Spirulina Protects against Hepatic Inflammation in Aging: An Effect Related to the Modulation of the Gut Microbiota?

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diet Intervention
2.2. Gut Microbiota Analyses
2.3. Biochemical Analysis
2.4. Tissue mRNA Analyses
2.5. TLR2 and TLR4 Agonists
2.6. Statistical Analysis
3. Results
3.1. Spirulina Did Not Change Body Weight Gain, Food Intake, and Organ Weights of Old Mice
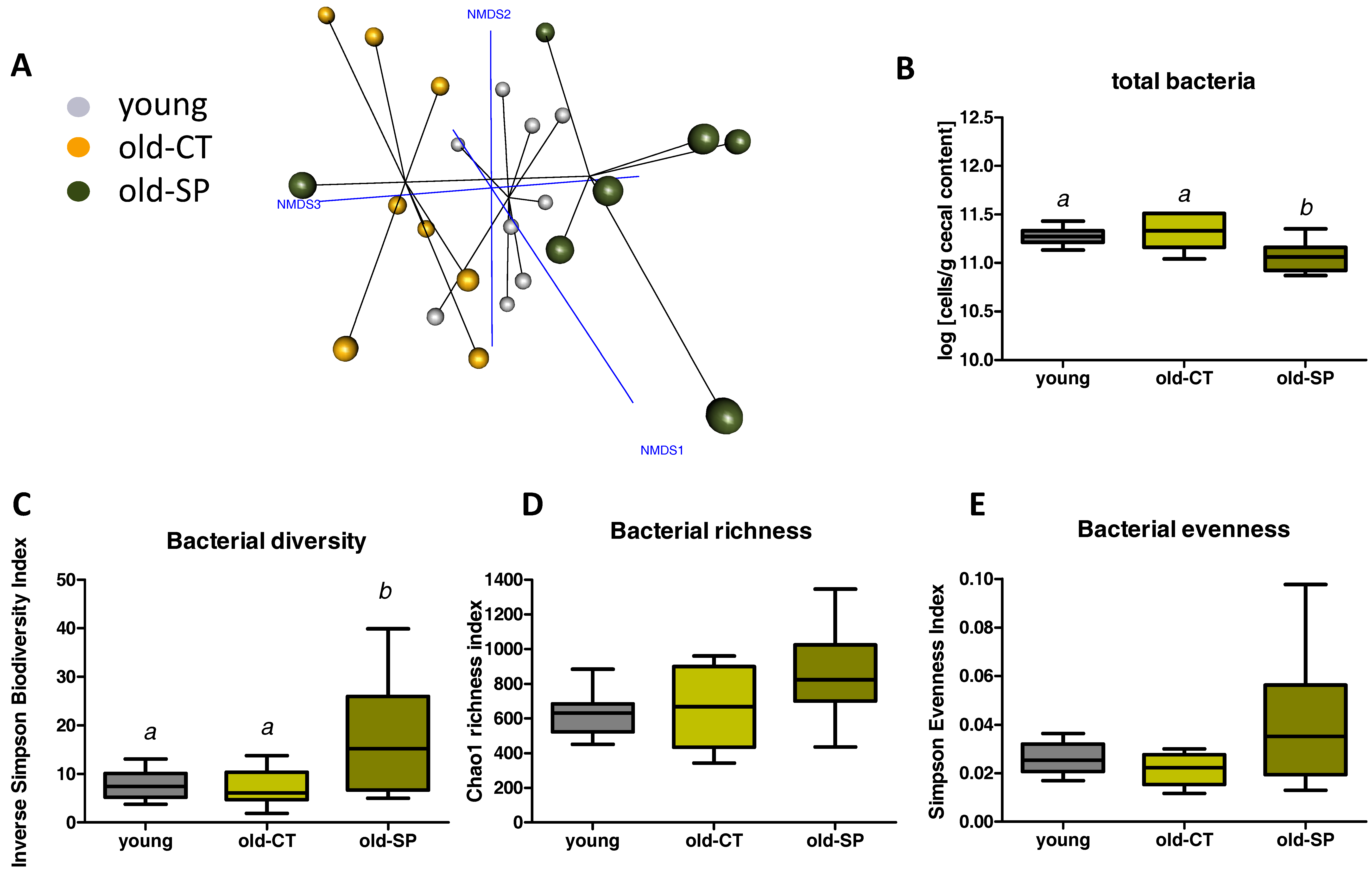
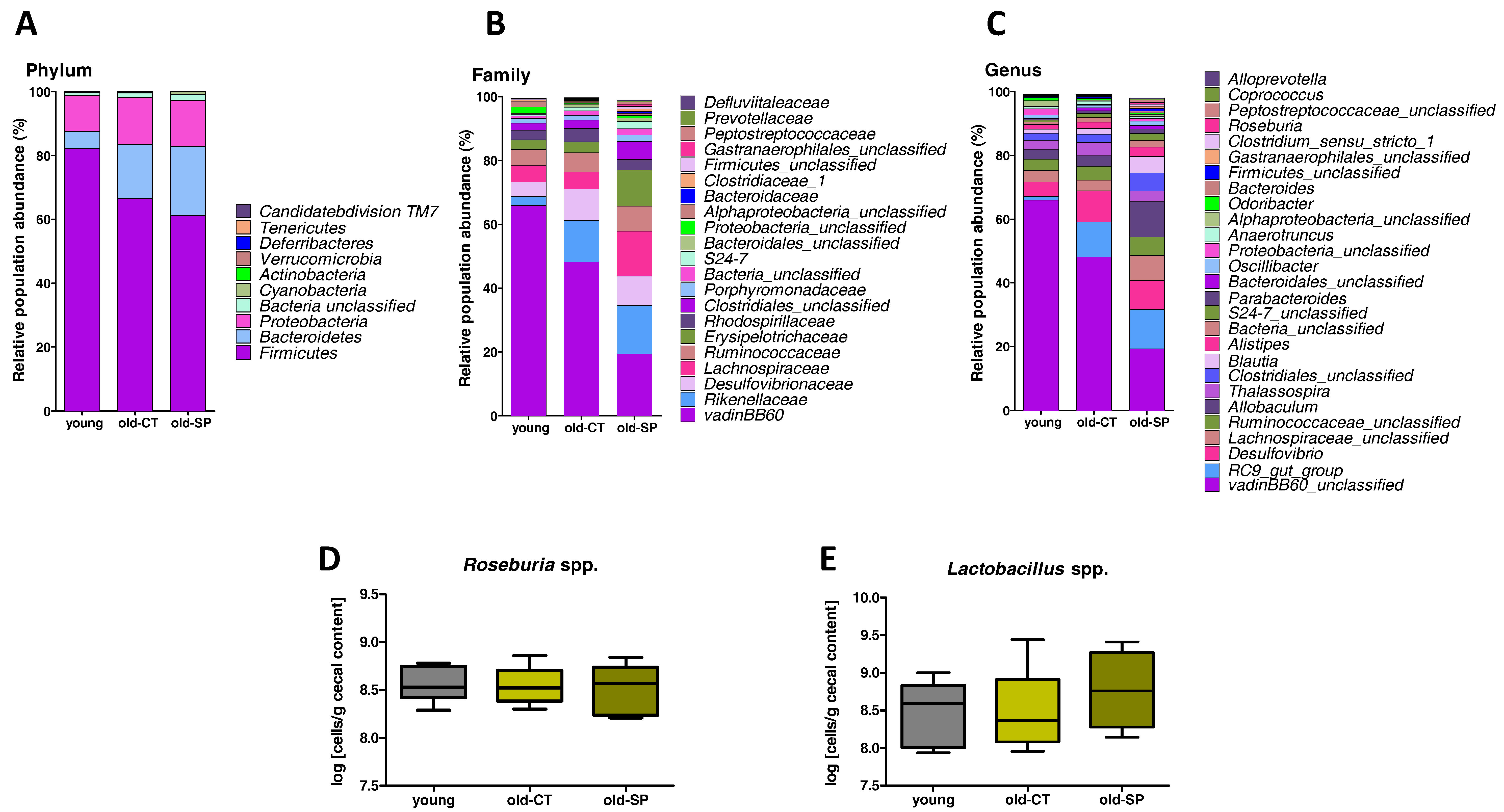
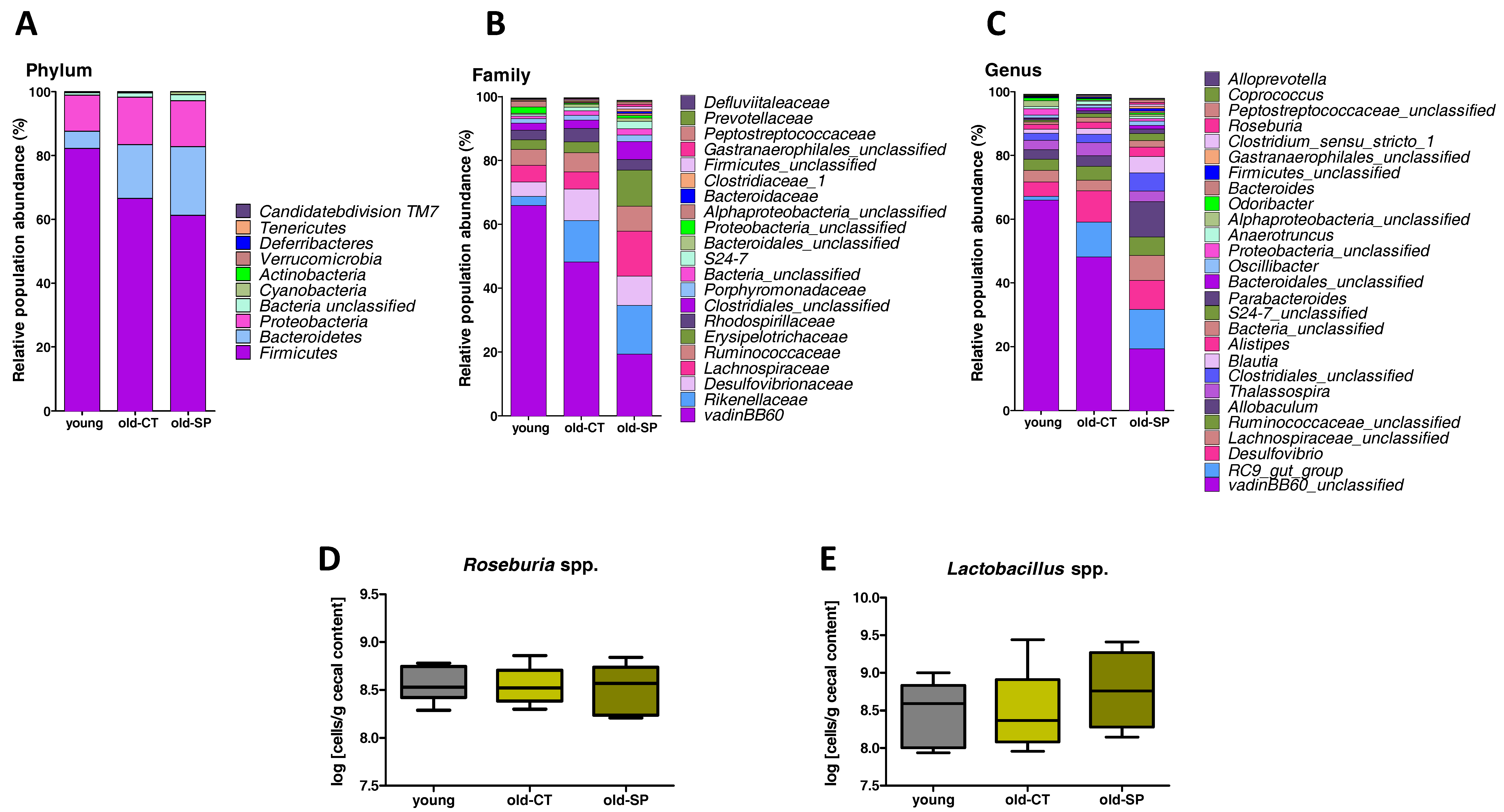
3.2. Spirulina Changed Microbial Diversity and Populations in Old Mice
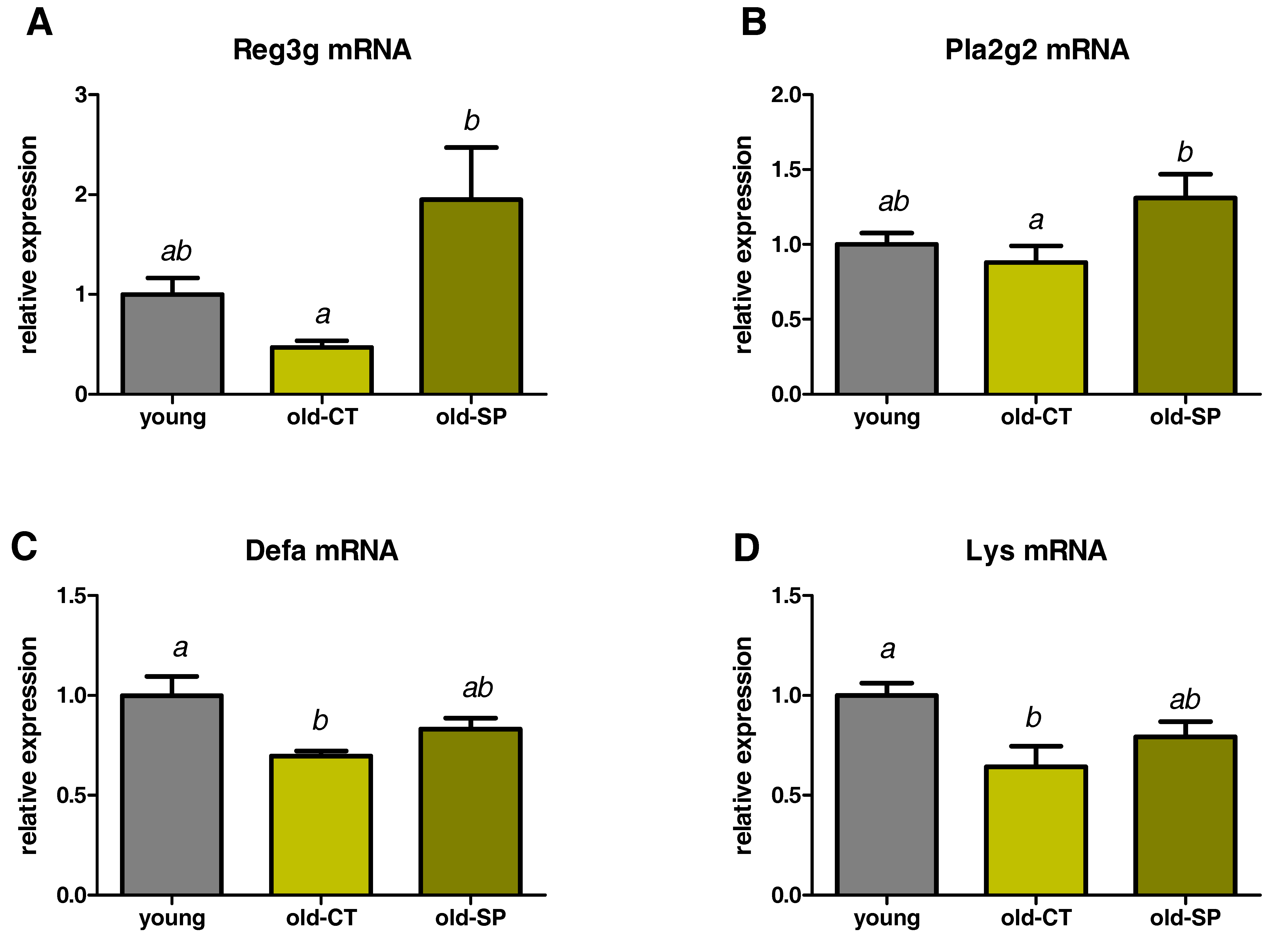
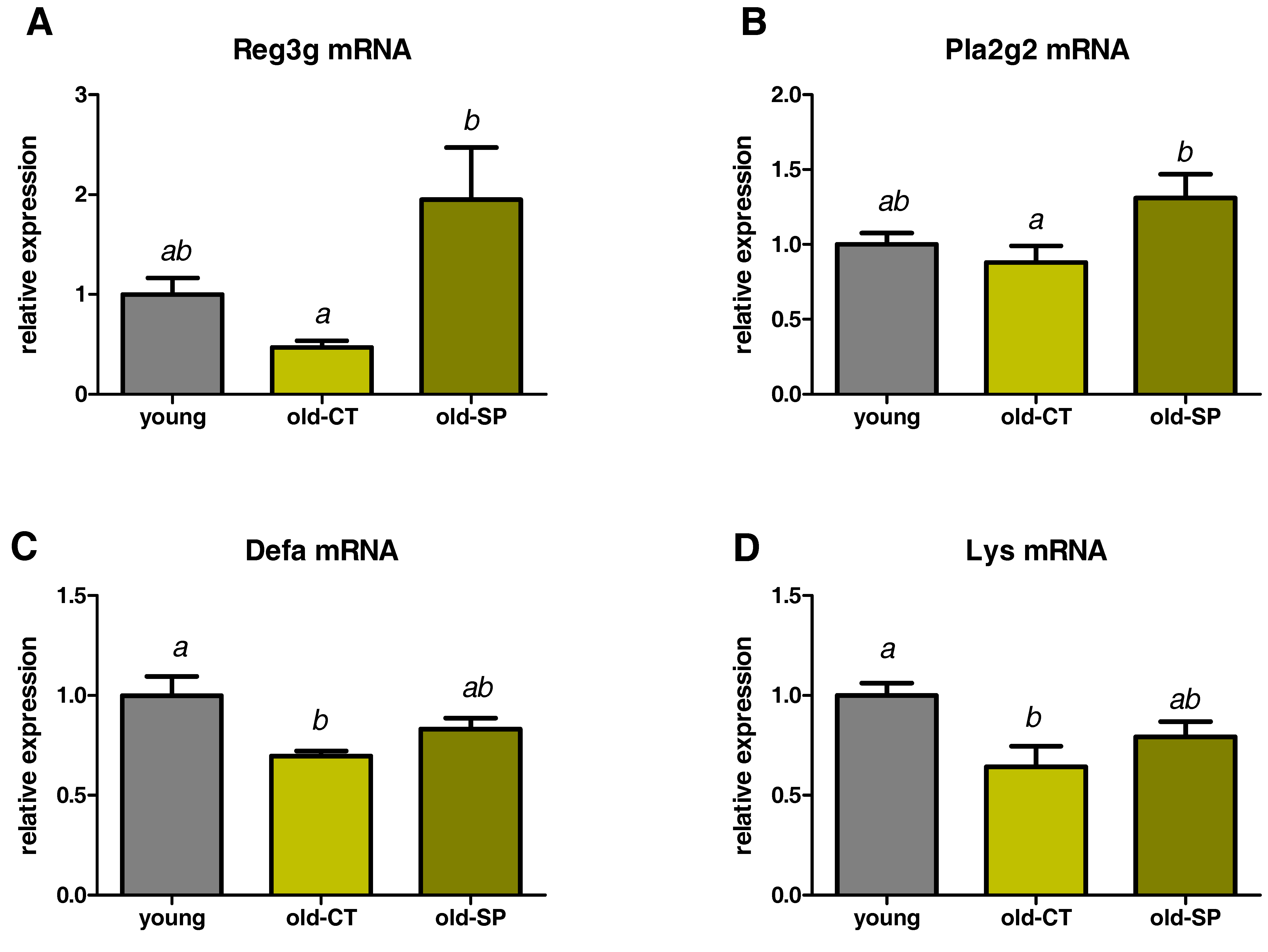
3.3. Spirulina Upregulated the Expression of Antimicrobial Peptides (AMPs) in the Small Intestine of Old Mice
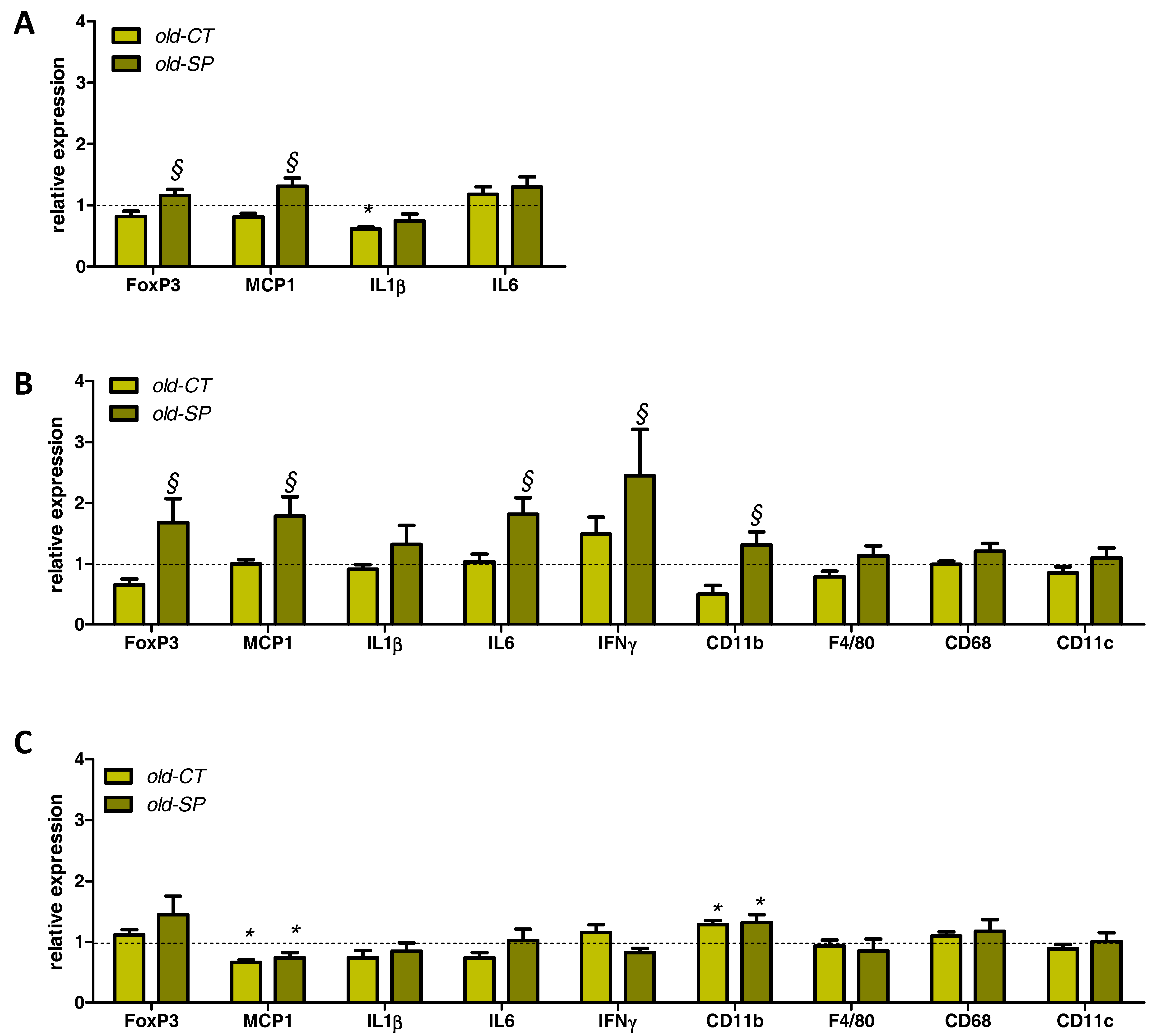
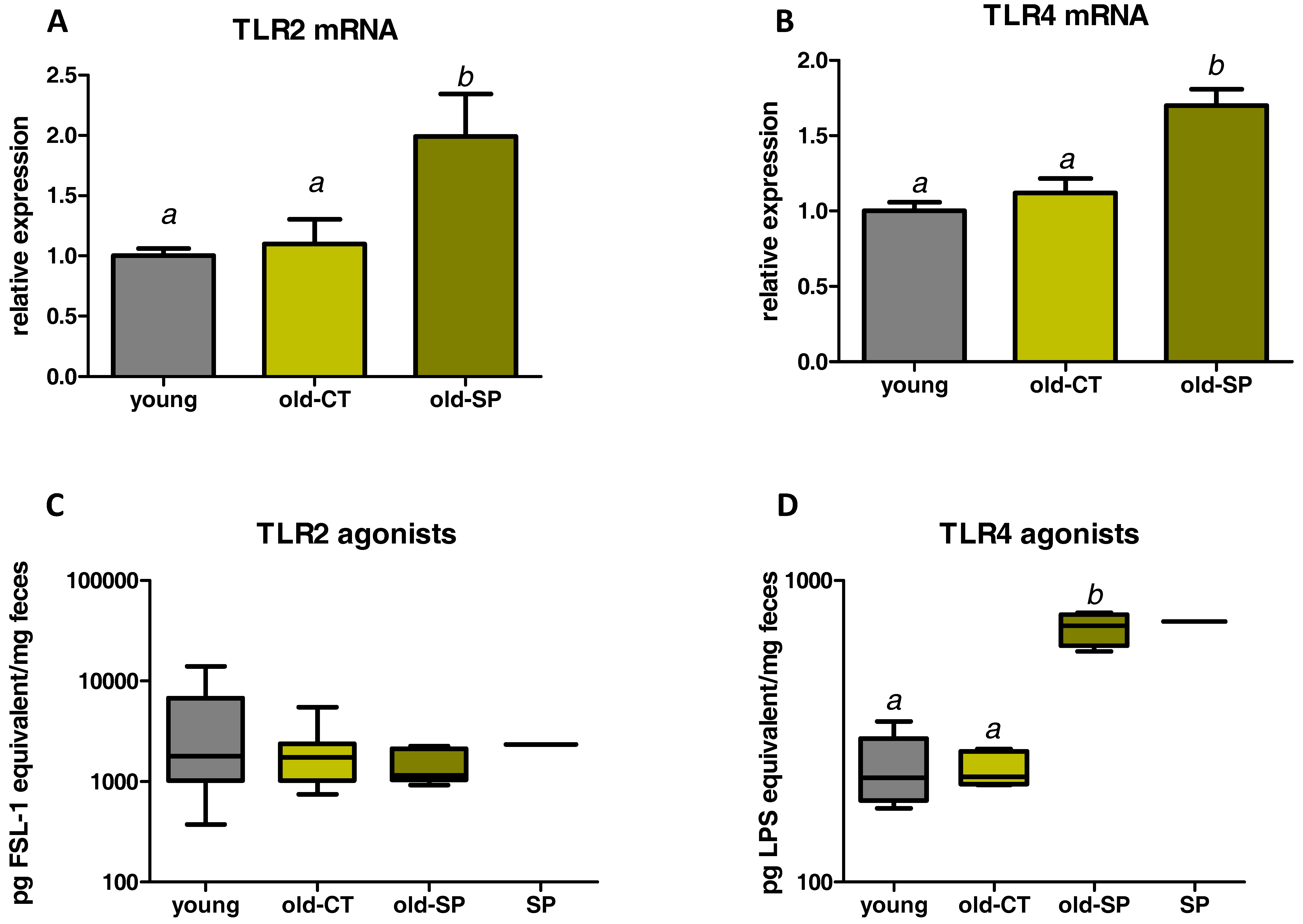
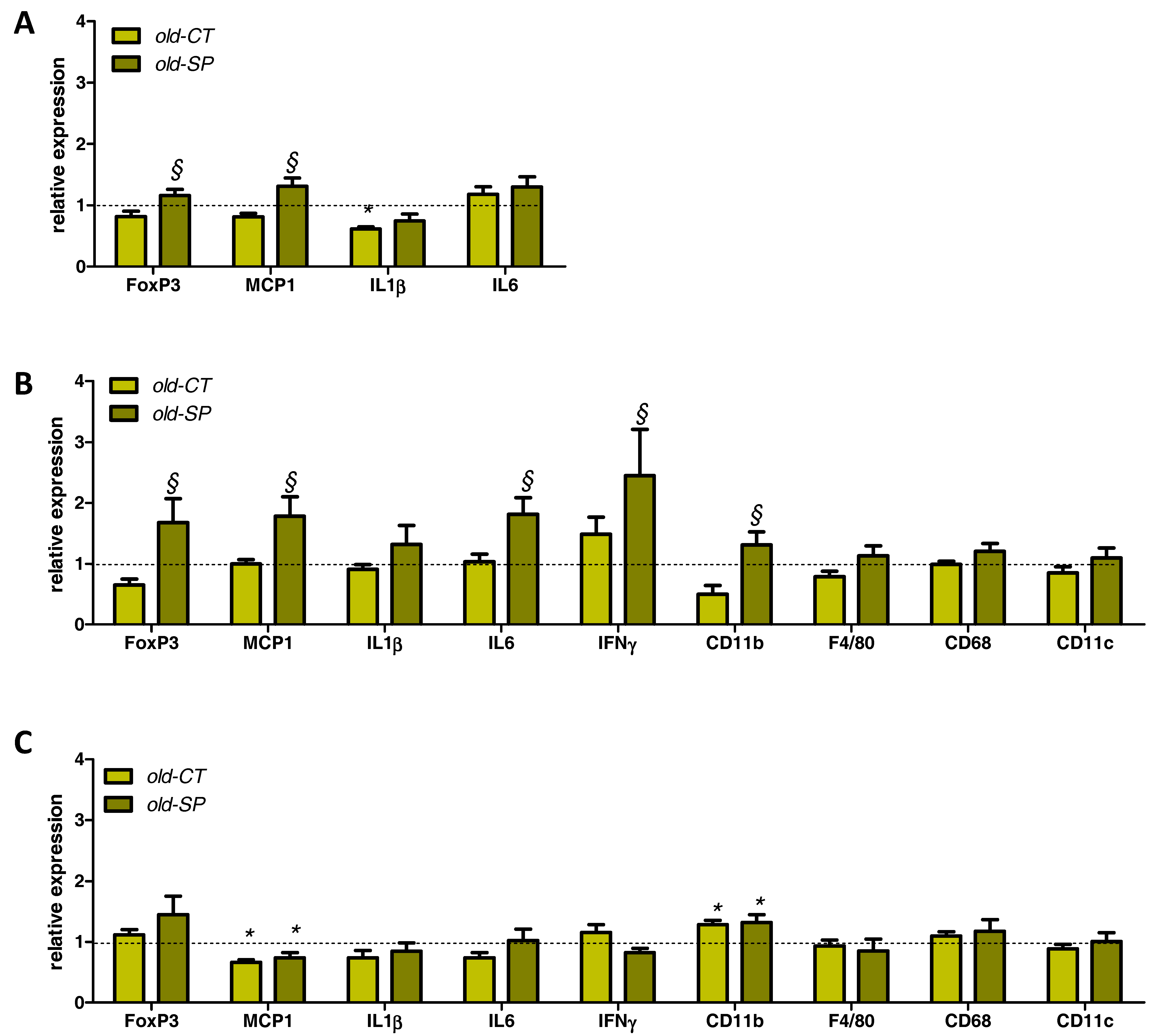
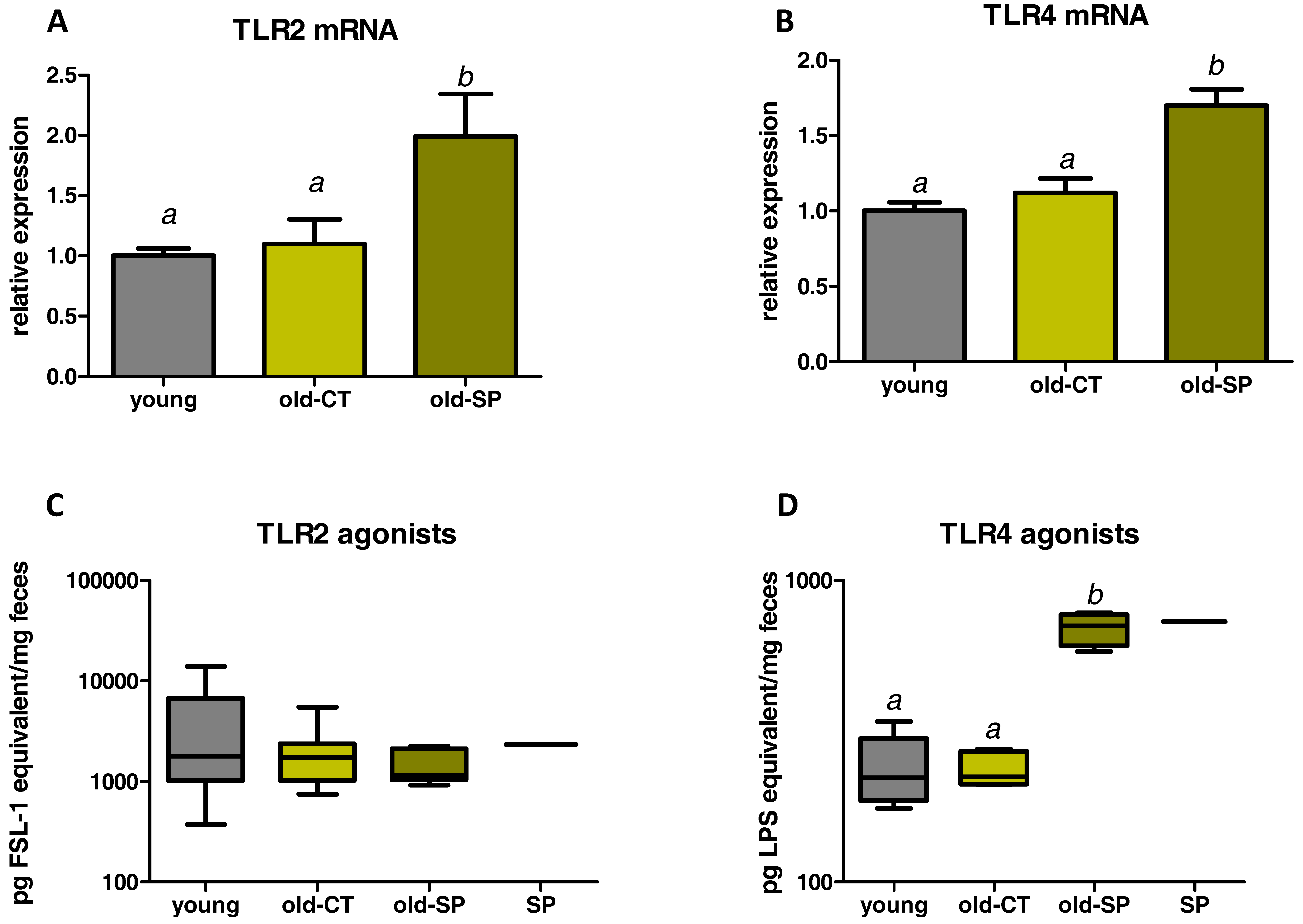
3.4. Spirulina Upregulated Parameters Related to the Innate Immunity in the Small Intestine of Old Mice
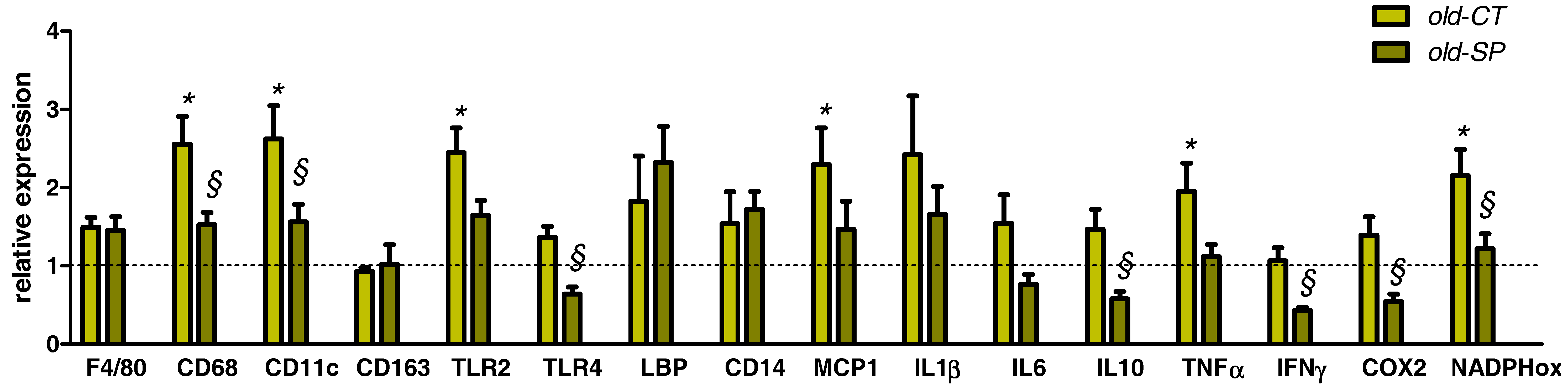
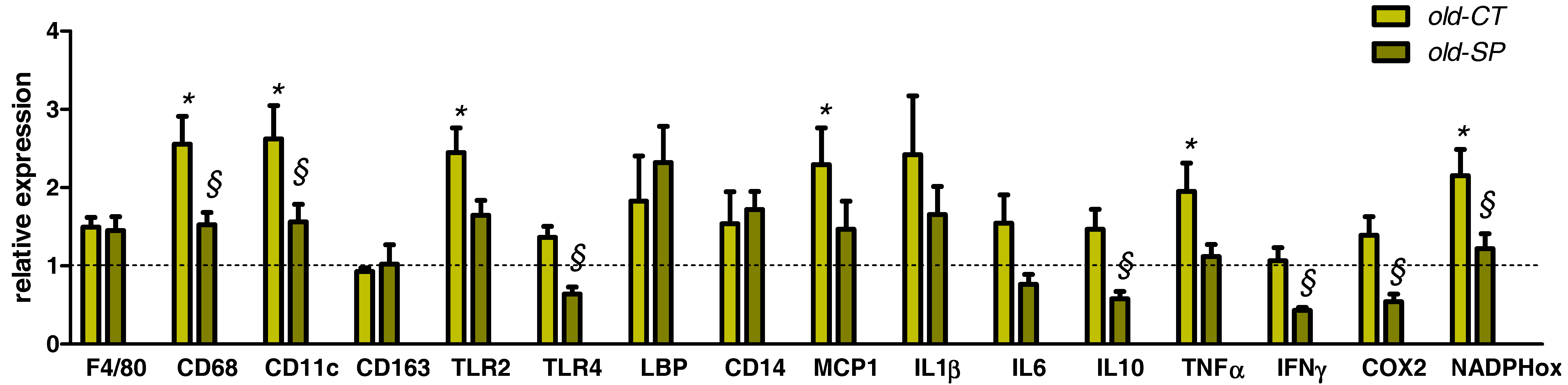
3.5. Spirulina Reduced Several Hepatic Inflammatory and Oxidative Stress Markers in Old Mice without Affecting Lipid Content
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nikoletopoulou, V.; Kyriakakis, E.; Tavernarakis, N. Cellular and molecular longevity pathways: The old and the new. Trends Endocrinol. Metab. 2014, 25, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Sheedfar, F.; Di Biase, S.; Koonen, D.; Vinciguerra, M. Liver diseases and aging: Friends or foes? Aging Cell 2013, 12, 950–954. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Zhao, E.; Amir, M.; Dong, H.; Tanaka, K.; Czaja, M.J. Aging promotes the development of diet-induced murine steatohepatitis but not steatosis. Hepatology 2013, 57, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Lyons, C.L.; Kennedy, E.B.; Roche, H.M. Metabolic inflammation-differential modulation by dietary constituents. Nutrients 2016, 8, 247. [Google Scholar] [CrossRef] [PubMed]
- Deng, R.; Chow, T.J. Hypolipidemic, antioxidant, and antiinflammatory activities of microalgae spirulina. Cardiovasc. Ther. 2010, 28, e33–e45. [Google Scholar] [CrossRef] [PubMed]
- Serban, M.C.; Sahebkar, A.; Dragan, S.; Stoichescu-Hogea, G.; Ursoniu, S.; Andrica, F.; Banach, M. A systematic review and meta-analysis of the impact of spirulina supplementation on plasma lipid concentrations. Clin. Nutr. 2016, 35, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.S.; Yang, Y.; Park, Y.; Lee, J. Health benefits of blue-green algae: Prevention of cardiovascular disease and nonalcoholic fatty liver disease. J. Med. Food 2013, 16, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Biagi, E.; Nylund, L.; Candela, M.; Ostan, R.; Bucci, L.; Pini, E.; Nikkila, J.; Monti, D.; Satokari, R.; Franceschi, C.; et al. Through ageing, and beyond: Gut microbiota and inflammatory status in seniors and centenarians. PLoS ONE 2010, 5, e10667. [Google Scholar] [CrossRef]
- Neyrinck, A.M.; Etxeberria, U.; Taminiau, B.; Daube, G.; van Hul, M.; Everard, A.; Cani, P.D.; Bindels, L.B.; Delzenne, N.M. Rhubarb extract prevents hepatic inflammation induced by acute alcohol intake, an effect related to the modulation of the gut microbiota. Mol. Nutr. Food Res. 2016, 61. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Possemiers, S.; Verstraete, W.; de Backer, F.; Cani, P.D.; Delzenne, N.M. Dietary modulation of clostridial cluster xiva gut bacteria (Roseburia spp.) by chitin-glucan fiber improves host metabolic alterations induced by high-fat diet in mice. J. Nutr. Biochem. 2012, 23, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Mukherjee, S.; Hooper, L.V. Antimicrobial defense of the intestine. Immunity 2015, 42, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Lazarevic, V.; Gaia, N.; Johansson, M.; Stahlman, M.; Backhed, F.; Delzenne, N.M.; Schrenzel, J.; Francois, P.; Cani, P.D. Microbiome of prebiotic-treated mice reveals novel targets involved in host response during obesity. ISME J. 2014, 8, 2116–2130. [Google Scholar] [CrossRef] [PubMed]
- Claesson, M.J.; Jeffery, I.B.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusack, S.; Harris, H.M.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Sze, M.A.; Schloss, P.D. Looking for a signal in the noise: Revisiting obesity and the microbiome. MBio 2016, 7, e01016–e01018. [Google Scholar] [CrossRef] [PubMed]
- Biagi, E.; Candela, M.; Fairweather-Tait, S.; Franceschi, C.; Brigidi, P. Aging of the human metaorganism: The microbial counterpart. Age 2012, 34, 247–267. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Fei, N.; Pang, X.; Shen, J.; Wang, L.; Zhang, B.; Zhang, M.; Zhang, X.; Zhang, C.; Li, M.; et al. A gut microbiota-targeted dietary intervention for amelioration of chronic inflammation underlying metabolic syndrome. FEMS Microbiol. Ecol. 2014, 87, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Schüppel, V.L.; Lockett, T.; Haller, D.; Delzenne, N.M. Microbiome and metabolic disorders related to obesity: Which lessons to learn from experimental models? Trends Food Sci. Technol. 2016, 57, 256–264. [Google Scholar] [CrossRef]
- Nakamura, N.; Lin, H.C.; McSweeney, C.S.; Mackie, R.I.; Gaskins, H.R. Mechanisms of microbial hydrogen disposal in the human colon and implications for health and disease. Annu. Rev. Food Sci. Technol. 2010, 1, 363–395. [Google Scholar] [CrossRef] [PubMed]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in il10-/- mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Barlow, G.M.; Yu, A.; Mathur, R. Role of the gut microbiome in obesity and diabetes mellitus. Nutr. Clin. Pract. 2015, 30, 787–797. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Schneeberger, M.; Everard, A.; Gomez-Valades, A.G.; Matamoros, S.; Ramirez, S.; Delzenne, N.M.; Gomis, R.; Claret, M.; Cani, P.D. Akkermansia muciniphila inversely correlates with the onset of inflammation, altered adipose tissue metabolism and metabolic disorders during obesity in mice. Sci. Rep. 2015, 5, 16643. [Google Scholar] [CrossRef] [PubMed]
- Usharani, G.; Srinivasan, S.; Sivasakthi, S.; Saranraj, P. Antimicrobial activity of spirulina platensis solvent s against pathogenic bacteria and fungi. Adv. Biol. Res. 2015, 9, 292–298. [Google Scholar]
- Nuhu, A.A. An important source of nutritional and medicinal compounds. J. Mar. Biol. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Balachandran, P.; Pugh, N.D.; Ma, G.; Pasco, D.S. Toll-like receptor 2-dependent activation of monocytes by spirulina polysaccharide and its immune enhancing action in mice. Int. Immunopharmacol. 2006, 6, 1808–1814. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Ohnishi, H. Role of gut microbiota and toll-like receptors in nonalcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 7381–7391. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Hashiguchi, M.; Toda, E.; Iwasaki, A.; Hachimura, S.; Kaminogawa, S. Cd11b+ peyer’s patch dendritic cells secrete IL-6 and induce IgA secretion from naive B cells. J. Immunol. 2003, 171, 3684–3690. [Google Scholar] [CrossRef] [PubMed]
- Santiago, A.F.; Alves, A.C.; Oliveira, R.P.; Fernandes, R.M.; Paula-Silva, J.; Assis, F.A.; Carvalho, C.R.; Weiner, H.L.; Faria, A.M. Aging correlates with reduction in regulatory-type cytokines and t cells in the gut mucosa. Immunobiology 2011, 216, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Beutler, B. Microbe sensing, positive feedback loops, and the pathogenesis of inflammatory diseases. Immunol. Rev. 2009, 227, 248–263. [Google Scholar] [CrossRef] [PubMed]
- Pak, W.; Takayama, F.; Mine, M.; Nakamoto, K.; Kodo, Y.; Mankura, M.; Egashira, T.; Kawasaki, H.; Mori, A. Anti-oxidative and anti-inflammatory effects of spirulina on rat model of non-alcoholic steatohepatitis. J. Clin. Biochem. Nutr. 2012, 51, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Remirez, D.; Fernandez, V.; Tapia, G.; Gonzalez, R.; Videla, L.A. Influence of C-phycocyanin on hepatocellular parameters related to liver oxidative stress and kupffer cell functioning. Inflamm. Res. 2002, 51, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Romay, C.; Delgado, R.; Remirez, D.; Gonzalez, R.; Rojas, A. Effects of phycocyanin extract on tumor necrosis factor-alpha and nitrite levels in serum of mice treated with endotoxin. Arzneimittelforschung 2001, 51, 733–736. [Google Scholar] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Young | Old-CT | Old-SP | |
|---|---|---|---|
| Body weight gain (g) | 3.11 ± 0.38 a | 0.23 ± 0.29 b | 0.64 ± 0.65 b |
| Total food intake (g) | 108 ± 7 | 130 ± 17 | 121 ± 2 |
| Visceral adipose tissue (g) | 0.24 ± 0.03 | 0.25 ± 0.03 | 0.19 ± 0.04 |
| Subcutaneous adipose tissue (g) | 0.42 ± 0.04 | 0.32 ± 0.04 | 0.30 ± 0.05 |
| Epididymal adipose tissue (g) | 0.51 ± 0.05 | 0.37 ± 0.05 | 0.33 ± 0.08 |
| Liver (g/100 g body weight) | 3.71 ± 0.09 a | 4.25 ± 0.24 b | 4.09 ± 0.10 a,b |
| Spleen (g/100 g body weight) | 0.30 ± 0.01 | 0.41± 0.06 | 0.35 ± 0.06 |
| Caecal tissue (g/100 g body weight) | 0.64 ± 0.06 | 0.62 ± 0.03 | 0.61 ± 0.06 |
| Caecal content (g) | 0.17 ± 0.02 | 0.19 ± 0.01 | 0.19 ± 0.02 |
| Young | Old-CT | Old-SP | |
|---|---|---|---|
| Triglyceride content (nmol/mg tissue) | 15.1 ± 1.7 | 10.4 ± 0.7 | 13.8 ± 2.2 |
| Cholesterol content (nmol/mg tissue) | 7.4 ± 0.6 a | 9.3 ± 0.3 a,b | 10.8 ± 1.3 b |
| ALAT (U/L) | 4.1 ± 1.2 | 3.1 ± 0.3 | 3.8 ± 0.8 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neyrinck, A.M.; Taminiau, B.; Walgrave, H.; Daube, G.; Cani, P.D.; Bindels, L.B.; Delzenne, N.M. Spirulina Protects against Hepatic Inflammation in Aging: An Effect Related to the Modulation of the Gut Microbiota? Nutrients 2017, 9, 633. https://doi.org/10.3390/nu9060633
Neyrinck AM, Taminiau B, Walgrave H, Daube G, Cani PD, Bindels LB, Delzenne NM. Spirulina Protects against Hepatic Inflammation in Aging: An Effect Related to the Modulation of the Gut Microbiota? Nutrients. 2017; 9(6):633. https://doi.org/10.3390/nu9060633
Chicago/Turabian StyleNeyrinck, Audrey M., Bernard Taminiau, Hannah Walgrave, Georges Daube, Patrice D. Cani, Laure B. Bindels, and Nathalie M. Delzenne. 2017. "Spirulina Protects against Hepatic Inflammation in Aging: An Effect Related to the Modulation of the Gut Microbiota?" Nutrients 9, no. 6: 633. https://doi.org/10.3390/nu9060633
APA StyleNeyrinck, A. M., Taminiau, B., Walgrave, H., Daube, G., Cani, P. D., Bindels, L. B., & Delzenne, N. M. (2017). Spirulina Protects against Hepatic Inflammation in Aging: An Effect Related to the Modulation of the Gut Microbiota? Nutrients, 9(6), 633. https://doi.org/10.3390/nu9060633