation (up to 2 years) Eczema (up to 2 years) | ↓ TGF-β2 (colostrum) | lower incidence of sensitisation during the first 2 years of life a trend of protective effect on eczema development |
| Kuitunen 2012 [126] | Allergic diseases (up to 5 years) Eczema (up to 5 years) Allergic sensitisation (up to 2 years) | ↑ TGF-β2 (3 month HM) IL-10 and TGF-β2 (3 month HM) | higher risk of allergic disease and eczema at 2 years of age no significant association with allergic outcomes at 2 and 5 years of age |
| Soto-Ramirez 2012 [136] | Asthma-like symptoms (up to 12 months) | infants in the highest quartile of IL-5 and IL-13 (2 weeks HM) | higher risk of asthma-like symptoms development |
| Ismail 2013 [137] | Eczema (up to 12 months) Allergic sensitisation (up to 12 months) | TGF-β1, sCD14, total IgA (7 and 28 days HM) | no significant association with any of the atopic manifestations |
| Orivuori 2014 [138] | Eczema (up to 4 years) Asthma (up to 6 years) Allergic sensitisation (up to 6 years) | ↑ sIgA (2 months HM) TGF-β1 (2 months HM) sIgA (2 months HM) | lower eczema incidence up to the age of 2 years no significant association with the outcomes no significant association with atopy or asthma up to the age of 6 |
| Savilahti 2015 [127] | Allergic diseases (up to 5 years) | ↑ sCD14 (3 months HM) | higher incidence of allergic sensitisation and eczema |
| Jepsen 2016 [58] | Eczema (up to 3 years) Recurrent wheeze (up to 3 years) | ↑ IL-1β (1 month HM) CXCL10, TNF-α, CCL2, CCL4, CCL5, CCL17, CCL22, CCL26, TSLP, IL17, CXCL1, CXCL8, TGF-β1 (1 month HM) | lower eczema incidence up to the age of 3 years no significant association with eczema or wheeze |
| Munblit 2017 [139] | Eczema-like symptoms (up to 6 months) Wheeze (up to 6 months) Food allergy parental-reported (up to 6 months) | ↑ TGF-β2 (1 month HM) detectable IL-13 (colostrum) detectable IL-13 (1 month HM) HGF, TGF-β1, TGF-β3, IL-2, IL-4, IL-5, IL-10, IFN-γ, IL-12 (colostrum and 1 month HM) | higher risk of eczema lower risk of food allergy lower risk of eczema no significant association with eczema, wheeze or food allergy |
| Component Affected by Maternal Diet | Concentration | Component Unaffected by Maternal Diet | Concentration |
|---|---|---|---|
| Fat Soluble Vitamins | |||
| K | 0.12–0.98 ug/dL [241,242,243] | Tocopherol (vit E) | 207–366 ug/dL [244,245,246] |
| D | 0.008–0.62 ug/dL [242,246,247,248] | ||
| Retinol (Vit. A) * | 40–485 μg/L [242,245,249] | ||
| Water Soluble Vitamins | |||
| Thiamin (vit B-1) | 21.1–228 ug/L [249,250,251] | Folate | 53–133 ug/L [241,252,253] |
| Riboflavin (vit B-2) | 0.03–0.35 mg/L [249,251] | ||
| Niacin (vit B-3) | 68.7–260 ug/L [251,254] | ||
| Vit B-6 | 0.06–0.31 mg/L [241,249,251,255] | ||
| Cobalamin (vit B-12) | 85–970 ng/L [249,255,256] | ||
| Ascorbic acid (vit C) | 35–105 mg/L [241,246,249] | ||
| Pantothenic acid (vit B-5) | 2.0–2.5 mg/L [241,251] | ||
| Choline | 144–258 mg/L [241,257] | ||
| Minerals | |||
| Selenium | 3–60 ng/mL [241,249,258,259] | Zinc | 0.68–12 ug/mL [241,245,260,261,262] |
| Iodine | 9–250 ug/L [241,249,263,264,265] | Copper | 0.006–0.5 ug/mL [241,245,253] |
| Iron | 0.3–0.9 ug/mL [245,262,266] | ||
| Calcium | 259–300 mg/L [241,245,262] | ||
| Phosphorus | 130–170 mg/L [241,245,262] | ||
| Magnesium | 30.5–31.4 mg/L [241,245] | ||
| Sodium | 111–300 mg/L [241,245,262] | ||
| Potassium | 380–630 mg/L [241,245,262] | ||
| Chromium | 0.15–0.8 ng/mL [241,247,253] | ||
| Chloride | 453–690 mg/L [241,262] | ||
| Manganese | 0.33–125 ng/mL [241,245,253,262] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munblit, D.; Peroni, D.G.; Boix-Amorós, A.; Hsu, P.S.; Land, B.V.; Gay, M.C.L.; Kolotilina, A.; Skevaki, C.; Boyle, R.J.; Collado, M.C.; et al. Human Milk and Allergic Diseases: An Unsolved Puzzle. Nutrients 2017, 9, 894. https://doi.org/10.3390/nu9080894
Munblit D, Peroni DG, Boix-Amorós A, Hsu PS, Land BV, Gay MCL, Kolotilina A, Skevaki C, Boyle RJ, Collado MC, et al. Human Milk and Allergic Diseases: An Unsolved Puzzle. Nutrients. 2017; 9(8):894. https://doi.org/10.3390/nu9080894
Chicago/Turabian StyleMunblit, Daniel, Diego G. Peroni, Alba Boix-Amorós, Peter S. Hsu, Belinda Van’t Land, Melvin C. L. Gay, Anastasia Kolotilina, Chrysanthi Skevaki, Robert J. Boyle, Maria Carmen Collado, and et al. 2017. "Human Milk and Allergic Diseases: An Unsolved Puzzle" Nutrients 9, no. 8: 894. https://doi.org/10.3390/nu9080894
APA StyleMunblit, D., Peroni, D. G., Boix-Amorós, A., Hsu, P. S., Land, B. V., Gay, M. C. L., Kolotilina, A., Skevaki, C., Boyle, R. J., Collado, M. C., Garssen, J., Geddes, D. T., Nanan, R., Slupsky, C., Wegienka, G., Kozyrskyj, A. L., & Warner, J. O. (2017). Human Milk and Allergic Diseases: An Unsolved Puzzle. Nutrients, 9(8), 894. https://doi.org/10.3390/nu9080894