Yerba Maté (Ilex paraguariensis) Metabolic, Satiety, and Mood State Effects at Rest and during Prolonged Exercise


Abstract
:1. Introduction
2. Materials and Methods
2.1. Design and Participants
2.2. Experimental Procedures and Exercise Protocols
2.3. Baseline Assessments (Visit One)
2.4. Exercise Assessment and Supplementation (Visits Two and Three)
2.5. Appetite and Mood State
2.6. Data Processing, Analyses, and Statistics
3. Results
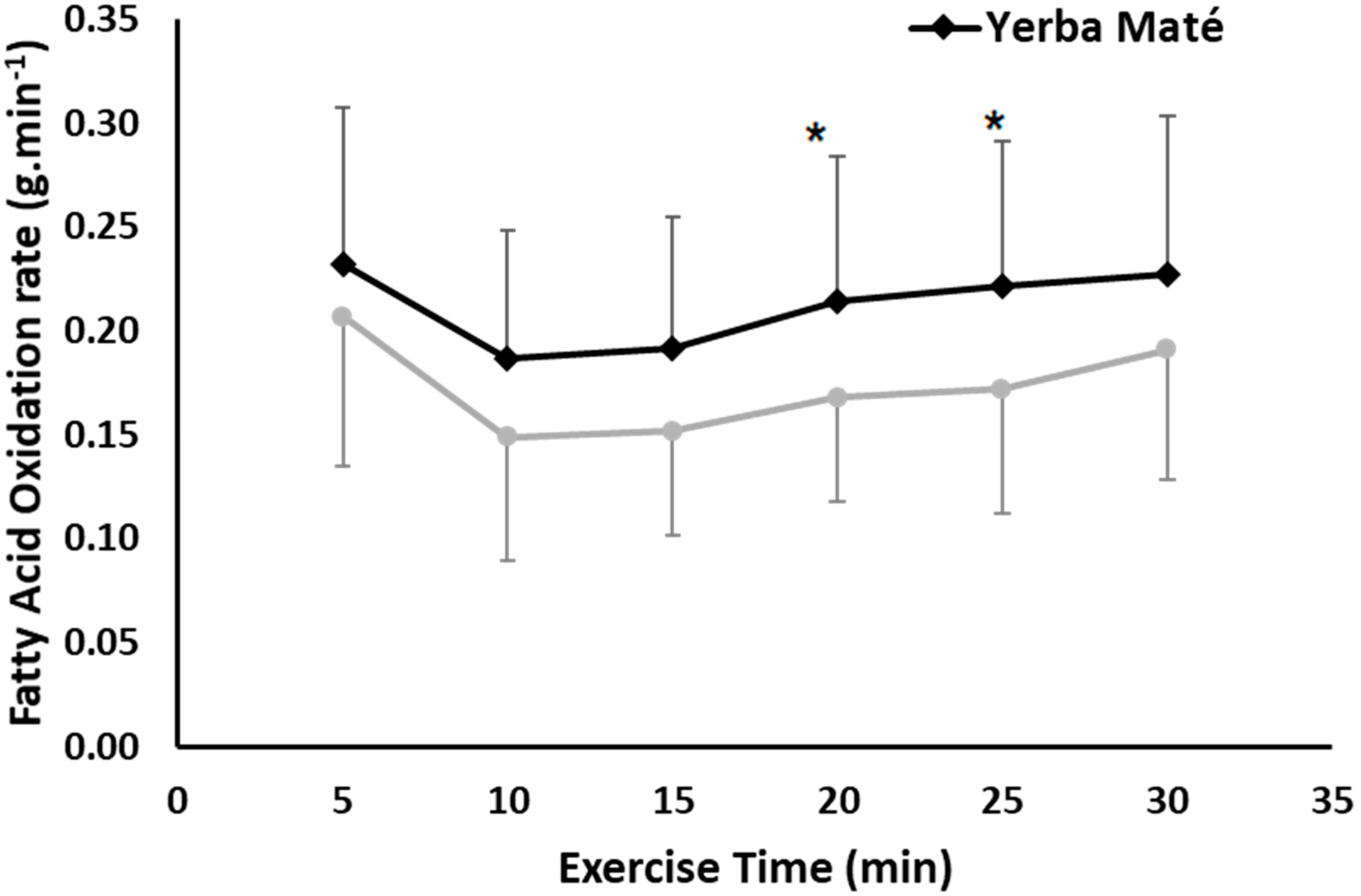
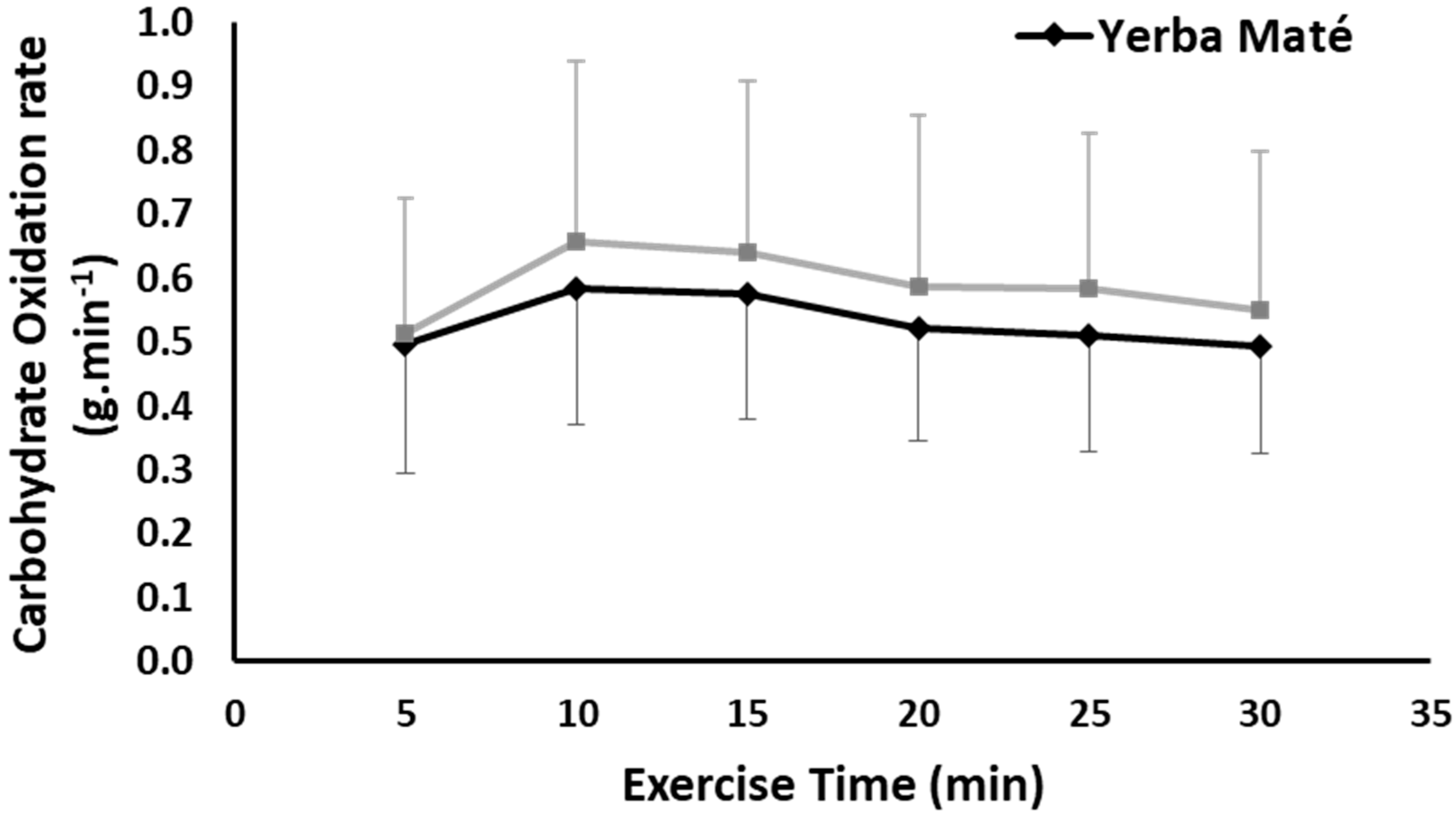
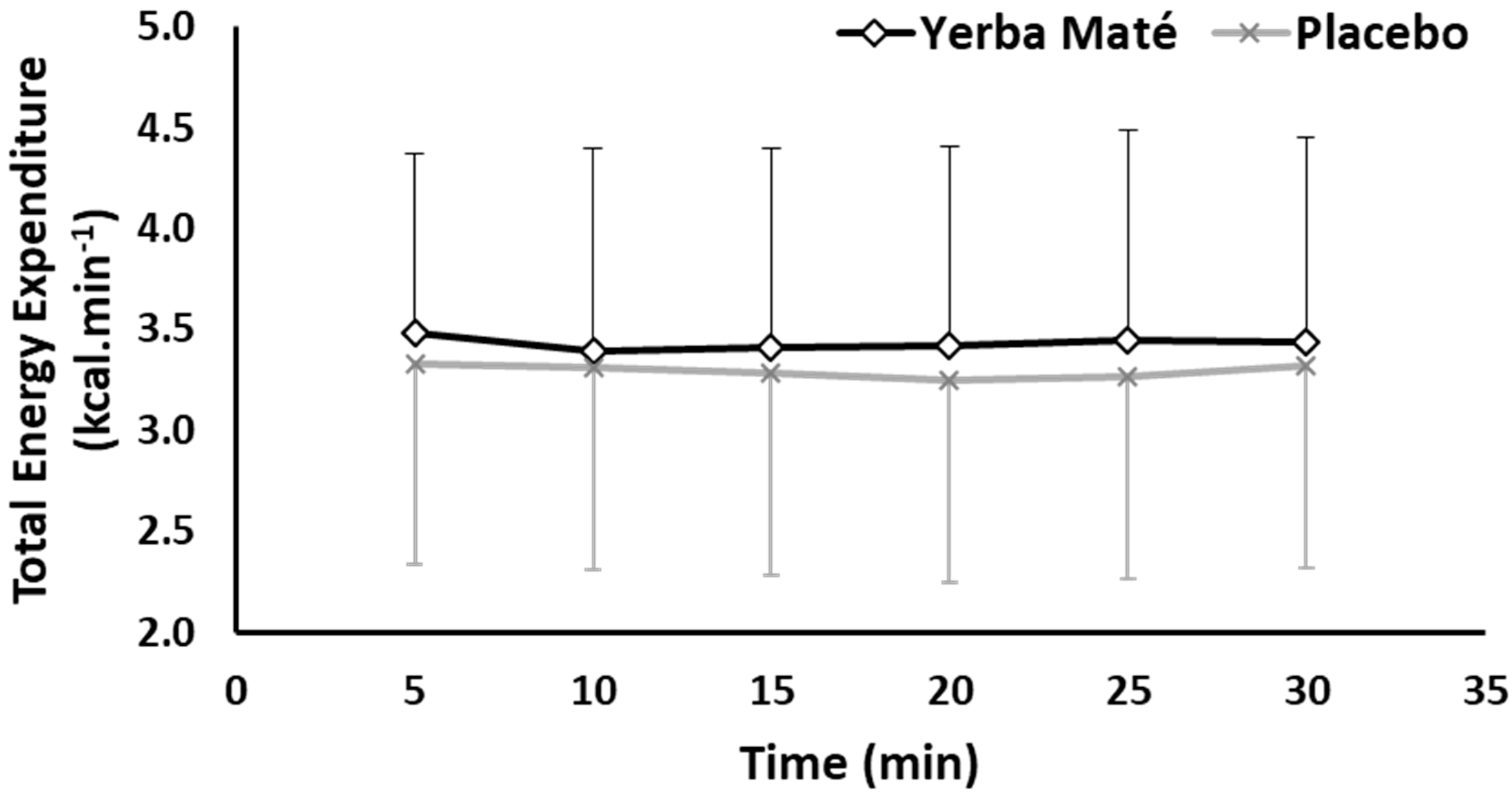
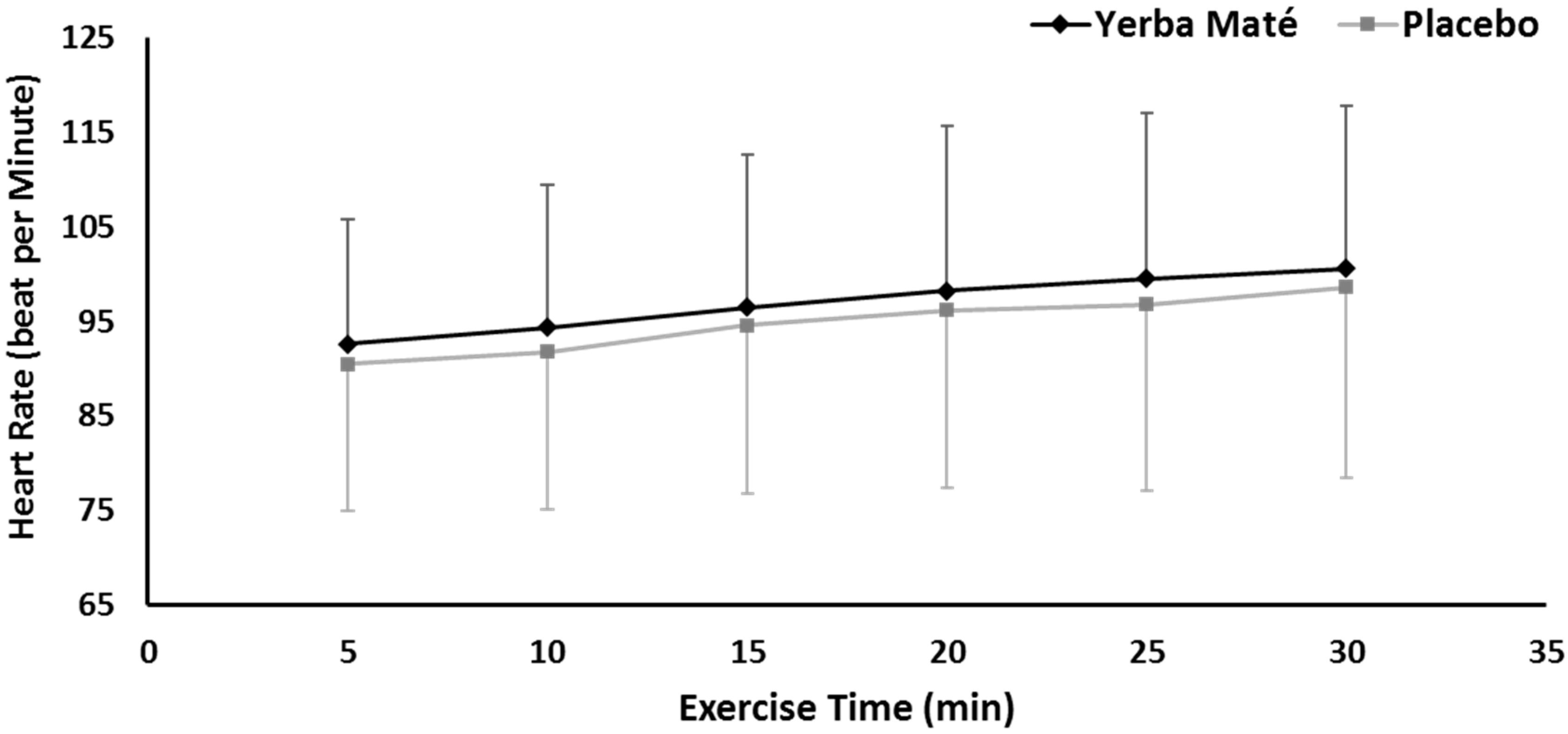
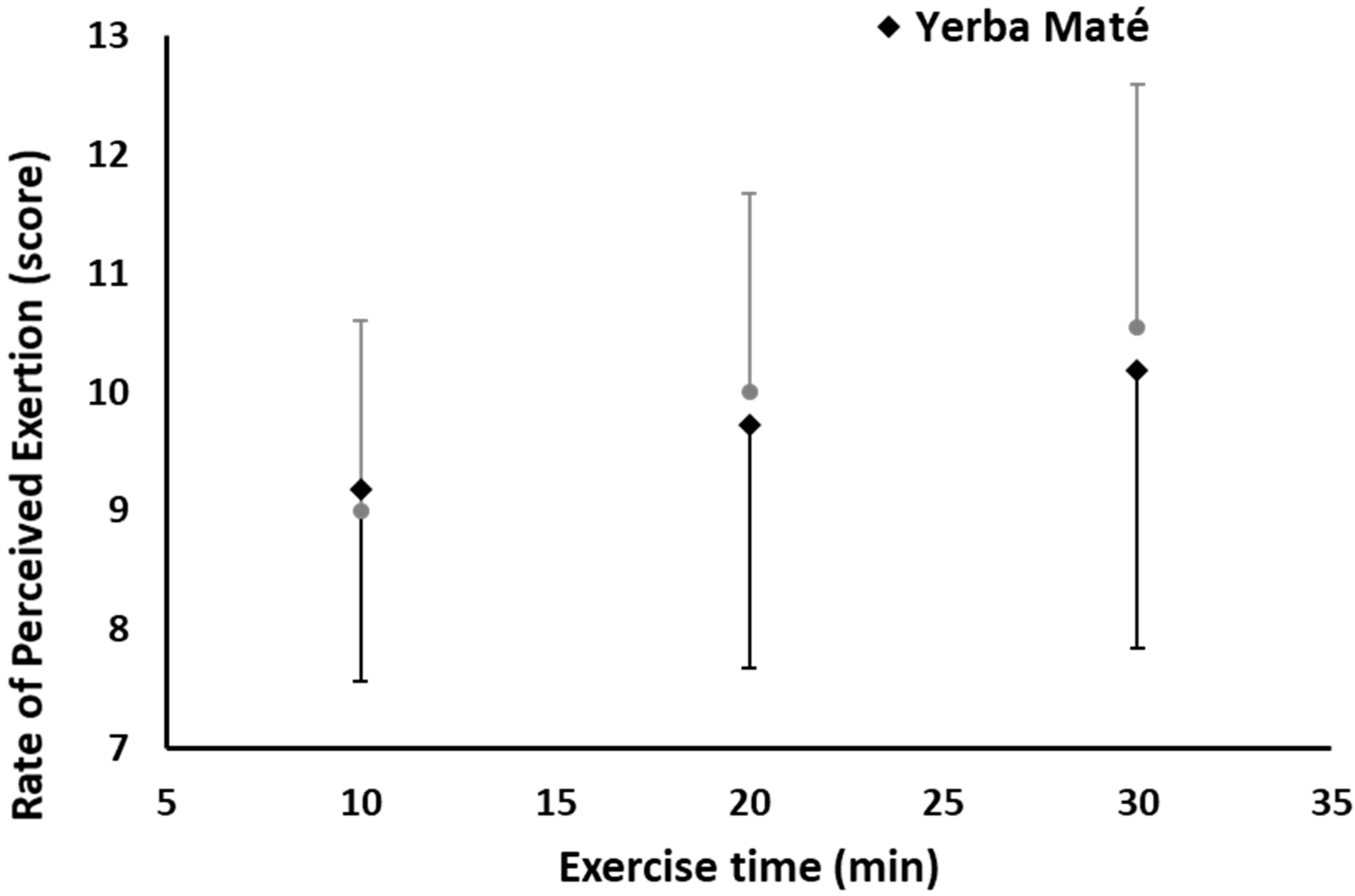
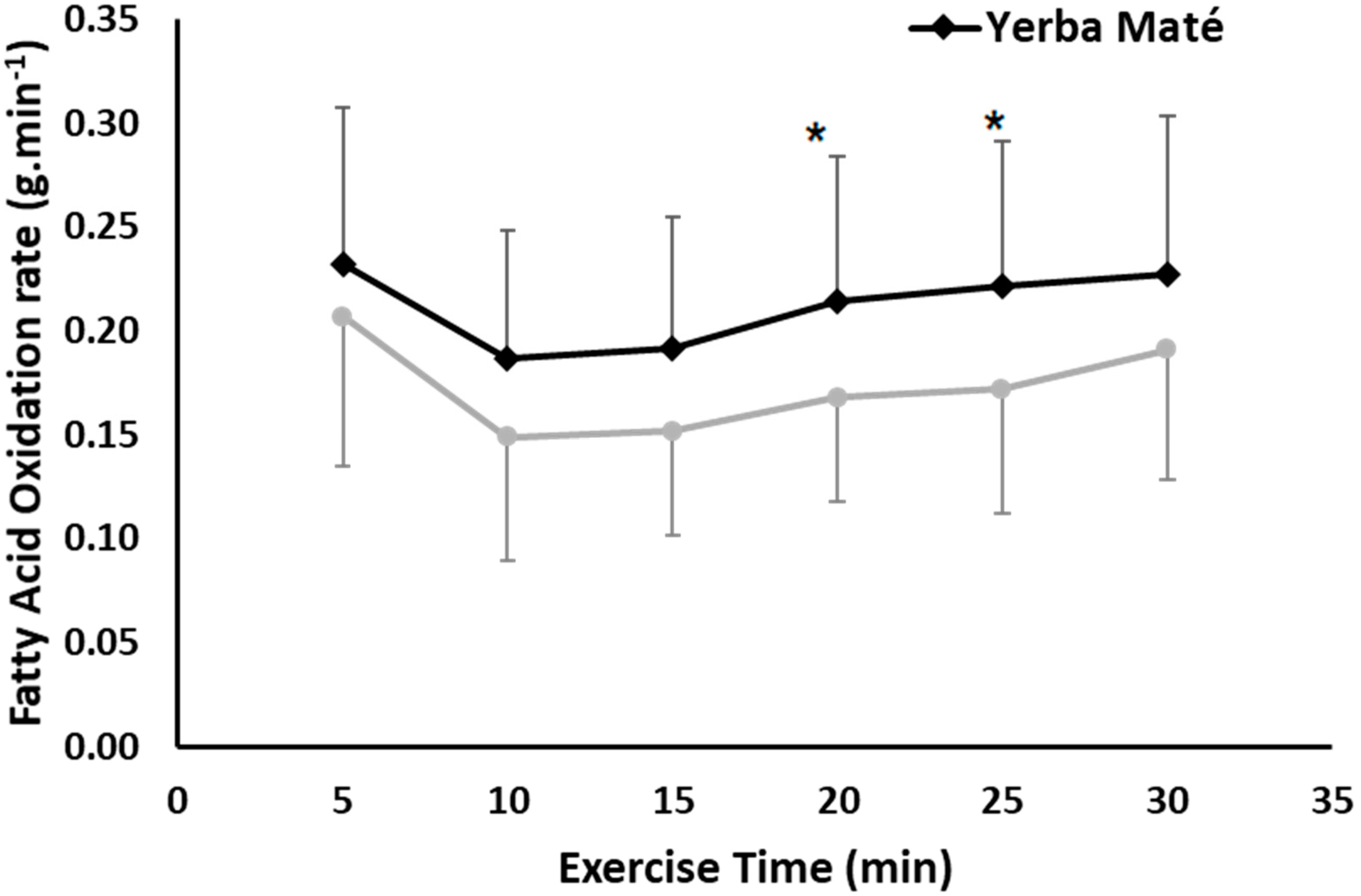
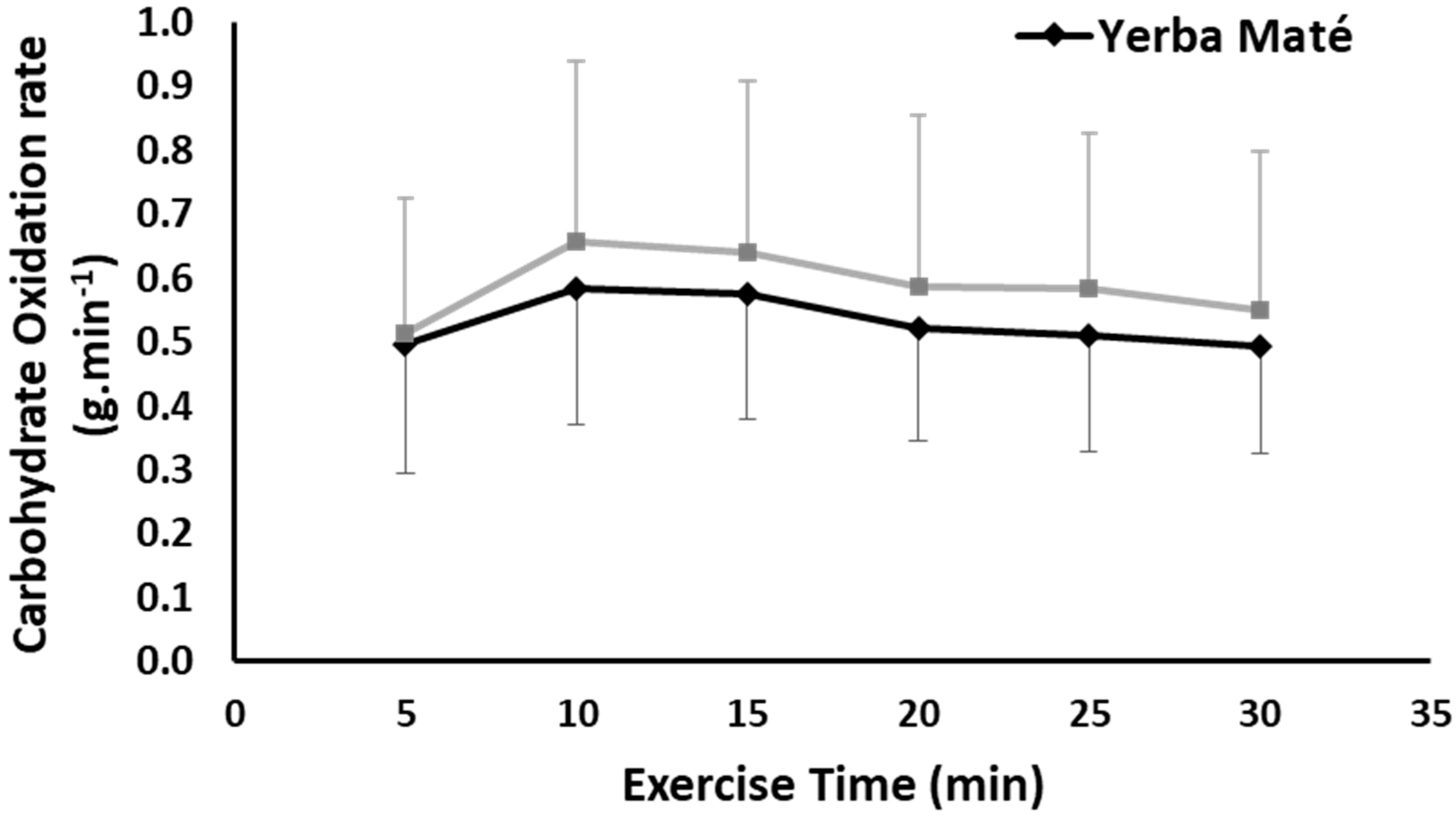
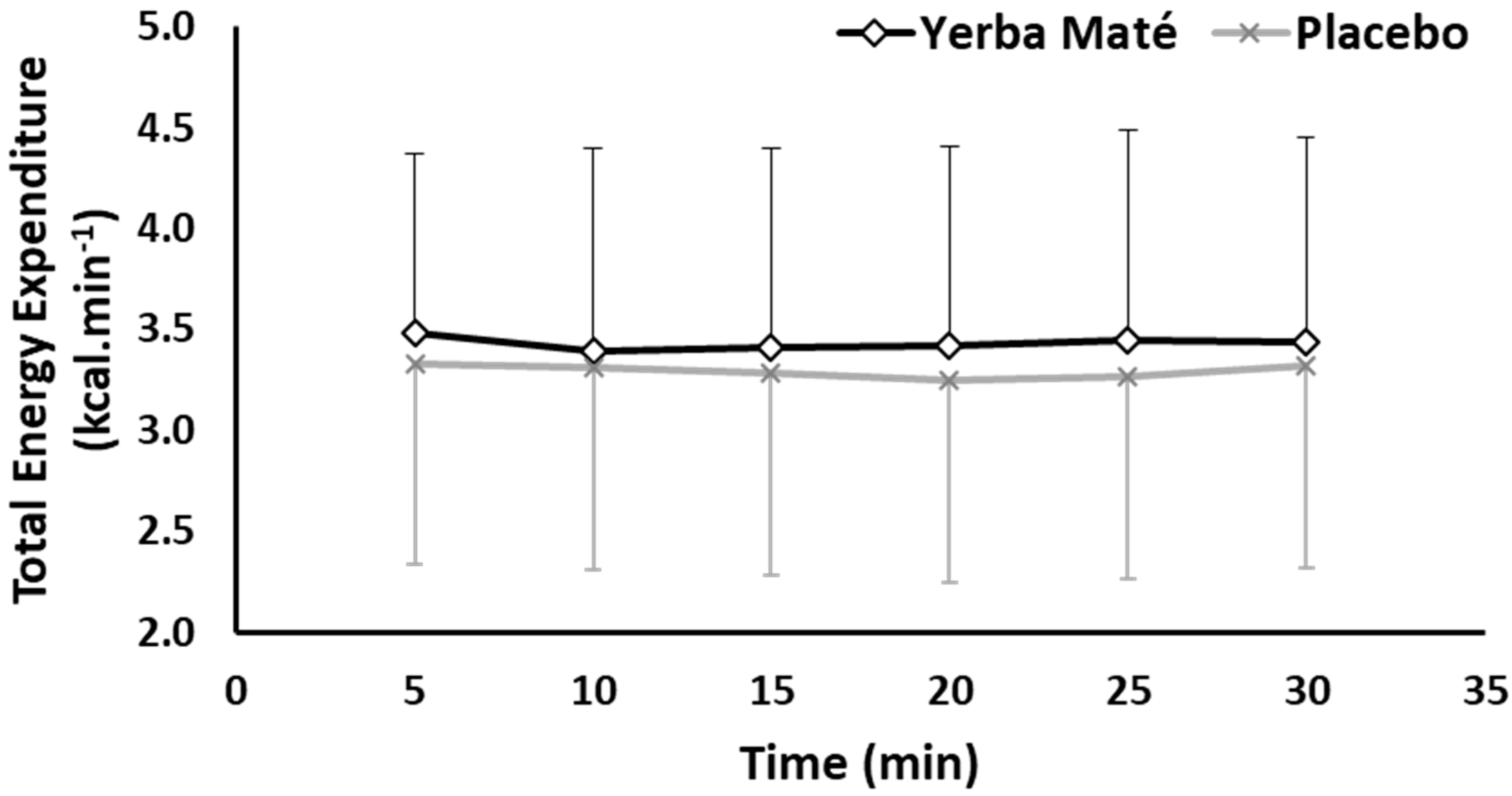
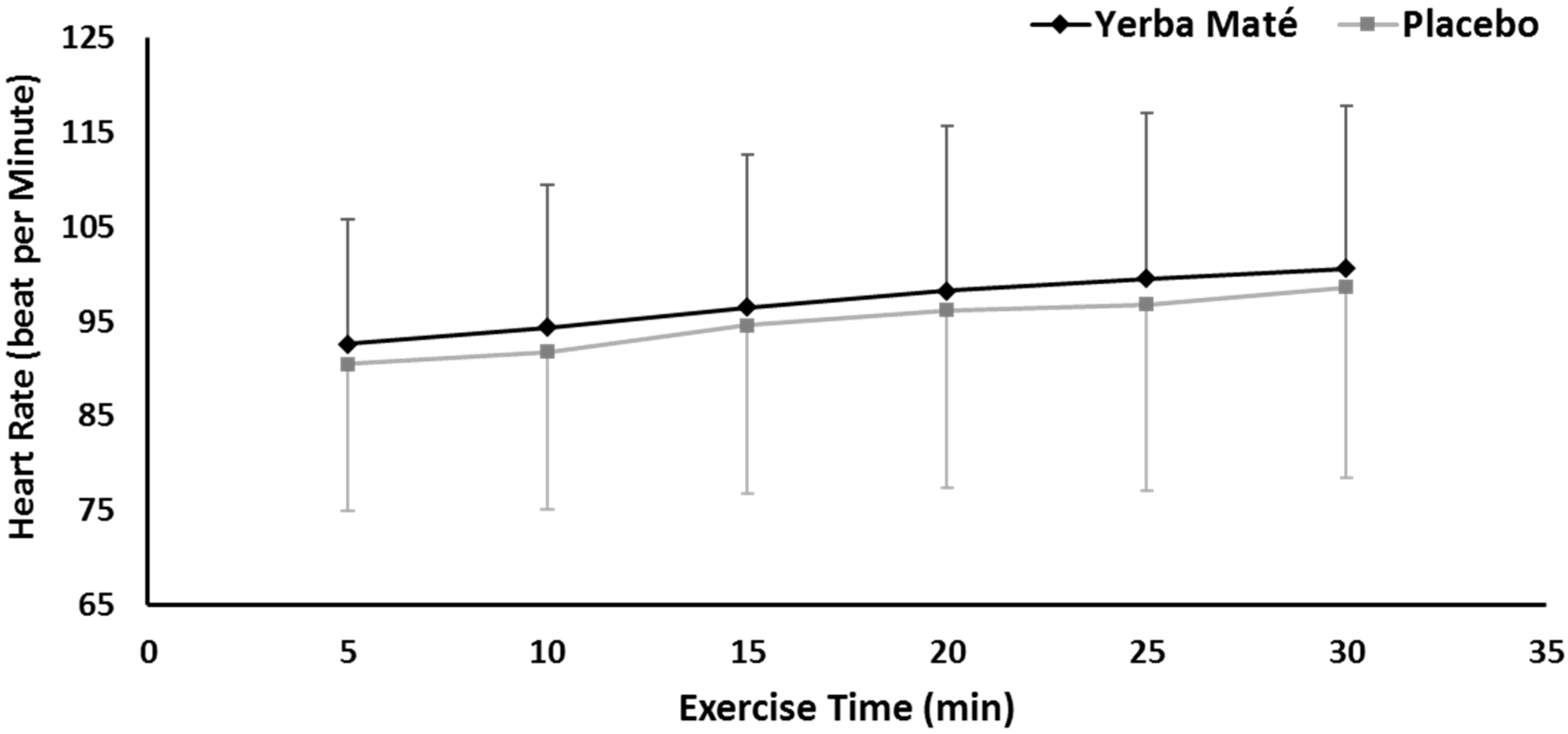
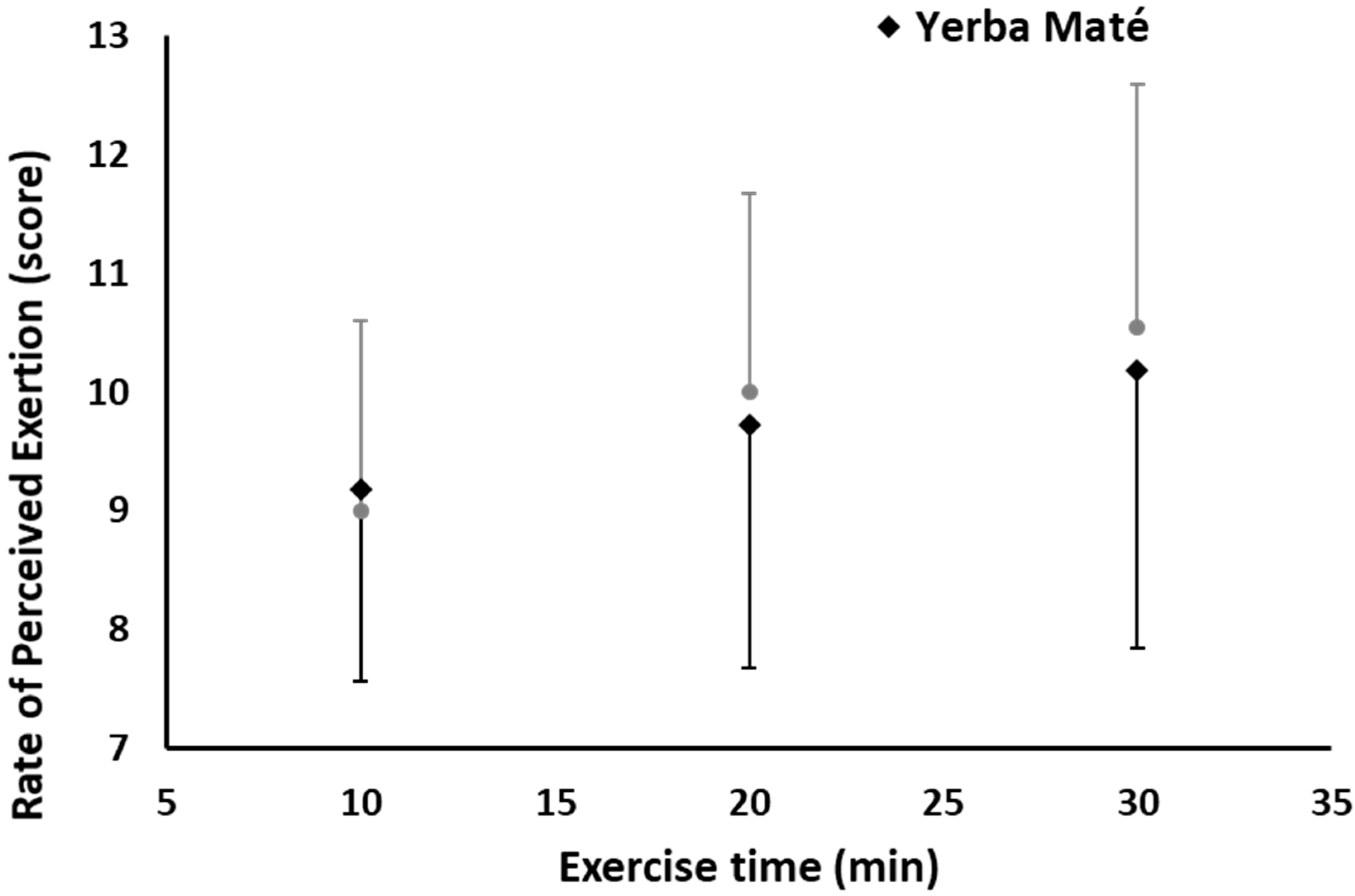
3.1. FAO, CHO, EE, and HR during Exercise
3.2. Satiety Measures of VAS
3.3. Mood State Measures of POMS
4. Discussion
5. Limitations
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sun, N.N.; Wu, T.Y.; Chau, C.F. Natural dietary and herbal products in anti-obesity treatment. Molecules 2016, 21, 1351. [Google Scholar] [CrossRef] [PubMed]
- Pillitteri, J.L.; Shiffman, S.; Rohay, J.M.; Harkins, A.M.; Burton, S.L.; Wadden, T.A. Use of dietary supplements for weight loss in The United States: Results of a national survey. Obesity 2008, 16, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Bracesco, N.; Sanchez, A.G.; Contreras, V.; Gugliucci, A. Recent advances on Ilex paraguariensis research: Minireview. J. Ethnopharmacol. 2011, 136, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Heck, C.I.; de Mejia, E.G. Yerba mate tea (Ilex paraguariensis): A comprehensive review on chemistry, health implications, and technological considerations. J. Food Sci. 2007, 72, R138–R151. [Google Scholar] [CrossRef] [PubMed]
- Bastos, D.H.M.; De Oliveira, D.M.; Matsumoto, R.L.T.; Carvalho, P.O.; Ribeiro, M.L. Yerba maté: Pharmacological properties, research and biotechnology. Med. Aromat. Plant Sci. Biotechnol. 2007, 1, 37–46. [Google Scholar]
- Borges, M.C.; Vinolo, M.A.; Nakajima, K.; de Castro, I.A.; Bastos, D.H.; Borelli, P.; Fock, R.A.; Tirapegui, J.; Curi, R.; Rogero, M.M. The effect of mate tea (Ilex paraguariensis) on metabolic and inflammatory parameters in high-fat diet-fed Wistar rats. Int. J. Food Sci. Nutr. 2013, 64, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.R.; Lee, H.Y.; Kim, J.H.; Moon, D.I.; Seo, M.Y.; Park, S.H.; Choi, K.H.; Kim, C.R.; Kim, S.H.; Oh, J.H.; et al. Anti-obesity and anti-diabetic effects of Yerba Mate (Ilex paraguariensis) in C57BL/6 J mice fed a high-fat diet. Lab. Anim. Res. 2012, 28, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Arçari, D.P.; Bartchewsky, W., Jr.; dos Santos, T.W.; Oliveira, K.A.; DeOliveira, C.C.; Gotardo, É.M.; Pedrazzoli, J., Jr.; Gambero, A.; Ferraz, L.F.; Carvalho, P.O.; et al. Anti-inflammatory effects of yerba maté extract (Ilex paraguariensis) ameliorate insulin resistance in mice with high fat diet-induced obesity. Mol. Cell. Endocrinol. 2011, 335, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Arçari, D.P.; Bartchewsky, W.; dos Santos, T.W.; Oliveira, K.A.; Funck, A.; Pedrazzoli, J.; de Souza, M.F.; Saad, M.J.; Bastos, D.H.; Gambero, A.; et al. Antiobesity effects of yerba maté extract (Ilex paraguariensis) in high-fat diet-induced obese mice. Obesity 2009, 17, 2127–2133. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Ko, J.; Storni, C.; Song, H.J.; Cho, Y.G. Effect of green mate in overweight volunteers: A randomized placebo-controlled human study. J. Funct. Foods 2012, 4, 287–293. [Google Scholar] [CrossRef]
- Kim, S.Y.; Oh, M.R.; Kim, M.G.; Chae, H.J.; Chae, S.W. Anti-obesity effects of Yerba Mate (Ilex paraguariensis): A randomized, double-blind, placebo-controlled clinical trial. BMC Complement. Altern. Med. 2015, 15, 338. [Google Scholar] [CrossRef] [PubMed]
- Andersen, T.; Fogh, J. Weight loss and delayed gastric emptying following a South American herbal preparation in overweight patients. J. Hum. Nutr. Diet. 2001, 14, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Hussein, G.M.E.; Matsuda, H.; Nakamura, S.; Hamao, M.; Akiyama, T.; Tamura, K.; Yoshikawa, M. Mate tea (Ilex paraguariensis) promotes satiety and body weight lowering in mice: Involvement of glucagon-like peptide-1. Biol. Pharm. Bull. 2011, 34, 1849–1855. [Google Scholar] [CrossRef] [PubMed]
- Yimam, M.; Jiao, P.; Hong, M.; Brownell, L.; Lee, Y.C.; Hyun, E.J.; Kim, H.J.; Kim, T.W.; Nam, J.B.; Kim, M.R.; et al. Appetite suppression and antiobesity effect of a botanical composition composed of Morus alba, Yerba mate, and Magnolia officinalis. J. Obes. 2016, 2016, 4670818. [Google Scholar] [CrossRef] [PubMed]
- Alkhatib, A.; Seijo, M.; Larumbe, E.; Naclerio, F. Acute effectiveness of a “fat-loss” product on substrate utilization, perception of hunger, mood state and rate of perceived exertion at rest and during exercise. J. Int. Soc. Sports Nutr. 2015, 25, 44. [Google Scholar] [CrossRef] [PubMed]
- Outlaw, J.; Wilborn, C.; Smith, A.; Urbina, S.; Hayward, S.; Foster, C.; Wells, S.; Wildman, R.; Taylor, L. Effects of ingestion of a commercially available thermogenic dietary supplement on resting energy expenditure, mood state and cardiovascular measures. J. Int. Soc. Sports Nutr. 2013, 10, 25. [Google Scholar] [CrossRef] [PubMed]
- Dalle Grave, R.; Centis, E.; Marzocchi, R.; El Ghoch, M.; Marchesini, G. Major factors for facilitating change in behavioral strategies to reduce obesity. Psychol. Res. Behav. Manag. 2013, 6, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Mazzeschi, C.; Piana, N.; Capezzali, D.; Mommi, A.; Aiello, C.; Gatti, M.; Romani, G.; Buratta, L.; Battistini, D.; Nasini, G.; et al. The impact of strenuous group physical activity on mood states, personal views, body composition, and markers of myocardial damage in overweight/obese adults: The “Step-by-step Italy’s coast to coast” trek. Biomed Res. Int. 2014, 2014, 854129. [Google Scholar] [CrossRef] [PubMed]
- Mama, S.K.; Schembre, S.M.; O’Connor, D.P.; Kaplan, C.D.; Bode, S.; Lee, R.E. Effectiveness of lifestyle interventions to reduce binge eating symptoms in African American and Hispanic women. Appetite 2015, 95, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Kanning, M.; Schlicht, W. Be active and become happy: An ecological momentary assessment of physical activity and mood. J. Sport Exerc. Psychol. 2010, 32, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Martinet, A.; Hostettmann, K.; Schutz, Y. Thermogenic effects of commercially available plant preparations aimed at treating human obesity. Phytomedicine 1999, 6, 231–238. [Google Scholar] [CrossRef]
- Alkhatib, A. Yerba maté (Illex Paraguariensis) ingestion augments fat oxidation and energy expenditure during exercise at various submaximal intensities. Nutr. Metab. 2014, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Achten, J.; Gleeson, M.; Jeukendrup, A.E. Determination of the exercise intensity that elicits maximal fat oxidation. Med. Sci. Sports Exerc. 2002, 34, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A.; Mercier, J. Balance of carbohydrate and lipid utilization during exercise: The “crossover” concept. J. Appl. Physiol. 1994, 76, 2253–2261. [Google Scholar] [PubMed]
- Venables, M.C.; Jeukendrup, A.E. Endurance training and obesity: Effect on substrate metabolism and insulin sensitivity. Med. Sci. Sports Exerc. 2008, 40, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Alkhatib, A.; Klonizakis, M. Effects of exercise training and Mediterranean diet on vascular risk reduction in post-menopausal women. Clin. Hemorheol. Microcirc. 2014, 57, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Venables, M.C.; Hulston, C.J.; Cox, H.R.; Jeukendrup, A.E. Green tea extract ingestion, fat oxidation and glucose tolerance in healthy humans. Am. J. Clin. Nutr. 2008, 87, 778–784. [Google Scholar] [PubMed]
- Venables, M.C.; Achten, J.; Jeukendrup, A.E. Determinants of fat oxidation during exercise in healthy men and women: A cross-sectional study. J. Appl. Physiol. 2005, 98, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Borg, G.A. Psychophysical bases of perceived exertion. Med. Sci. Sports Exerc. 1982, 14, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Blundell, J.; de Graaf, C.; Hulshof, T.; Jebb, S.; Livingstone, B.; Lluch, A.; Mela, D.; Salah, S.; Schuring, E.; van der Knaap, H.; et al. Appetite control: Methodological aspects of the evaluation of foods. Obes. Rev. 2010, 11, 251–270. [Google Scholar] [CrossRef] [PubMed]
- Morgan, W.P.; Brown, D.R.; Raglin, J.S.; O’Connor, P.J.; Ellickson, K.A. Psychological monitoring of overtraining and staleness. Br. J. Sports Med. 1987, 21, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Brun, J.F.; Jean, E.; Ghanassia, E.; Flavier, S.; Mercier, J. Metabolic training: New paradigms of exercise training for metabolic diseases with exercise calorimetry targeting individuals. Ann. Readapt. Med. Phys. 2007, 50, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Martin, A.; Dumortier, M.; Raynaud, E.; Brun, J.F.; Fédou, C.; Bringer, J.; Mercier, J. Balance of substrate oxidation during submaximal exercise in lean and obese people. Diabetes Metab. 2001, 27, 466–474. [Google Scholar] [PubMed]
- Hoffman, J.R.; Kang, J.; Ratamess, N.A.; Rashti, S.L.; Tranchina, C.P.; Faigenbaum, A.D. Thermogenic effect of an acute ingestion of a weight loss supplement. J. Int. Soc. Sports Nutr. 2009, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Gahreman, D.; Wang, R.; Boutcher, Y.; Boutcher, S. Green tea, intermittent sprinting exercise, and fat oxidation. Nutrients 2015, 7, 5646–5663. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, A.B.; Randell, R.K.; Boon, N.; Garczarek, U.; Mela, D.J.; Jeukendrup, A.E.; Jacobs, D.M. Metabolic response to green tea extract during rest and moderate-intensity exercise. J. Nutr. Biochem. 2013, 24, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Woodward, K.A.; Hopkins, N.D.; Draijer, R.; de Graaf, Y.; Low, D.A.; Thijssen, D.H. Acute black tea consumption improves cutaneous vascular function in healthy middle-aged humans. Clin. Nutr. 2016. [Google Scholar] [CrossRef] [PubMed]
- Alghadir, A.H.; Gabr, S.A.; Al-Eisa, E.S. Green tea and exercise interventions as nondrug remedies in geriatric patients with rheumatoid arthritis. J. Phys. Ther. Sci. 2016, 28, 2820–2829. [Google Scholar] [CrossRef] [PubMed]
- Panza, V.P.; Diefenthaeler, F.; Tamborindeguy, A.C.; de Camargo, C.Q.; de Moura, B.M.; Brunetta, H.S.; Sakugawa, R.L.; de Oliveira, M.V.; Puel Ede, O.; Nunes, E.A.; et al. Effects of mate tea consumption on muscle strength and oxidative stress markers after eccentric exercise. Br. J. Nutr. 2016, 115, 1370–1378. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, E.S.; Slettenaar, M.; Meer, V.; Transler, C.; Jans, L.; Quadt, F.; Berry, M. Differential contributions of theobromine and caffeine on mood psychomotor performance and blood pressure. Physiol. Behav. 2011, 104, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Howe, S.M.; Hand, T.M.; Larson-Meyer, D.E.; Austin, K.J.; Alexander, B.M.; Manore, M.M. No effect of exercise intensity on appetite in highly-trained endurance women. Nutrients 2016, 8, 223. [Google Scholar] [CrossRef] [PubMed]
- Duckworth, L.C.; Backhouse, S.H.; O’Hara, J.P.; Stevenson, E.J. Effect of galactose ingestion before and during exercise on substrate oxidation, postexercise satiety, and subsequent energy intake in females. J. Am. Coll. Nutr. 2016, 35, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Muccillo, B.A.L.; Johnston, K.B.; Paganini, S.F.L. Endothelium-dependent vasorelaxing activity of aqueous extracts of Ilex paraguariensis on mesenteric arterial bed of rats. J. Ethnopharmacol. 1998, 60, 133–139. [Google Scholar] [CrossRef]
- Yu, S.; Yue, S.W.; Liu, Z.; Zhang, T.; Xiang, N.; Fu, H. Yerba mate (Ilex paraguariensis) improves microcirculation of volunteers with high blood viscosity: A randomized, double-blind, placebo-controlled trial. Exp. Gerontol. 2015, 62, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.D.; Bueno, A.L.; Gallon, C.W.; Gomes, L.F.; Kaiser, S.; Pavei, C.; Ortega, G.G.; Kucharski, L.C.; Jahn, M.P. The effect of aqueous extract of gross and commercial yerba mate (Ilex paraguariensis) on intra-abdominal and epididymal fat and glucose levels in male Wistar rats. Fitoterapia 2011, 82, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Horton, T.J.; Miller, E.K.; Glueck, D.; Tench, K. No effect of menstrual cycle phase on glucose kinetics and fuel oxidation during moderate-intensity exercise. Am. J. Physiol. Endocrinol. Metab. 2002, 282, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Blaak, E. Gender differences in fat metabolism. Curr. Opin. Clin. Nutr. Metab. Care 2001, 4, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Isacco, L.; Duché, P.; Boisseau, N. Influence of hormonal status on substrate utilization at rest and during exercise in the female population. Sports Med. 2012, 42, 327–342. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VAS Measures | Time (Min) | Score for YM | Score for PLC |
|---|---|---|---|
| Hunger | 0 | 5.34 ± 2.62 | 4.76 ± 2.18 |
| 30 | 5.01 ± 2.30 | 5.21 ± 2.74 | |
| 60 | 5.32 ± 2.43 | 5.80 ± 2.69 | |
| 90 | 5.85 ± 2.42 | 6.07 ± 2.23 | |
| 120 | 6.08 ± 2.16 | 5.83 ± 2.04 | |
| 150 | 5.89 ± 2.38 | 5.50 ± 2.29 | |
| 180 | 6.45 ± 2.52 | 6.88 ± 1.76 | |
| Fullness | 0 | 2.06 ± 1.45 | 1.63 ± 1.16 |
| 30 | 2.11 ± 1.37 | 1.73 ± 1.38 | |
| 60 | 1.55 ± 1.22 | 1.64 ± 1.36 | |
| 90 | 1.43 ± 1.05 | 1.67 ± 1.55 | |
| 120 | 1.55 ± 1.26 | 1.60 ± 1.27 | |
| 150 | 1.77 ± 2.17 | 1.46 ± 1.18 | |
| 180 | 1.94 ± 2.49 | 1.02 ± 0.96 | |
| Desire to Eat | 0 | 5.45 ± 2.57 | 5.67 ± 2.08 |
| 30 | 5.37 ± 2.25 | 6.10 ± 2.41 | |
| 60 | 5.96 ± 2.54 | 6.08 ± 2.49 | |
| 90 | 6.07 ± 2.39 | 6.54 ± 2.09 | |
| 120 | 5.76 ± 2.09 | 6.43 ± 1.89 | |
| 150 | 5.99 ± 2.70 | 5.74 ± 2.62 | |
| 180 | 6.34 ± 2.35 | 7.07 ± 1.95 | |
| Prospective Consumption | 0 | 4.30 ± 1.90 | 4.65 ± 1.89 |
| 30 | 4.47 ± 1.63 | 4.68 ± 1.86 | |
| 60 | 5.01 ± 2.02 | 4.80 ± 1.81 | |
| 90 | 4.97 ± 1.95 | 5.16 ± 1.62 | |
| 120 | 5.17 ± 1.96 | 5.36 ± 1.43 | |
| 150 | 5.21 ± 1.82 | 4.87 ± 2.17 | |
| 180 | 5.28 ± 1.59 | 5.64 ± 1.66 |
| POMS Measures | Time (Min) | Score for YM | Score for PLC |
|---|---|---|---|
| Alertness | 0 | 3.18 ± 0.98 | 3.18 ± 1.17 |
| 30 | 3.27 ± 0.79 | 2.82 ± 0.87 | |
| 60 | 3.36 ± 0.50 | 3.00 ± 1.00 | |
| 90 | 3.09 ± 0.83 | 2.73 ± 0.79 | |
| 120 | 3.18 ± 0.60 | 2.82 ± 0.87 | |
| 150 | 3.73 ± 0.79 | 3.73 ± 0.65 | |
| 180 | 3.36 ± 0.92 | 3.00 ± 1.00 | |
| Focus | 0 | 3.45 ± 1.04 | 3.36 ± 1.21 |
| 30 | 3.36 ± 1.03 | 2.82 ± 0.98 | |
| 60 | 3.18 ± 0.75 | 2.91 ± 1.14 | |
| 90 | 3.09 ± 0.70 | 2.82 ± 1.08 | |
| 120 | 3.18 ± 0.60 | 2.91 ± 0.70 | |
| 150 | 3.18 ± 0.60 | 3.45 ± 0.82 | |
| 180 | 2.82 ± 0.75 | 2.91 ± 0.94 | |
| Energy | 0 | 3.00 ± 1.00 | 3.09 ± 0.94 |
| 30 | 3.00 ± 1.10 | 2.82 ± 0.87 | |
| 60 | 3.00 ± 0.63 | 2.82 ± 0.87 | |
| 90 | 2.91 ± 0.70 | 2.91 ± 0.83 | |
| 120 | 2.82 ± 0.60 | 2.82 ± 0.75 | |
| 150 | 3.27 ± 0.47 | 3.73 ± 0.79 | |
| 180 | 2.64 ± 0.67 | 3.00 ± 0.89 | |
| Fatigue | 0 | 2.73 ± 0.90 | 2.73 ± 1.01 |
| 30 | 2.45 ± 0.82 | 2.91 ± 0.83 | |
| 60 | 2.55 ± 0.82 | 2.82 ± 0.60 | |
| 90 | 2.55 ± 0.93 | 2.82 ± 0.60 | |
| 120 | 2.64 ± 0.81 | 2.55 ± 0.69 | |
| 150 | 2.36 ± 0.81 | 2.18 ± 0.75 | |
| 180 | 2.64 ± 0.92 | 2.73 ± 0.79 | |
| Concentration | 0 | 3.09 ± 1.04 | 3.27 ± 1.10 |
| 30 | 3.00 ± 1.00 | 2.91 ± 1.14 | |
| 60 | 3.27 ± 0.79 | 2.91 ± 1.14 | |
| 90 | 2.91 ± 0.70 | 2.82 ± 1.08 | |
| 120 | 2.91 ± 0.70 | 2.73 ± 0.90 | |
| 150 | 3.18 ± 0.98 | 3.55 ± 0.93 | |
| 180 | 2.64 ± 0.81 | 2.82 ± 0.87 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkhatib, A.; Atcheson, R. Yerba Maté (Ilex paraguariensis) Metabolic, Satiety, and Mood State Effects at Rest and during Prolonged Exercise. Nutrients 2017, 9, 882. https://doi.org/10.3390/nu9080882
Alkhatib A, Atcheson R. Yerba Maté (Ilex paraguariensis) Metabolic, Satiety, and Mood State Effects at Rest and during Prolonged Exercise. Nutrients. 2017; 9(8):882. https://doi.org/10.3390/nu9080882
Chicago/Turabian StyleAlkhatib, Ahmad, and Roisin Atcheson. 2017. "Yerba Maté (Ilex paraguariensis) Metabolic, Satiety, and Mood State Effects at Rest and during Prolonged Exercise" Nutrients 9, no. 8: 882. https://doi.org/10.3390/nu9080882
APA StyleAlkhatib, A., & Atcheson, R. (2017). Yerba Maté (Ilex paraguariensis) Metabolic, Satiety, and Mood State Effects at Rest and during Prolonged Exercise. Nutrients, 9(8), 882. https://doi.org/10.3390/nu9080882