Dietary Management of Skin Health: The Role of Genistein



Abstract
:1. Introduction
2. In Vitro Evidence
2.1. Genistein Effects on Fibroblasts
2.2. Genistein Affects Glycosaminoglycan Synthesis
2.3. Genistein Effects on ultraviolet (UV)-Protection
3. In Vivo Studies
3.1. Genistein Effect on Photodamage
3.2. Genistein Effect on Wound Healing
4. Human Studies
5. The Problem of Delivery
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Hall, G.; Phillips, T.J. Estrogen and skin: The effects of estrogen, menopause, and hormone replacement therapy on the skin. J. Am. Acad. Dermatol. 2005, 53, 555–568. [Google Scholar] [CrossRef]
- Verdier-Sevrain, S.; Bontè, F.; Gilchrest, B. Biology of estrogens in skin: Implications for skin aging. Exp. Dermatol. 2006, 15, 83–94. [Google Scholar] [CrossRef]
- Wolff, E.F.; Narayan, D.; Taylor, H.S. Long-term effects of hormone therapy on skin rigidity and wrinkles. Fertil. Steril. 2005, 84, 285–288. [Google Scholar] [CrossRef]
- Albertazzi, P.; Purdie, D.W. The life and times of the estrogen receptors: An interim report. Climacteric 2001, 4, 194–202. [Google Scholar] [CrossRef]
- Younes, M.; Honma, N. Estrogen receptor β. Arch. Pathol. Lab. Med. 2011, 135, 63–66. [Google Scholar]
- Thornton, M.J.; Taylor, A.H.; Mulligan, K.; Al-Azzawi, F.; Lyon, C.C.; O’Driscoll, J.B.; Messenger, A.G. Estrogen receptor beta (ERβ) is the predominant estrogen receptor in human scalp. Exp. Dermatol. 2003, 12, 181–190. [Google Scholar] [CrossRef]
- Thornton, M.J.; Taylor, A.H.; Mulligan, K.; Al-Azzawi, F.; Lyon, C.C.; O’Driscoll, J.B.; Messenger, A.G. The distribution of estrogen receptor beta (ERβ) is distinct to that of ER alpha and the androgen receptor in human skin and the pilosebaceous unit. J. Investig. Dermatol. Symp. Proc. 2003, 8, 100–103. [Google Scholar] [CrossRef]
- Thornton, M.J. The biological actions of estrogens on skin. Exp. Dermatol. 2002, 11, 487–502. [Google Scholar] [CrossRef]
- Fitzpatrick, L.A. Selective estrogen receptor modulators and phytoestrogens: New therapies for the postmenopausal women. Mayo Clin. Proc. 1999, 74, 601–607. [Google Scholar] [CrossRef]
- Beck, V.; Unterrieder, E.; Krenn, L.; Kubelka, W.; Jungbauer, A. Comparison of hormonal activity (estrogen, androgen and progestin) of standardized plant extracts for large scale use in hormone replacement therapy. J. Steroid Biochem. Mol. Biol. 2003, 84, 259–268. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Lemmen, J.G.; Carlsson, B.; Corton, J.C.; Safe, S.H.; van der Saag, P.T.; van der Burg, B.; Gustafsson, J.A. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta. Endocrinology 1998, 139, 4252–4263. [Google Scholar] [CrossRef]
- Akiyama, T.; Ishida, J.; Nakagawa, S.; Ogawara, H.; Watanabe, S.; Itoh, N.; Shibuya, M.; Fukami, Y. Genistein, a specific inhibitor of tyrosine-specific protein kinases. J. Biol. Chem. 1987, 262, 5592–5595. [Google Scholar]
- Steensma, A.; Faassen-Peters, M.A.; Noteborn, H.P.; Rietjens, I.M. Bioavailability of genistein and its glycoside genistin as measured in the portal vein of freely moving unanesthetized rats. J. Agric. Food Chem. 2006, 54, 8006–8012. [Google Scholar] [CrossRef]
- Cederroth, C.R.; Zimmermann, C.; Nef, S. Soy, phytoestrogens and their impact on reproductive health. Mol. Cell. Endocrinol. 2012, 355, 192–200. [Google Scholar] [CrossRef]
- Chin, G.S.; Liu, W.; Steinbrech, D.; Hsu, M.; Levinson, H.; Longaker, M.T. Cellular signaling by tyrosine phosphorylation in keloid and normal human dermal fibroblasts. Plast. Reconstr. Surg. 2000, 106, 1532–1540. [Google Scholar] [CrossRef]
- Cao, C.; Li, S.; Dai, X.; Chen, Y.; Feng, Z.; Zhao, Y.; Wu, J. Genistein inhibits proliferation and functions of hypertrophic scar fibroblasts. Burns 2009, 35, 89–97. [Google Scholar] [CrossRef]
- Jurzak, M.; Adamczyk, K.; Antończak, P.; Garncarczyk, A.; Kuśmierz, D.; Latocha, M. Evaluation of genistein ability to modulate CTGF mRNA/protein expression, genes expression of TGFβ isoforms and expression of selected genes regulating cell cycle in keloid fibroblasts in vitro. Acta Pol. Pharm. 2014, 71, 972–986. [Google Scholar]
- Jurzak, M.; Adamczyk, K. Influence of genistein on c-Jun, c-Fos and Fos-B of AP-1 subunits expression in skin keratinocytes, fibroblasts and keloid fibroblasts cultured in vitro. Acta Pol. Pharm. 2013, 70, 205–213. [Google Scholar]
- Sienkiewicz, P.; Surazyński, A.; Pałka, J.; Miltyk, W. Nutritional concentration of genistein protects human dermal fibroblasts from oxidative stress-induced collagen biosynthesis inhibition through IGF-I receptor-mediated signaling. Acta Pol. Pharm. 2008, 65, 203–211. [Google Scholar]
- Piotrowska, E.; Jakóbkiewicz-Banecka, J.; Barańska, S.; Tylki-Szymańska, A.; Czartoryska, B.; Wegrzyn, A.; Wegrzyn, G. Genistein-mediated inhibition of glycosaminoglycan synthesis as a basis for gene expression-targeted isoflavone therapy for mucopolysaccharidoses. Eur. J. Hum. Genet. 2006, 14, 846–852. [Google Scholar] [CrossRef]
- Otomo, T.; Hossain, M.A.; Ozono, K.; Sakai, N. Genistein reduces heparan sulfate accumulation in human mucolipidosis II skin fibroblasts. Mol. Genet. Metab. 2012, 105, 266–269. [Google Scholar] [CrossRef]
- Kloska, A.; Jakóbkiewicz-Banecka, J.; Narajczyk, M.; Banecka-Majkutewicz, Z.; Węgrzyn, G. Effects of flavonoids on glycosaminoglycan synthesis: Implications for substrate reduction therapy in Sanfilippo disease and other mucopolysaccharidoses. Metab. Brain Dis. 2011, 26, 1–8. [Google Scholar] [CrossRef]
- Isoherranen, K.; Punnonen, K.; Jansen, C.; Uotila, P. Ultraviolet irradiation induces cyclooxygenase-2 expression in keratinocytes. Br. J. Dermatol. 1999, 140, 1017–1022. [Google Scholar] [CrossRef]
- Iovine, B.; Iannella, M.L.; Gasparri, F.; Monfrecola, G.; Bevilacqua, M.A. Synergic Effect of Genistein and Daidzein on UVB-Induced DNA Damage: An Effective Photoprotective Combination. J. Biomed. Biotechnol. 2011, 2011, 692846:1–692846:8. [Google Scholar] [CrossRef]
- Wang, Y.N.; Wu, W.; Chen, H.C.; Fang, H. Genistein protects against UVB-induced senescence-like characteristics in human dermal fibroblast by p66Shc down-regulation. J. Dermatol. Sci. 2010, 58, 19–27. [Google Scholar] [CrossRef]
- Lee, T.H.; Do, M.H.; Oh, Y.L.; Cho, D.W.; Kim, S.H.; Kim, S.Y. Dietary fermented soybean suppresses UVB-induced skin inflammation in hairless mice via regulation of the MAPK signaling pathway. J. Agric. Food Chem. 2014, 62, 8962–8972. [Google Scholar] [CrossRef]
- Wei, H.; Bowen, R.; Zhang, X.; Lebwohl, M. Isoflavone genistein inhibits the initiation and promotion of two-stage skin carcinogenesis in mice. Carcinogenesis 1998, 19, 1509–1514. [Google Scholar] [CrossRef]
- Khan, N.; Afaq, F.; Mukhtar, H. Cancer chemoprevention through dietary antioxidants: Progress and promise. Antioxid. Redox Signal. 2008, 10, 475–510. [Google Scholar] [CrossRef]
- Terra, V.A.; Souza-Neto, F.P.; Frade, M.A.; Ramalho, L.N.; Andrade, T.A.; Pasta, A.A.; Conchon, A.C.; Guedes, F.A.; Luiz, R.C.; Cecchini, R.; et al. Genistein prevents ultraviolet B radiation-induced nitrosative skin injury and promotes cell proliferation. J. Photochem. Photobiol. B 2015, 144, 20–27. [Google Scholar] [CrossRef]
- Wei, H.; Saladi, R.; Lu, Y.; Wang, Y.; Palep, S.R.; Moore, J.; Phelps, R.; Shyong, E.; Lebwohl, M.G. Isoflavone genistein: Photoprotection and clinical implications in dermatology. J. Nutr. 2003, 133 (Suppl. 1), 3811S–3819S. [Google Scholar]
- Park, E.; Lee, S.M.; Jung, I.K.; Lim, Y.; Kim, J.H. Effects of genistein on early-stage cutaneous wound healing. Biochem. Biophys. Res. Commun. 2011, 410, 514–519. [Google Scholar] [CrossRef]
- Emmerson, E.; Campbell, L.; Ashcroft, G.S.; Hardman, M.J. The phytoestrogen genistein promotes wound healing by multiple independent mechanisms. Mol. Cell. Endocrinol. 2010, 321, 184–193. [Google Scholar] [CrossRef]
- Marini, H.; Polito, F.; Altavilla, D.; Irrera, N.; Minutoli, L.; Calò, M.; Adamo, E.B.; Vaccaro, M.; Squadrito, F.; Bitto, A. Genistein aglycone improves skin repair in an incisional model of wound healing: A comparison with raloxifene and oestradiol in ovariectomized rats. Br. J. Pharmacol. 2010, 160, 1185–1194. [Google Scholar] [CrossRef]
- Polito, F.; Marini, H.; Bitto, A.; Irrera, N.; Vaccaro, M.; Adamo, E.B.; Micali, A.; Squadrito, F.; Minutoli, L.; Altavilla, D. Genistein aglycone, a soy-derived isoflavone, improves skin changes induced by ovariectomy in rats. Br. J. Pharmacol. 2012, 165, 994–1005. [Google Scholar] [CrossRef]
- Kessel, B. Alternatives to estrogen for menopausal women. Soc. Exp. Biol. Med. 1998, 217, 38–44. [Google Scholar] [CrossRef]
- Izumi, T.; Saito, M.; Obata, A.; Arii, M.; Yamaguchi, H.; Matsuyama, A. Oral intake of soy isoflavone aglycone improves the aged skin of adult women. J. Nutr. Sci. Vitaminol. (Tokyo) 2007, 53, 57–62. [Google Scholar] [CrossRef]
- Moraes, A.B.; Haidar, M.A.; Soares Júnior, J.M.; Simões, M.J.; Baracat, E.C.; Patriarca, M.T. The effects of topical isoflavones on postmenopausal skin: Double-blind and randomized clinical trial of efficacy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2009, 146, 188–192. [Google Scholar] [CrossRef]
- Delgadillo, V.; O’Callaghan Mdel, M.; Artuch, R.; Montero, R.; Pineda, M. Genistein supplementation in patients affected by Sanfilippo disease. J. Inherit. Metab. Dis. 2011, 34, 1039–1044. [Google Scholar] [CrossRef]
- Setchell, K.D.; Brown, N.M.; Desai, P.; Zimmer-Nechemias, L.; Wolfe, B.E.; Brashear, W.T.; Kirschner, A.S.; Cassidy, A.; Heubi, J.E. Bioavailability of pure isoflavones in healthy humans and analysis of commercial soy isoflavone supplements. J. Nutr. 2001, 131, 1362S–1375S. [Google Scholar]
- Setchell, K.D.; Brown, N.M.; Desai, P.B.; Zimmer-Nechimias, L.; Wolfe, B.; Jakate, A.S.; Creutzinger, V.; Heubi, J.E. Bioavailability, disposition, and dose-response effects of soy isoflavones when consumed by healthy women at physiologically typical dietary intakes. J. Nutr. 2003, 133, 1027–1035. [Google Scholar]
- Zubik, L.; Meydani, M. Bioavailability of soybean isoflavones from aglycone and glucoside forms in American women. Am. J. Clin. Nutr. 2003, 77, 1459–1465. [Google Scholar]
- Georgetti, S.R.; Casagrande, R.; Verri, W.A., Jr.; Lopez, R.F.; Fonseca, M.J. Evaluation of in vivo efficacy of topical formulations containing soybean extract. Int. J. Pharm. 2008, 352, 189–196. [Google Scholar] [CrossRef]
- Müller, R.H.; Petersen, R.D.; Hommoss, A.; Pardeike, J. Nanostructured lipid carriers (NLC) in cosmetic dermal products. Adv. Drug Deliv. Rev. 2007, 59, 522–530. [Google Scholar] [CrossRef]
- Andrade, L.M.; de Fátima Reis, C.; Maione-Silva, L.; Anjos, J.L.; Alonso, A.; Serpa, R.C.; Marreto, R.N.; Lima, E.M.; Taveira, S.F. Impact of lipid dynamic behavior on physical stability, in vitro release and skin permeation of genistein-loaded lipid nanoparticles. Eur. J. Pharm. Biopharm. 2014, 88, 40–47. [Google Scholar] [CrossRef]
- Minghetti, P.; Cilurzo, F.; Casiraghi, A.; Montanari, L. Evaluation of ex vivo human skin permeation of genistein and daidzein. Drug Deliv. 2006, 13, 411–415. [Google Scholar] [CrossRef]
- Huang, Z.R.; Hung, C.F.; Lin, Y.K.; Fang, J.Y. In vitro and in vivo evaluation of topical delivery and potential dermal use of soy isoflavones genistein and daidzein. Int. J. Pharm. 2008, 364, 36–44. [Google Scholar] [CrossRef]
- Chadha, G.; Sathigari, S.; Parsons, D.L.; Jayachandra Babu, R. In vitro percutaneous absorption of genistein from topical gels through human skin. Drug Dev. Ind. Pharm. 2011, 37, 498–505. [Google Scholar] [CrossRef]
- Kitagawa, S.; Inoue, K.; Teraoka, R.; Morita, S.Y. Enhanced skin delivery of genistein and other two isoflavones by microemulsion and prevention against UV irradiation-induced erythema formation. Chem. Pharm. Bull. (Tokyo) 2010, 58, 398–401. [Google Scholar] [CrossRef]
- Arcoraci, V.; Atteritano, M.; Squadrito, F.; D’Anna, R.; Marini, H.; Santoro, D.; Minutoli, L.; Messina, S.; Altavilla, D.; Bitto, A. Antiosteoporotic Activity of Genistein Aglycone in Postmenopausal Women: Evidence from a Post-Hoc Analysis of a Multicenter Randomized Controlled Trial. Nutrients 2017, 9, 179. [Google Scholar] [CrossRef]
- Bitto, A.; Polito, F.; Squadrito, F.; Marini, H.; D’Anna, R.; Irrera, N.; Minutoli, L.; Granese, R.; Altavilla, D. Genistein aglycone: A dual mode of action anti-osteoporotic soy isoflavone rebalancing bone turnover towards bone formation. Curr. Med. Chem. 2010, 17, 3007–3018. [Google Scholar] [CrossRef]
- Marini, H.; Bitto, A.; Altavilla, D.; Burnett, B.P.; Polito, F.; Di Stefano, V.; Minutoli, L.; Atteritano, M.; Levy, R.M.; D’Anna, R.; et al. Breast safety and efficacy of genistein aglycone for postmenopausal bone loss: A follow-up study. J. Clin. Endocrinol. Metab. 2008, 93, 4787–4796. [Google Scholar] [CrossRef] [PubMed]
- Marini, H.; Minutoli, L.; Polito, F.; Bitto, A.; Altavilla, D.; Atteritano, M.; Gaudio, A.; Mazzaferro, S.; Frisina, A.; Frisina, N.; et al. Effects of the phytoestrogen genistein on bone metabolism in osteopenic postmenopausal women: A randomized trial. Ann. Intern. Med. 2007, 146, 839–847. [Google Scholar] [CrossRef] [PubMed]
- D’Anna, R.; Cannata, M.L.; Marini, H.; Atteritano, M.; Cancellieri, F.; Corrado, F.; Triolo, O.; Rizzo, P.; Russo, S.; Gaudio, A.; et al. Effects of the phytoestrogen genistein on hot flushes, endometrium, and vaginal epithelium in postmenopausal women: A 2-year randomized, double-blind, placebo-controlled study. Menopause 2009, 16, 301–306. [Google Scholar] [CrossRef] [PubMed]
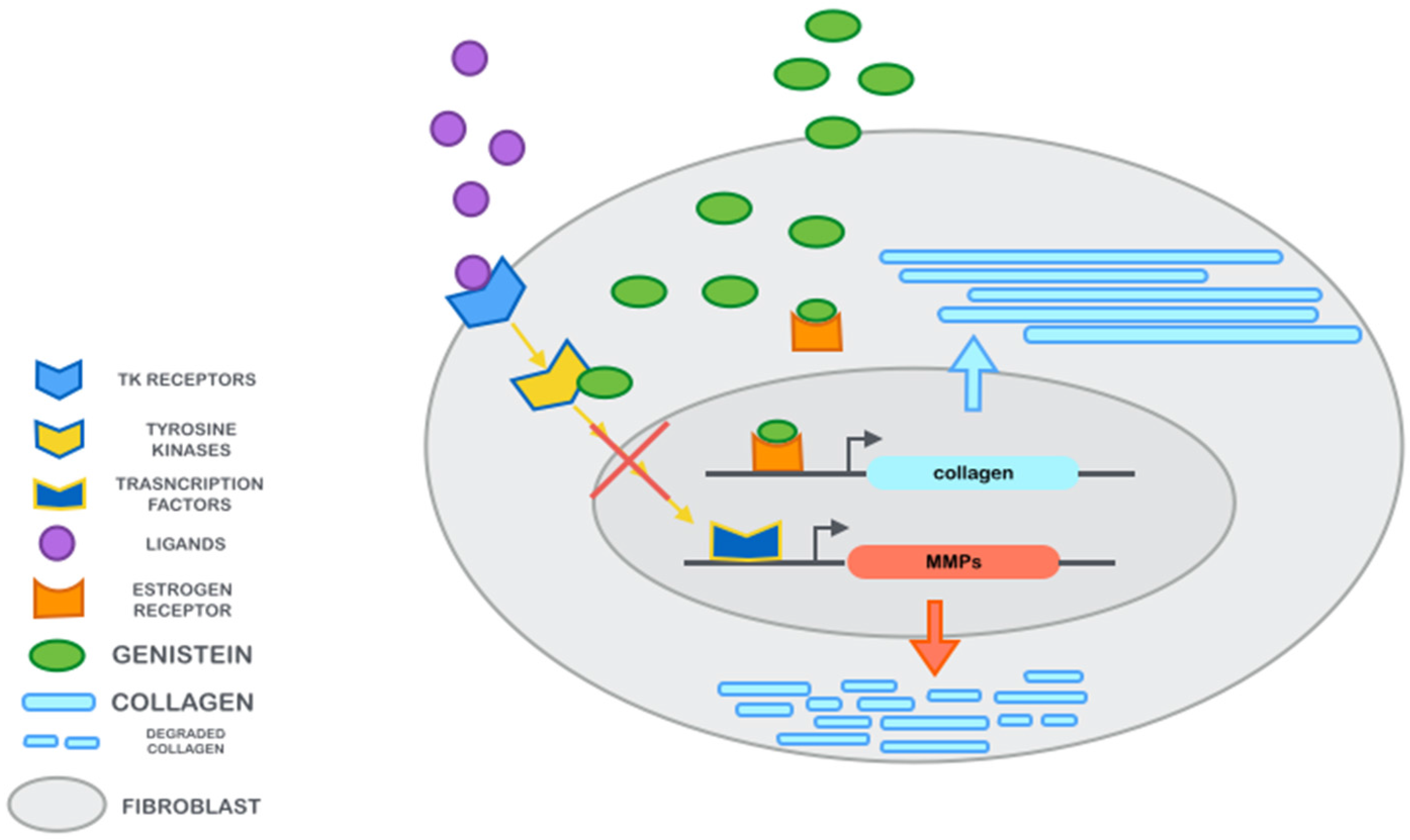

{kind=link}
| In Vitro Experimental Models | Genistein Effects |
|---|---|
| Fibroblasts from hypertrophic burn scars, normal human dermal fibroblasts (NHDF) keloid fibroblasts (KEL FIB), normal human epidermal keratinocytes (NHEK), NHDF stimulated with t-BHP (t-butylhydroperoxide) [16,17,18,19] | ↓ RAS, RAF, ERK, p38 |
| ↓ CTGF, TGFβ1, β2 and β3 | |
| ↑ BCL-2 | |
| ↑ C-JUN | |
| Fibroblasts from patients with mucolipidosis II and Sanfilippo disease [21,22] | ↓ GAG |
| HaCaT, BJ-5ta, and human dermal fibroblasts exposed to UV radiation [23,24,25] | ↓ COX-2 |
| ↑ Gadd45 gene expression | |
| ↓ p66Shc protein |
| In Vivo Model | Dose | Outcomes | Genistein Effects |
|---|---|---|---|
| UVB-irradiation and carcinogenesis models [26,27,29] | Dietary supplementation 0.2083 μg/mg of fermented soy beans 1, 5, 10, 20 μmol genistein topically applied Intraperitoneal injections 10–15 mg/kg | Anti-inflammatory effect in hairless mice with photodamage Reduction of skin tumorigenesis in mice Anti-nitrosative effect in hairless mice | ↓ skin inflammation |
| ↓ H2O2 | |||
| ↓ 8-hydroxy-2-deoxyguanosine | |||
| ↓ lipid peroxides | |||
| ↓ nitrotyrosine | |||
| ↓ tyrosine protein kinase | |||
| ↓ EGF-receptor phosphorylation | |||
| ↓ MAPK activation | |||
| ↓ oncoprotein expression | |||
| ↑ PCNA | |||
| ↑ Ki67 | |||
| Wound healing models [31,32,33,34] | Dietary supplementation 0.25–1 g/kg Subcutaneous injections 50 mg/kg Subcutaneous injections 1–10 mg/kg Subcutaneous injections 1–10 mg/kg | Improving wound healing in intact mice Wound healing in OVX mice Wound healing in OVX rats Skin aging in OVX rats | ↑ wound repair |
| ↓ ROS | |||
| ↓ NF-κB | |||
| ↓ TNF-α | |||
| ↑ TGF-β1 | |||
| ↑ VEGF | |||
| ↑ MMP-2 and MMP-9 | |||
| ↑ TIMP-1 and TIMP-2 | |||
| ↑ collagen thickness | |||
| ↑ skin breaking strength |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irrera, N.; Pizzino, G.; D’Anna, R.; Vaccaro, M.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Dietary Management of Skin Health: The Role of Genistein. Nutrients 2017, 9, 622. https://doi.org/10.3390/nu9060622
Irrera N, Pizzino G, D’Anna R, Vaccaro M, Arcoraci V, Squadrito F, Altavilla D, Bitto A. Dietary Management of Skin Health: The Role of Genistein. Nutrients. 2017; 9(6):622. https://doi.org/10.3390/nu9060622
Chicago/Turabian StyleIrrera, Natasha, Gabriele Pizzino, Rosario D’Anna, Mario Vaccaro, Vincenzo Arcoraci, Francesco Squadrito, Domenica Altavilla, and Alessandra Bitto. 2017. "Dietary Management of Skin Health: The Role of Genistein" Nutrients 9, no. 6: 622. https://doi.org/10.3390/nu9060622
APA StyleIrrera, N., Pizzino, G., D’Anna, R., Vaccaro, M., Arcoraci, V., Squadrito, F., Altavilla, D., & Bitto, A. (2017). Dietary Management of Skin Health: The Role of Genistein. Nutrients, 9(6), 622. https://doi.org/10.3390/nu9060622