Black‐Box Gastrointestinal Tract—Needs and Prospects of Gaining Insights of Fate of Fat, Protein, and Starch in Case of Exocrine Pancreatic Insufficiency by Using Fistulated Pigs


Abstract
:1. Background—Digestive Processes in the Gastrointestinal Tract
2. Use of Ileo-Cecal Fistulated Pigs in Research on Diverse Effects of Exocrine Pancreatic Insufficiency (EPI) on Digestive Processes and Optimization of Treatment and Dietetic Measures
3. Conclusions
Author Contributions
Conflicts of Interest
References
- McDonald, P.; Edwards, R.A.; Greenhalgh, J.F.D.; Morgan, C.A. Animal Nutrition, 6th ed.; Pearson Education Limited: Harlow, UK, 2002. [Google Scholar]
- Pond, W.G.; Church, D.C.; Pond, K.R.; Schoknecht, P.A. Basic Animal Nutrition and Feeding, 5th ed.; Wiley: Hoboken, NJ, USA, 2005. [Google Scholar]
- Laplace, J.P.; Souffrant, W.B.; Hennig, U.; Chabeauti, E.; Fevrier, C. Measurement of praecaecal dietary protein and plant cell wall digestion in pigs; Comparison of four surgical procedures for ileorectal anastomosis. Livest. Prod. Sci. 1994, 40, 313–328. [Google Scholar] [CrossRef]
- Sauer, W.C.; Fan, M.Z.; Mosenthin, R.; Drochner, W. Methods for measuring ileal amino acid digestibility in pigs. In Farm Animal Metabolism and Nutrition; D’Mello, J.P.F., Ed.; CABI Publ.: Wallingford, UK, 2000; pp. 279–306. [Google Scholar]
- Laplace, J.P.; Darcy-Vrillon, B.; Pe’rez, J.M.; Henry, Y.; Giger, S.; Sauvant, D. Associative effects between two fibre sources on ileal and overall digestibilities of amino acids, energy and cell-wall components in growing pigs. Br. J. Nutr. 1989, 61, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Wünsche, J.; Bock, H.D.; Henning, U.; Kreienbring, F.; Borgmann, E. Untersuchungen über die Proteinverdaulichkeit und Aminosäurenresorption in verschiedenen Abschnitten des Verdauungstraktes beim Schwein—2.: Die Protein-und Aminosäurenbilanz am Ende des Dünndarms und des gesamten Verdauungstraktes (scheinbare und wahre Protein-und Aminosäurenverdaulichkeit). Arch. Tierern. 1979, 29, 221–234. [Google Scholar]
- Schmitz, M.; Ahrens, F.; Schön, J.; Hagemeister, H. Amino acids absorption in the large intestine of cow, pig and horse and its significance for protein supply. Adv. Anim. Phys. Anim. Nutr. 1991, 22, 67–71. [Google Scholar]
- Mosenthin, R.; Rademacher, M.; Sauer, W.C. Zur scheinbaren präzäkalen Verdaulichkeit von Aminosäuren in Futtermitteln für Schweine. Übers Tierernarg. 1997, 25, 41–85. [Google Scholar]
- Just, A.; Jorgensen, H.; Fernandez, J.A. The digestive capacity of the caecum-colon and the value of the nitrogen absorbed from the hind gut for protein synthesis in pigs. Br. J. Nutr. 1981, 46, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Zebrowska, T.; Buraczewska, L.; Buraczewski, S. The apparent digestibility of amino acids in the small intestine and in the whole digestive tract of pigs fed diets containing different sources of protein. Rocz. Nauk. Roln. 1978, 99, B87–B98. [Google Scholar]
- Williams, P.E.V. Digestible amino acids for non-ruminant animals: Theory and recent challenges. Anim. Feed Sci. Technol. 1995, 53, 173–187. [Google Scholar] [CrossRef]
- GfE (Gesellschaft für Ernährungsphysiologie); Committee for Requirement Standards of the Society of Nutrition Physiology. Recommendations for the Supply of Energy and Nutrients to Pigs; DLG-Verlag: Frankfurt, Germany, 2008. [Google Scholar]
- Mosenthin, R.; Sauer, W.C. Zur Methodik der präzäkalen Verdaulichkeitsbestimmung beim Schwein. Übers Tierernarg. 1992, 20, 65–90. [Google Scholar]
- Flachowsky, G. Bestimmung der praecaecalen Verdaulichkeit von Aminosäuren beim Schwein- Empfehlungen zur standardisierten Versuchsdurchführung. Mitteilungen des Ausschusses für Bedarfsnormen der Gesellschaft für Ernährungsphysiologie. Proc. Soc. Nutr. Physiol. 2002, 11, 233–234. [Google Scholar]
- Miller, E.R.; Ullrey, D.E. The pig as a model for human nutrition. Ann. Rev. Nutr. 1987, 7, 361–382. [Google Scholar] [CrossRef] [PubMed]
- Shulman, R.J.; Henning, S.J.; Nichols, B.J. The miniature pig as an animal model for the study of intestinal enzyme development. Pediatr. Res. 1988, 23, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Kararli, T.T. Comparison of the gastrointestinal anatomy, physiology, and biochemistry of humans and commonly used laboratory animals. Biopharm. Drug Dispos. 1995, 16, 351–380. [Google Scholar] [CrossRef]
- Newport, M.J.; Henschel, M.J. Evaluation of the neonatal pig as a model for infant nutrition: Effects of different proportions of casein and whey protein in milk on nitrogen metabolism and composition of digesta in the stomach. Pediatr. Res. 1984, 18, 658–662. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.H. Animal models in nutrition research. J. Nutr. 2008, 138, 391–396. [Google Scholar] [PubMed]
- Walters, E.M.; Agca, Y.; Ganjam, V.; Ecans, T. Animal models got you puzzled?: Think pig. Ann. N. Y. Acad. Sci. 2011, 1245, 63–64. [Google Scholar] [CrossRef] [PubMed]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J., Jr.; Frazier, K.S. Swine as models in biomedical research and toxicology testing. Vet. Pathol. 2012, 49, 344–356. [Google Scholar] [CrossRef] [PubMed]
- Pekas, J.C.; Hays, V.W.; Thompson, A.M. Exclusion of the exocrine pancreatic secretion: Effect on digestibility of soybean and milk protein by baby pigs at various ages. J. Nutr. 1964, 82, 277–286. [Google Scholar] [PubMed]
- Imondi, A.R.; Stradley, R.P.; Wolgemuth, R. Enzyme replacement therapy in the pancreatic duct ligated swine. Proc. Soc. Exp. Biol. Med. 1972, 141, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Corring, T.; Bourdon, D. Exclusion of pancreatic exocrine secretion from intestine in the pig: Existence of a digestive compensation. J. Nutr. 1977, 107, 1216–1221. [Google Scholar] [PubMed]
- Rahko, T.; Gronlund, A.M.; Kalima, T.V. A new animal model for exocrine pancreatic insufficiency. Pathol. Res. Pract. 1985, 180, 309. [Google Scholar]
- Abello, J.; Pascaud, X.; Simoes-Nunes, C.; Cuber, J.C.; Junien, J.L.; Rozé, C. Total pancreatic insufficiency in pigs: A model to study intestinal enzymes and plasma levels of digestive hormones after pancreatic supplementation by a whole pancreas preparation. Pancreas 1998, 4, 556–564. [Google Scholar] [CrossRef]
- Saloniemi, H.; Kalima, T.V.; Rahko, T. Pancreatic enzyme supplementation in normal and exocrine pancreatic insufficient pigs. Acta Vet. Scand. 1989, 30, 367–370. [Google Scholar] [PubMed]
- Guillot, E.; Lemarchal, P.; Dhorne, T.; Rerat, A. Intestinal absorption of medium chain fatty acids: In vivo studies in pigs devoid of exocrine pancreatic secretion. Br. J. Nutr. 1994, 72, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Gregory, P.C.; Tabeling, R.; Kamphues, J. Growth and digestion in pancreatic duct ligated pigs: Effect of enzyme supplementation. In Biology of the Pancreas in Growing Animals; Pierzynowski, S.G., Zabielski, R., Eds.; Elsevier Science BV: Amsterdam, The Netherlands, 1999; pp. 381–393. [Google Scholar]
- Rengman, S.; Fedkiv, O.; Botermans, J.; Svendsen, J.; Weström, B.; Pierzynowski, S. An elemental diet fed, enteral or parenteral, does not support growth in young pigs with exocrine pancreatic insufficiency. Clin. Nutr. 2009, 28, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, J.; Fed’kiv, O.; Pawłowska, M.; Kowalik, S.; Erlwanger, K.; Weström, B.; Kruszewska, D.; Pierzynowski, S. The effectiveness of enzymatic replacement therapy measured by turbidimetry and the lipaemic index in exocrine pancreatic insufficient young, growing pigs, fed a high-fat diet. Adv. Med. Sci. 2009, 54, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Fedkiv, O.; Rengman, S.; Westrom, B.R.; Pierzynowski, S.G. Growth is dependent on the exocrine pancreas function in young weaners but not in growing-finishing pigs. J. Physiol. Pharm. 2009, 60 (Suppl. 3), 55–59. [Google Scholar]
- Schwarzmaier, T.E. Studies on the Effects of Substitution of Pancreatic Enzymes as Well as Special Supplementation of Vitamin A and E in Growing Pancreatic Duct Ligated Pigs as a Model for Pancreatic Insufficiency in Children. Doctoral Thesis, University of Veterinary Medicine, Foundation Hannover, Hannover, Germany, 2012. [Google Scholar]
- Mößeler, A.; Tabeling, R.; Gregory, P.C.; Kamphues, J. Compensatory digestion of fat, protein and starch (rates and amounts) in the large intestine of minipigs in case of reduced precaecal digestion due to pancreatic duct ligation—A short review. Livest. Sci. 2007, 109, 50–52. [Google Scholar]
- Mößeler, A.; Bergemann, J.; Becker, C.; Stemme, K.; Gregory, P.C.; Kamphues, J. NBT-PABA test to assess efficiency and kinetics of substituted proteolytic enzyme action in pancreatic duct ligated minipigs. J. Anim. Physiol. Anim. Nutr. 2008, 92, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Mößeler, A.; Schwarzmaier, T.; Gregory, P.C.; Piechotta, M.; Beyerbach, M.; Kamphues, J. Pancreatic exocrine insufficiency affects not only digestibility of nutrients and growth, but also body composition and endocrinological parameters—Study on piglets used as a model for children. Pancreat. Disord. Ther. 2015, 5. [Google Scholar] [CrossRef]
- Pierzynowski, S.; Szwiec, K.; Valverde Piedra, J.L.; Gruijc, D.; Szymanczyk, S.; Swieboda, P.; Prykhodko, O.; Fedkiv, O.; Kruszewska, D.; Filip, R.; et al. Exogenous pancreatic-like enzymes are recovered in the gut and improve growth of exocrine pancreatic insufficient pigs. J. Anim. Sci. 2012, 90, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Prykhodko, O.; Fedkiv, O.; Weström, B.R.; Pierzynowski, S.G. Effects on gut properties in exocrine pancreatic insufficient (EPI) pigs, being growth retarded due to pancreatic duct ligation at 7 weeks but not at 16 weeks of age. Adv. Med. Sci. 2014, 59, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Layer, P.; Keller, J. Lipase supplementation therapy: Standards, alternatives, and perspectives. Pancreas 2003, 26, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Borowitz, D.; Konstan, M.W.; O'Rourke, A.; Cohen, M.; Hendeles, L.; Murray, F.T. Coefficients of fat and nitrogen absorption in healthy subjects and individuals with cystic fibrosis. J. Pediatr. Pharmacol. Ther. 2007, 12, 47–52. [Google Scholar] [PubMed]
- Tabeling, R. Studies on the Effects of Enzyme Substitution on Nutrient Digestibilities (Pre-Caecal/Total) in Pancreas Ligated Pigs. Doctoral Thesis, University of Veterinary Medicine, Foundation Hannover, Hannover, Germany, 1998. [Google Scholar]
- Schönfelder, A. Fett for Life; Mylan Healthcare GmbH: Hanover, Germany, 2012. [Google Scholar]
- Mößeler, A.; Schwarzmaier, T.; Beyerbach, M.; Kamphues, J. Is starch digestion a point of interest in patients with exocrine pancreatic insufficiency? Pancreatology 2016, 16. [Google Scholar] [CrossRef]
- Gheorghe, C.; Seicean, A.; Saftoiu, A.; Tantau, M.; Dumitru, E.; Jinga, M.; Negreanu, L.; Mateescu, B.; Gheorghe, L.; Ciocirlan, M.; et al. Romanian Guidelines on the Diagnosis and Treatment of Exocrine Pancreatic Insufficiency. J. Gastroint. Liver Dis. 2015, 24, 117–123. [Google Scholar]
- Drochner, W.; Meyer, H. Digestion of organic matter in the large intestine of ruminants, horses, pigs and dogs. Adv. Anim. Phys. Anim. Nutr. 1991, 22, 18–40. [Google Scholar]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.E.; MacFarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Eisenmann, A.; Amann, A.; Said, M.; Datta, B.; Ledochowski, M. Implementation and interpretation of hydrogen breath tests. J. Breath Res. 2008. [Google Scholar] [CrossRef] [PubMed]
- Tabeling, R.; Gregory, P.; Kamphues, J. Studies on nutrient digestibilities (precaecal and total) in pancreatic duct ligated pigs and the effects of enzyme substitution. J. Anim. Physiol. Anim. Nutr. 1999, 82, 251–263. [Google Scholar] [CrossRef]
- Windey, K.; De Preter, V.; Verbeke, K. Relevance of protein fermentation to gut health. Mol. Nutr. Food Res. 2011, 56, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Pieper, R.; Boudry, C.; Bindelle, J.; Vahjen, W.; Zentek, J. Interaction between dietary protein content and the source of carbohydrates along the gastrointestinal tract of weaned piglets. Arch. Anim. Nutr. 2014, 68, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Easter, R.A.; Tanksley, T.D. A technique for re-entrant ileocaecal cannulation of swine. J. Anim. Sci. 1973, 36, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Hazem, A.S.; Drochner, W. Extracorporeal ileocecal bypass avoiding the ileocecal value in swine. Zeitschr. Versuchstierk. 1976, 18, 303–306. [Google Scholar]
- Green, S.; Bertrand, S.L.; Duron, M.J.C.; Maillard, R.A. Digestibility of amino acids in maize and barley meal, measured in pigs with ileo-rectal anastomosis and isolation of the large intestine. J. Sci. Food Agric. 1987, 41, 29–43. [Google Scholar] [CrossRef]
- Roth, F.X.; Kettler, S.I.; Roth-Maier, D.A.; Kirchgessner, M. Comparative evaluation of two different ileorectal-anastomosis techniques for determination of the apparent praecaecal digestibility of amino acids. Agribiol. Res. 1999, 52, 347–359. [Google Scholar]
- Ishihara, R.; Gregory, P.; Koch, H.F.; Kolleck, S.; Mößeler, A.; Kamphues, J.; Uchida, I. Comparison Study of Pharmacological Efficacy of Pancreatic Digestive Enzyme Replacement (LipaCreon Granules) and of Digestive Enzymes (Berizym Combination Granules and Excelase Combination Granules) on Digestibility of Fat, Protein and Starch in a Minipig Model of Pancreatic Exocrine Insufficiency (in vivo). J. New Rem. Clin. 2012, 61, 1044–1053. [Google Scholar]
- Grujic, D.; Fedkiv, O.; Nikandrova, E.; Mosiejchuk, N.; Kirko, S.; Osadchenko, I.; Kovalenko, T.; Drahanchuk, O.; Parshykov, O.; Goncharova, E.; et al. Nutritional formula pre-hydrolyzed by novel point of care lipase device (EFICTM) leads to increased fat absorption in young pigs with exocrine pancreatic insufficiency (EPI). J. Cyst. Fibros. 2015, 14, S50. [Google Scholar] [CrossRef]
- Kammlott, E.; Karthoff, J.; Stemme, K.; Gregory, P.; Kamphues, J. Experiments to optimize enzyme substitution therapy in pancreatic duct-ligated pigs. J. Anim. Physiol. Anim. Nutr. 2005, 89, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Mößeler, A.; Ahlfänger, B.; Kamphues, J. Is there need for a multienzyme product substitution when using high calorie liquid diets in patients with exocrine pancreatic insufficiency? Studies on praecaecal digestibility in pigs with experimentally induced pancreatic exocrine insufficiency. J. Cyst. Fibr. 2016, 15, S29. [Google Scholar] [CrossRef]
- Mößeler, A.; Kramer, N.; Becker, C.; Gregory, P.C.; Kamphues, J. Prececal digestibility of various sources of starch in minipigs with or without experimentally induced exocrine pancreatic insufficiency. J. Anim. Sci. 2012, 90, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Mößeler, A.; Vagt, S.; Beyerbach, M.; Kamphues, J. Starch origin and thermal processing affect starch digestion in a minipig model of pancreatic exocrine insufficiency. Gastroenterol. Res. Pract. 2015. [Google Scholar] [CrossRef] [PubMed]
- Stefaniak, N.B.J. Effects of a Galenic Modification of a Porcine Pancreatin-Product on the Efficacy—Tested in Pancreatic Duct-Ligated, Ileocecally Fistulated Minipigs. Doctoral Thesis, University of Veterinary Medicine, Foundation Hannover, Hanover, Germany, 2015. [Google Scholar]
- Mandischer, C. Studies on the Digestibility (Pre-Caecal/Total) of a High Starch Compound Feed in Pancreatic Duct Ligated Pigs and the Effects of Oral Enzyme Supplementation with Two Different Products. Doctoral Thesis, University of Veterinary Medicine, Foundation Hannover, Hanover, Germany, 2002. [Google Scholar]
- Koch, J. Studies on the Efficiency of an Established Multi-Enzyme Product of Porcine Origin Compared to Two Microbial Redevelopments in Pancreatic Duct Ligated, Ileocaecal Fistulated Minipigs. Doctoral Thesis, University of Veterinary Medicine, Foundation Hannover, Hanover, Germany, 2011. [Google Scholar]
- Kalla, K. Effects of the Combination of a Porcine Multienzyme Product with a Microbial Lipase on Digestibility (Praecaecal/in Toto) in Pancreatic Duct Ligated, Ileo-Caecal Fistulated Minipigs. Doctoral Thesis, University of Veterinary Medicine, Foundation Hannover, Hanover, Germany, 2009. [Google Scholar]
- Classen, J. Development of a Novel Screening Test to Assess the Efficacy of Multi-Enzyme Products by Using Pancreatic Duct Ligated, Ileo-Caecal Fistulated Minipigs. Doctoral Thesis, University of Veterinary Medicine, Foundation Hannover, Hanover, Germany, 2008. [Google Scholar]
- Karthoff, J. Evaluation of the Effectiveness of Enzyme Substitution in the Pancreatic Duct Ligated Minipig Model of Pancreatic Exocrine Insufficiency: Use of Various Methods and Parameters (Nutrient Digestibility, Concentrations of Absorbed Nutrients as Well as of Different Test Substances in Blood). Doctoral Thesis, University of Veterinary Medicine, Foundation Hannover, Hanover, Germany, 2004. [Google Scholar]
- Heldt, D. Studies of the Effects of Enzyme Supplementation with Two Different Products on Nutrient Digestibility (Pre-Caecal/Total) in Pancreatic Duct Ligated Pigs Fed a High Fat Diet. Doctoral Thesis, University of Veterinary Medicine, Foundation Hannover, Hanover, Germany, 2001. [Google Scholar]
- Fuente-Dege, A.M. Studies on the Relationship between Pre-Caecal Digestibility of Nutrients and the Digestibility over the Whole Gastrointestinal Tract in Pancreatic Duct Ligated Pigs. Doctoral Thesis, University of Veterinary Medicine, Foundation Hannover, Hanover, Germany, 2003. [Google Scholar]
- Fassmann, C. Studies on the Extent and Location of Nutrient Digestibilities of High-Fat Diets by Pancreatic Duct Ligated Pigs. Doctoral Thesis, University of Veterinary Medicine, Foundation Hannover, Hanover, Germany, 2001. [Google Scholar]
- Harris, R.; Norman, A.F.; Payne, W.W. The effect of pancreatin therapy on fat absorption and nitrogen retention in children with fibrocystic disease of the pancreas. Arch. Dis. Child. 1955, 30, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Konstan, M.W.; Liou, T.G.; Strausbaugh, S.D.; Ahrens, R.; Kanga, J.F.; Graff, G.R.; Moffett, K.; Millard, S.L.; Nasr, S.Z.; Siméon, E.; et al. Efficacy and Safety of a New Formulation of Pancrelipase (Ultrase MT20) in the Treatment of Malabsorption in Exocrine Pancreatic Insufficiency in Cystic Fibrosis. Gastroenterol. Res. Pract. 2010. [Google Scholar] [CrossRef] [PubMed]
- Löhr, M.; Oliver, M.R.; Frulloni, L. Synopsis of recent guidelines on pancreatic exocrine insufficiency. United Eur. Gastroenterol. J. 2013, 1, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Vagt, S. In Vivo and in Vitro Investigations on Prececal Starch Digestion (Related to Source of Starch and Thermal Treatment) in Pancreatic Duct Ligated Minipigs—Used as a Model for Human Exocrine Pancreatic Insufficiency. Doctoral Thesis, University of Veterinary Medicine, Foundation Hannover, Hannover, Germany, 2014. [Google Scholar]
- Turck, D.; Braegger, C.P.; Colombo, C.; Declercq, D.; Morton, A.; Pancheva, R.; Robberecht, E.; Stern, M.; Strandvik, B.; Wolfe, S.; et al. ESPEN-ESPGHAN-ECFS guidelines on nutrition care for infants, children, and adults with cystic fibrosis. Clin. Nutr. 2016. [Google Scholar] [CrossRef] [PubMed]
- Lückerath, E.; Müller, S.D. Diätetik und Ernährungsberatung; Das Praxisbuch. 4. Auflage; Haug Verlag: Stuttgart, Germany, 2011. [Google Scholar]
- Airinei, G.; Gaudichon, C.; Bos, C.; Bon, C.; Kapel, N.; Bejou, B.; Raynaud, J.J.; Luengo, C.; Aparicio, T.; Levy, P.; et al. Postprandial protein metabolism but not a fecal test reveals protein malabsorption in patients with pancreatic exocrine insufficiency. Clin. Nutr. 2011, 30, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Amarri, S.; Harding, M.; Coward, W.A.; Evans, T.J.; Weaver, L.T. 13C and H2 breath tests to study extent and site of starch digestion in children with cystic fibrosis. J. Pediatr. Gastr. Nutr. 1999, 29, 327–331. [Google Scholar] [CrossRef]
- Hiele, M.; Ghoos, Y.; Rutgeerts, P.; Vantrappen, G. Starch digestion in normal subjects and patients with pancreatic disease, using a 13CO2 breath test. Gastroenterology 1989, 96, 503–509. [Google Scholar] [CrossRef]
- Fuller, M.F. Methodologies for the measurement of digestion. In Digestive Physiology in Pigs, Proceedings of the 5th International Symposium, Wageningen, The Netherlands, 24–26 April 1991; Versteegen, M.W.A., Huisman, J., Den Hartog, L.A., Eds.; EAAP Publ.: Wageningen, The Netherlands, 1991; Volume 54, pp. 273–288. [Google Scholar]
- Gregory, P.C.; Hoffmann, K.; Kamphues, J.; Mößeler, A. The pancreatic duct ligated (mini)pig as model for pancreatic exocrine insufficiency in man. Pancreas 2016, 45, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
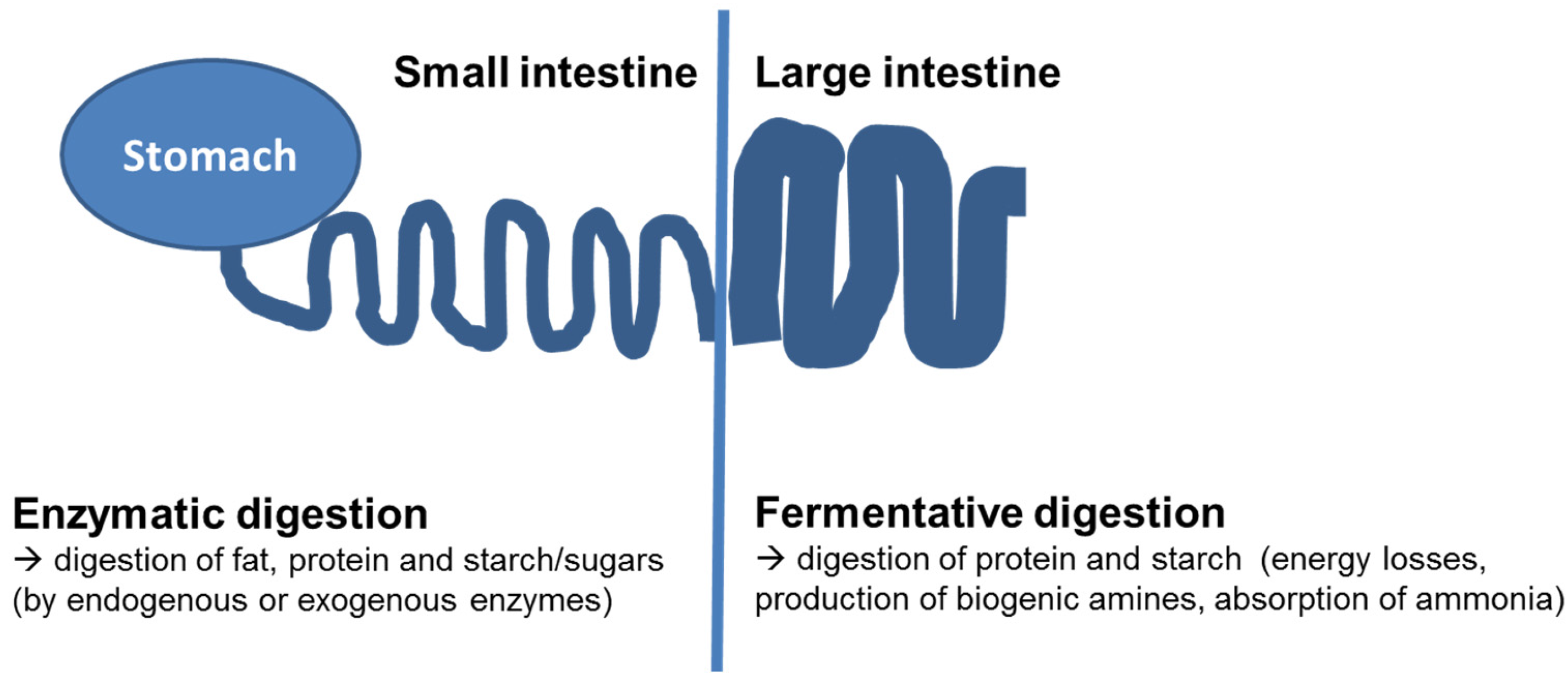


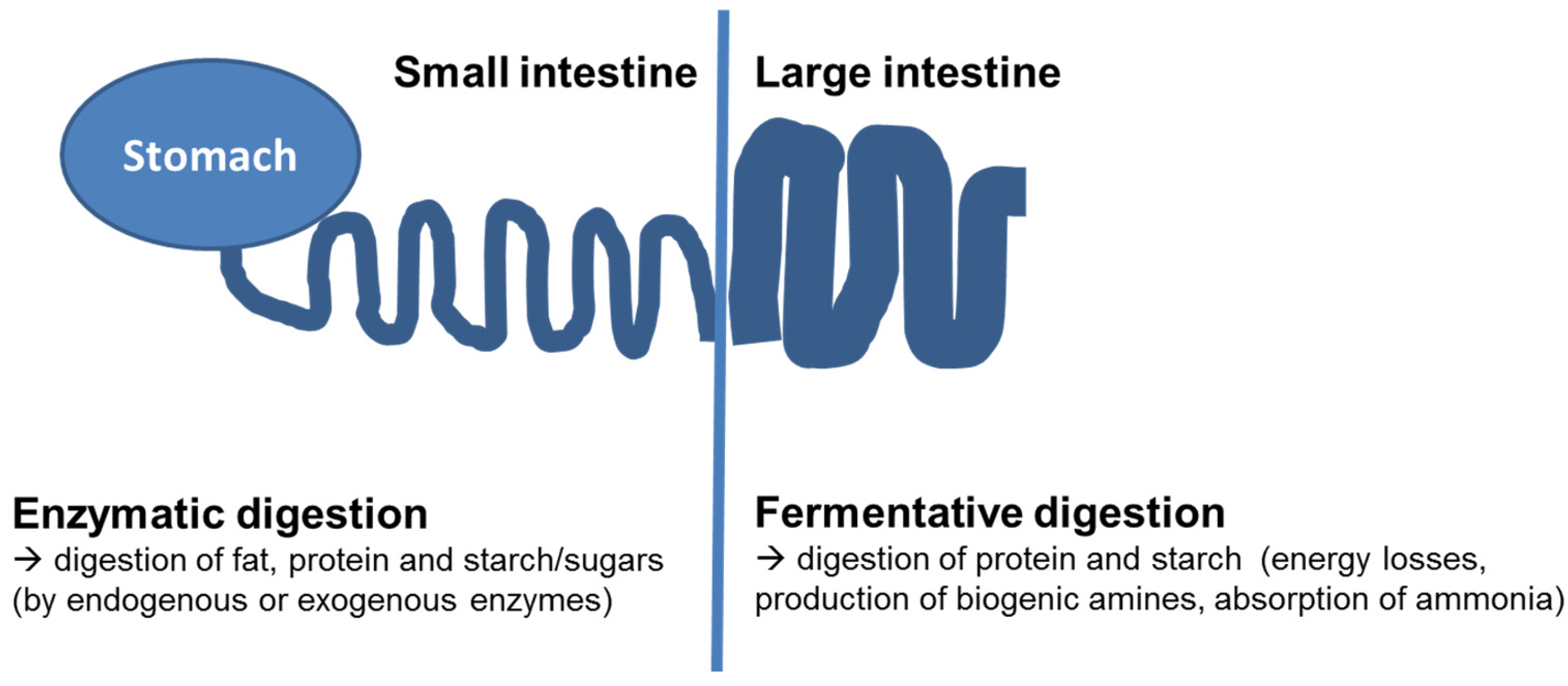{kind=link}
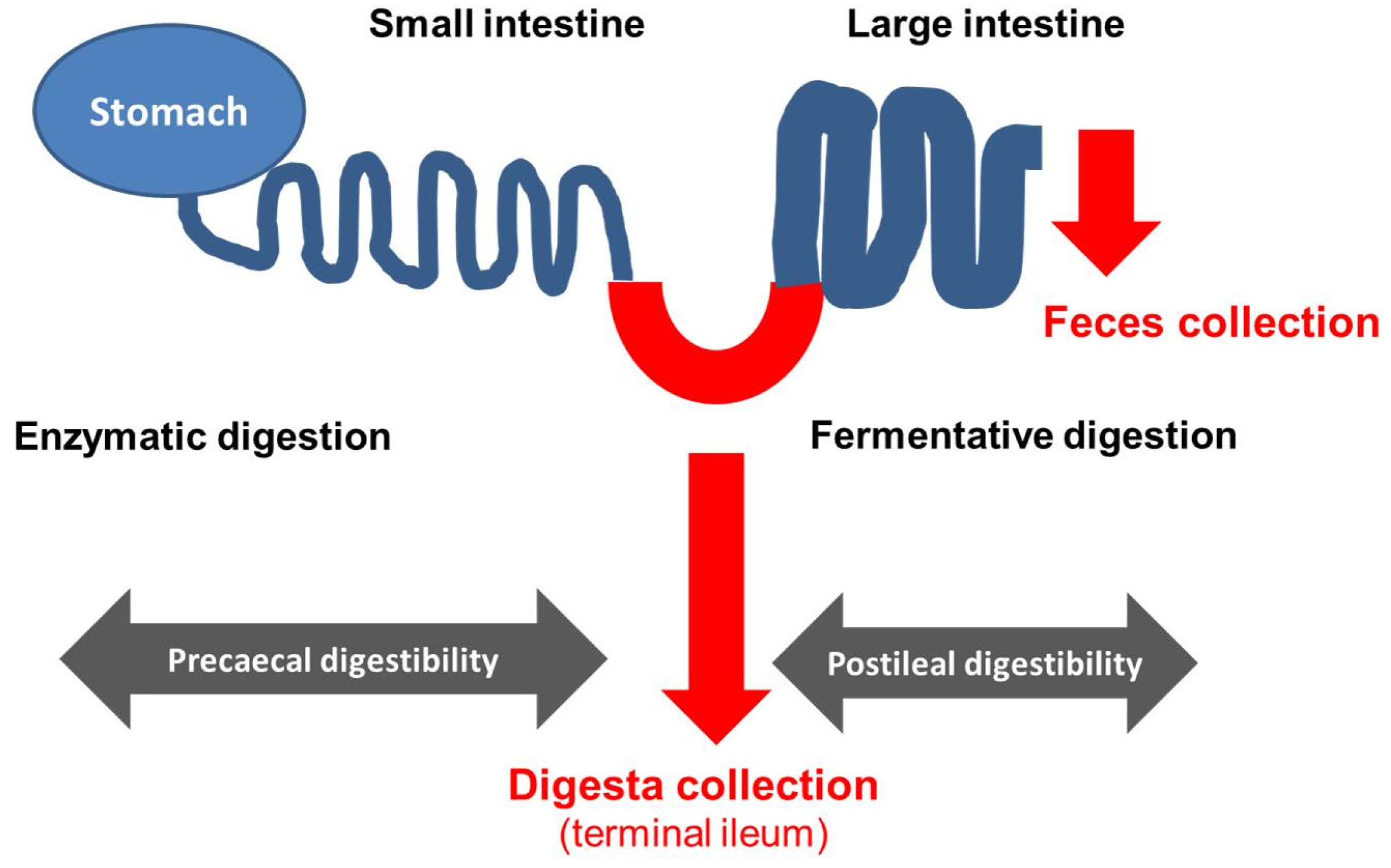{kind=link}
| Starch | Crude Fat | Crude Protein | Author |
|---|---|---|---|
| 60.0 | 9.01 | 12.8 | Tabeling (Starch Diet) [41] |
| 56.2 | 2.21 | 21.2 | Mandischer [62] |
| 41.2 | 29.7 | 15.4 | Stefaniak [61] |
| 40.0 | 19.0 | 18.3 | Kammlott [57] |
| 39.0 | 32.7 | 14.9 | Koch [63] |
| 37.8 | 28.3 | 15.6 | Kalla [64] |
| 35.8 | 31.3 | 14.6 | Classen [65] |
| 33.4 | 34.9 | 14.9 | Karthoff [66] |
| 27.3 | 34.2 | 16.3 | Heldt [67] |
| 27.1 | 33.5 | 16.7 | Fuente-Dege [68] |
| 26.6 | 32.2 | 15.9 | Tabeling (Fat Diet) [41] |
| 24.1 | 30.1 | 14.4 | Fassmann [69] |
| Starch | Crude Fat | Crude Protein | Author | ||||
|---|---|---|---|---|---|---|---|
| Prc. | Total | Prc. | Total | Prc. | Total | ||
| 61.9 | 99.3 | 9.30 | -6.60 | 29.6 | 56.7 | Tabeling (Starch Diet) [41] | PL-pigs |
| 63.5 | N.A. | 28.6 | 8.43 | 30.5 | 67.5 | Mandischer [62] | |
| 65.7 | N.A. | 22.7 | 28.1 | 31.9 | 48.4 | Stefaniak [61] | |
| 72.6 | N.A. | 19.4 | 23.3 | 30.9 | 60.0 | Kammlott [57] | |
| 61.1 | N.A. | 16.2 | 26.2 | 28.2 | 47.9 | Koch [63] | |
| 50.0 | N.A. | 14.0 | 22.0 | 24.8 | 44.9 | Kalla [64] | |
| 61.4 | 98.4 | 25.1 | 23.1 | 33.7 | 35.1 | Classen [65] | |
| 75.4 | N.A. | 29.7 | 29.2 | 32.3 | 48.6 | Karthoff [66] | |
| 63.8 | N.A. | 29.0 | 17.8 | 33.7 | 54.1 | Heldt [67] | |
| 75.6 | N.A. | 29.9 | 28.2 | 40.5 | 54.1 | Fuente-Dege [68] | |
| 87.7 | 99.1 | 43.0 | 31.5 | 27.3 | 56.9 | Tabeling (Fat Diet) [41] | |
| 92.5 | N.A. | 32.5 | 14.5 | 26.1 | 49.9 | Fassmann [69] | |
| 94.6–99.7 | ~100 | 88.2–98.3 | 81.3–97.4 | 79.1–82.3 | 87.5–92.2 | Control-pigs (all studies) | |
| Control | PL-pig | |||
|---|---|---|---|---|
| Without PERT | With PERT # | |||
| Prececal digestibility (%) * | Fat | 93.9 | 22.7 (24.2) | 84.0 (89.5) |
| Protein | 79.9 | 31.9 (39.9) | 73.9 (92.5) | |
| Starch | 98.8 | 65.7 (66.5) | 94.8 (96.0) | |
| Digestible energy (DE; kJ) per g ingested nutrient ** | Fat | 36.2 | 8.74 (24.1) | 32.3 (89.2) |
| Protein | 13.7 | 5.49 (40.1) | 12.7 (92.7) | |
| Starch | 17.0 | 11.3 (66.5) | 16.3 (95.9) | |
| Amount of nutrient intake needed (g) to generate 1000 kJ DE *** | Fat | 27.6 | 114 (413) | 31.0 (112) |
| Protein | 73.0 | 182 (249) | 78.7 (108) | |
| Starch | 58.8 | 88.5 (150) | 61.3 (104) | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mößeler, A.; Kamphues , J. Black‐Box Gastrointestinal Tract—Needs and Prospects of Gaining Insights of Fate of Fat, Protein, and Starch in Case of Exocrine Pancreatic Insufficiency by Using Fistulated Pigs. Nutrients 2017, 9, 150. https://doi.org/10.3390/nu9020150
Mößeler A, Kamphues J. Black‐Box Gastrointestinal Tract—Needs and Prospects of Gaining Insights of Fate of Fat, Protein, and Starch in Case of Exocrine Pancreatic Insufficiency by Using Fistulated Pigs. Nutrients. 2017; 9(2):150. https://doi.org/10.3390/nu9020150
Chicago/Turabian StyleMößeler, Anne, and Josef Kamphues . 2017. "Black‐Box Gastrointestinal Tract—Needs and Prospects of Gaining Insights of Fate of Fat, Protein, and Starch in Case of Exocrine Pancreatic Insufficiency by Using Fistulated Pigs" Nutrients 9, no. 2: 150. https://doi.org/10.3390/nu9020150
APA StyleMößeler, A., & Kamphues , J. (2017). Black‐Box Gastrointestinal Tract—Needs and Prospects of Gaining Insights of Fate of Fat, Protein, and Starch in Case of Exocrine Pancreatic Insufficiency by Using Fistulated Pigs. Nutrients, 9(2), 150. https://doi.org/10.3390/nu9020150