Towards an Understanding of the Low Bioavailability of Quercetin: A Study of Its Interaction with Intestinal Lipids

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Preparation of Solutions
2.2.2. Solubility Measurements
2.2.3. UV-Visible and Fluorescence Spectroscopy
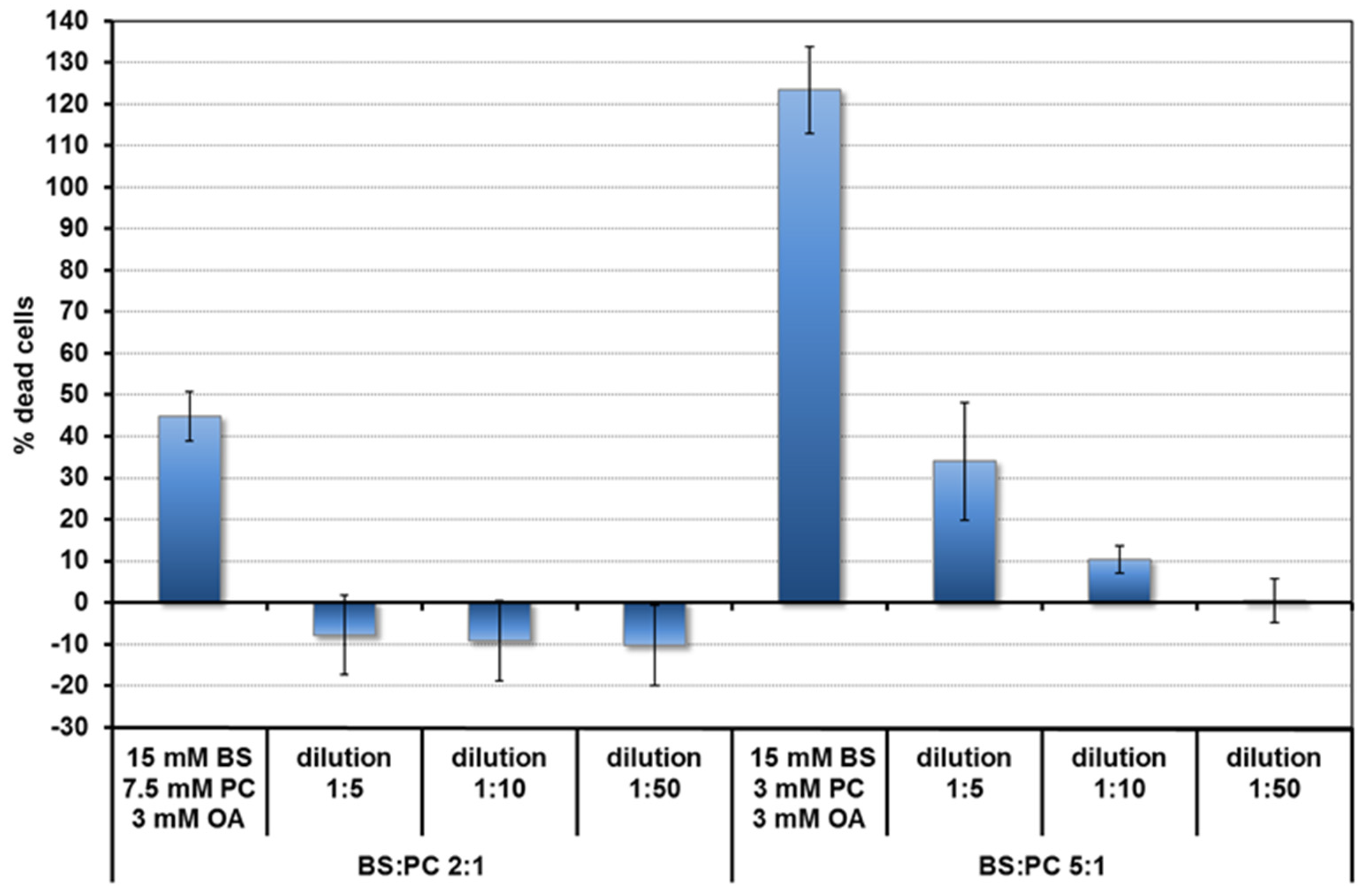
2.2.4. CaCo-2/TC7 Cell Culture and Quercetin Uptake
2.2.5. Centrifugal Ultrafiltration of Micelles and Liposomes
2.2.6. Quantification of Quercetin and BS
3. Results
3.1. Properties of Quercetin Relevant to Its Bioavailability
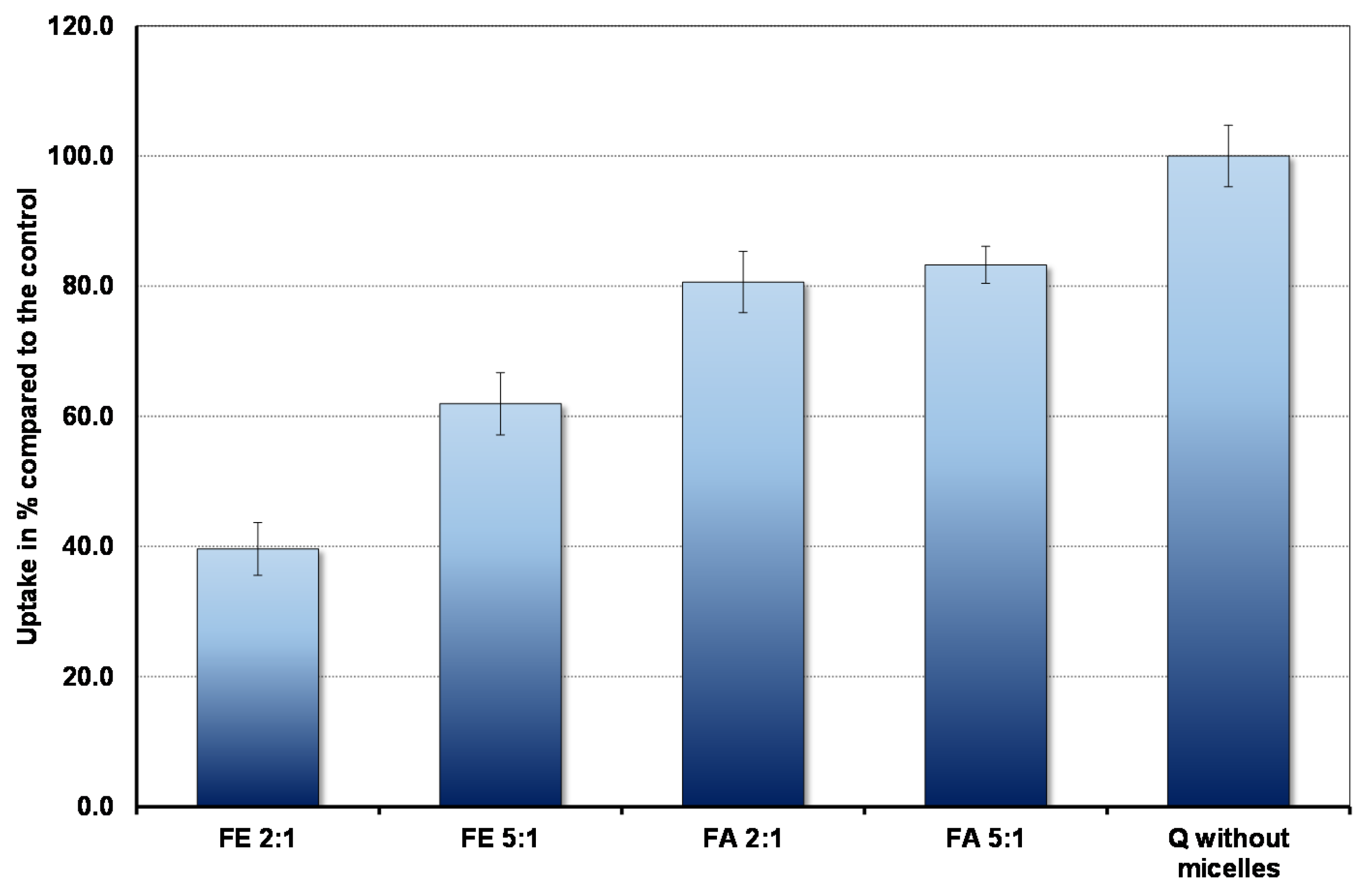
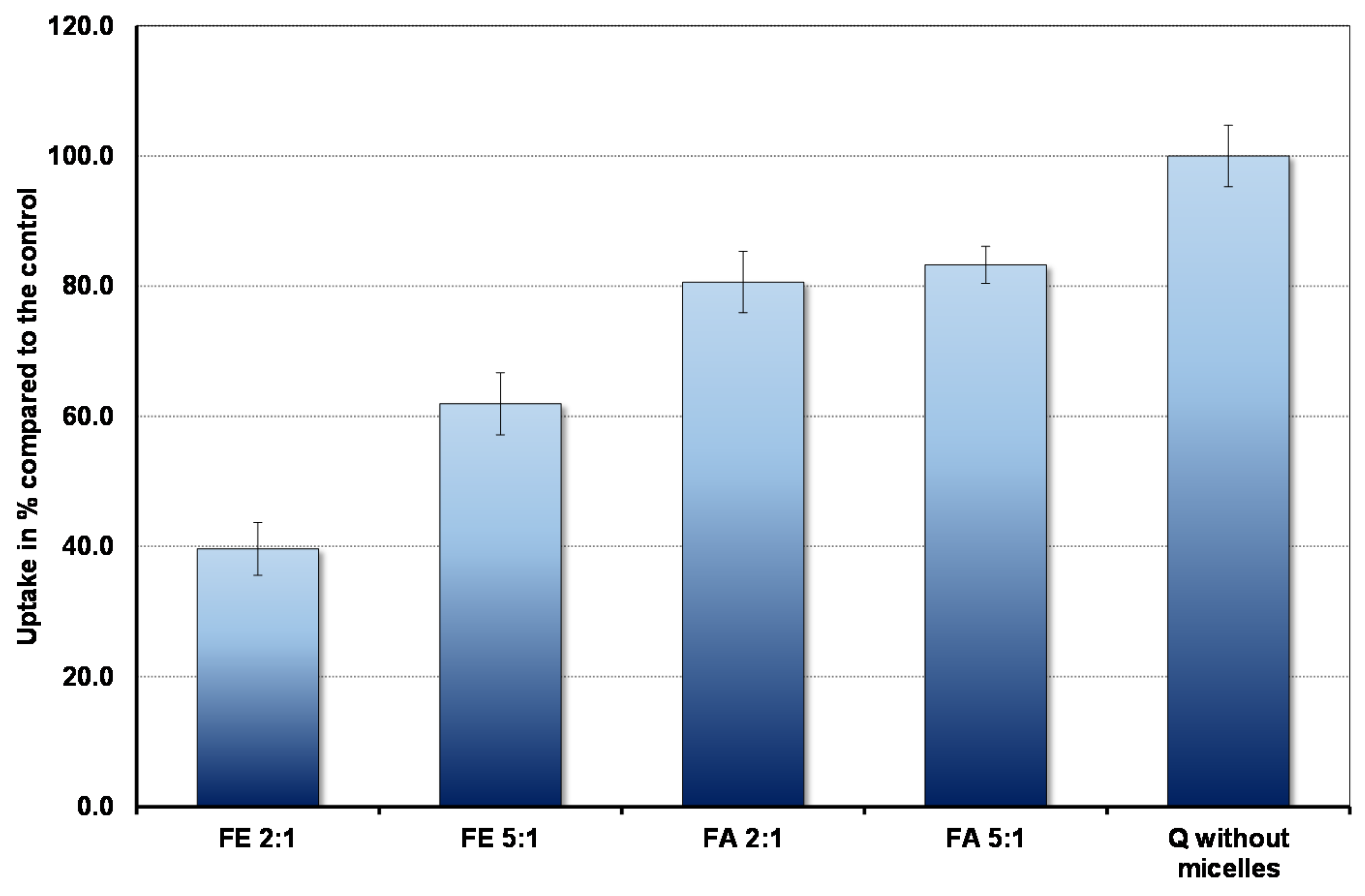
3.2. Uptake of Quercetin into Cells
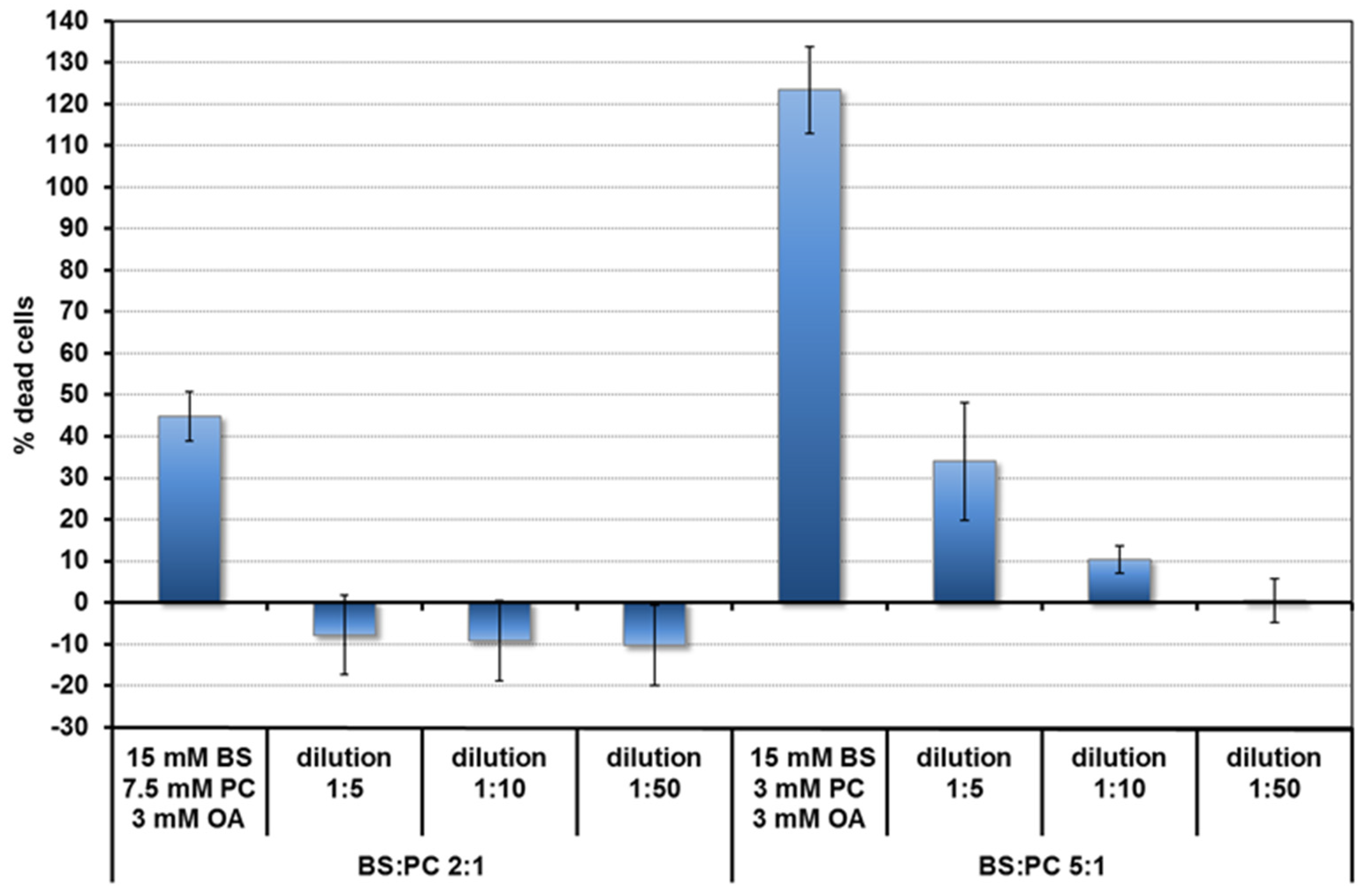
3.3. Nature of the Micelles and Quercetin–Micelle Interactions
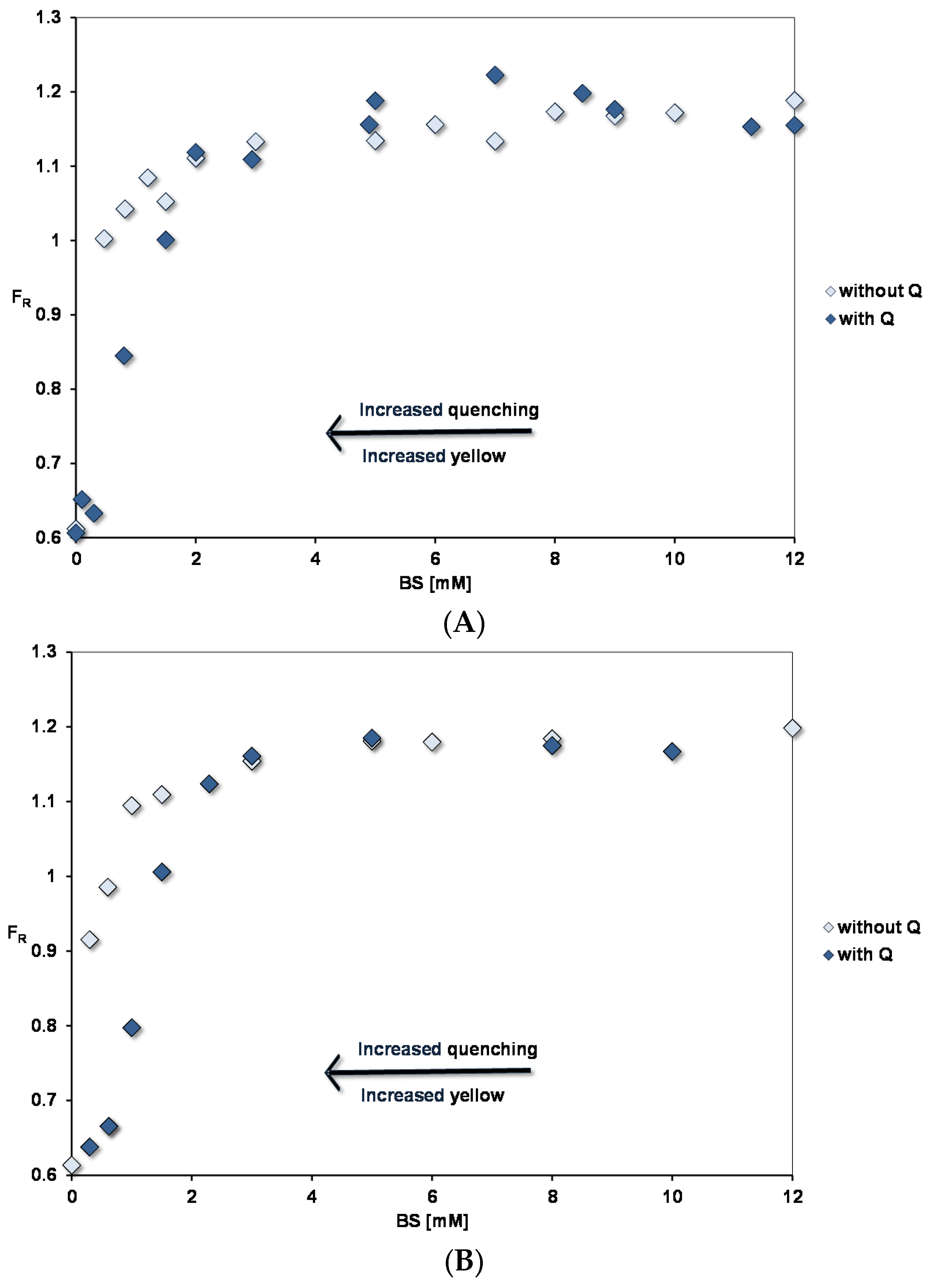
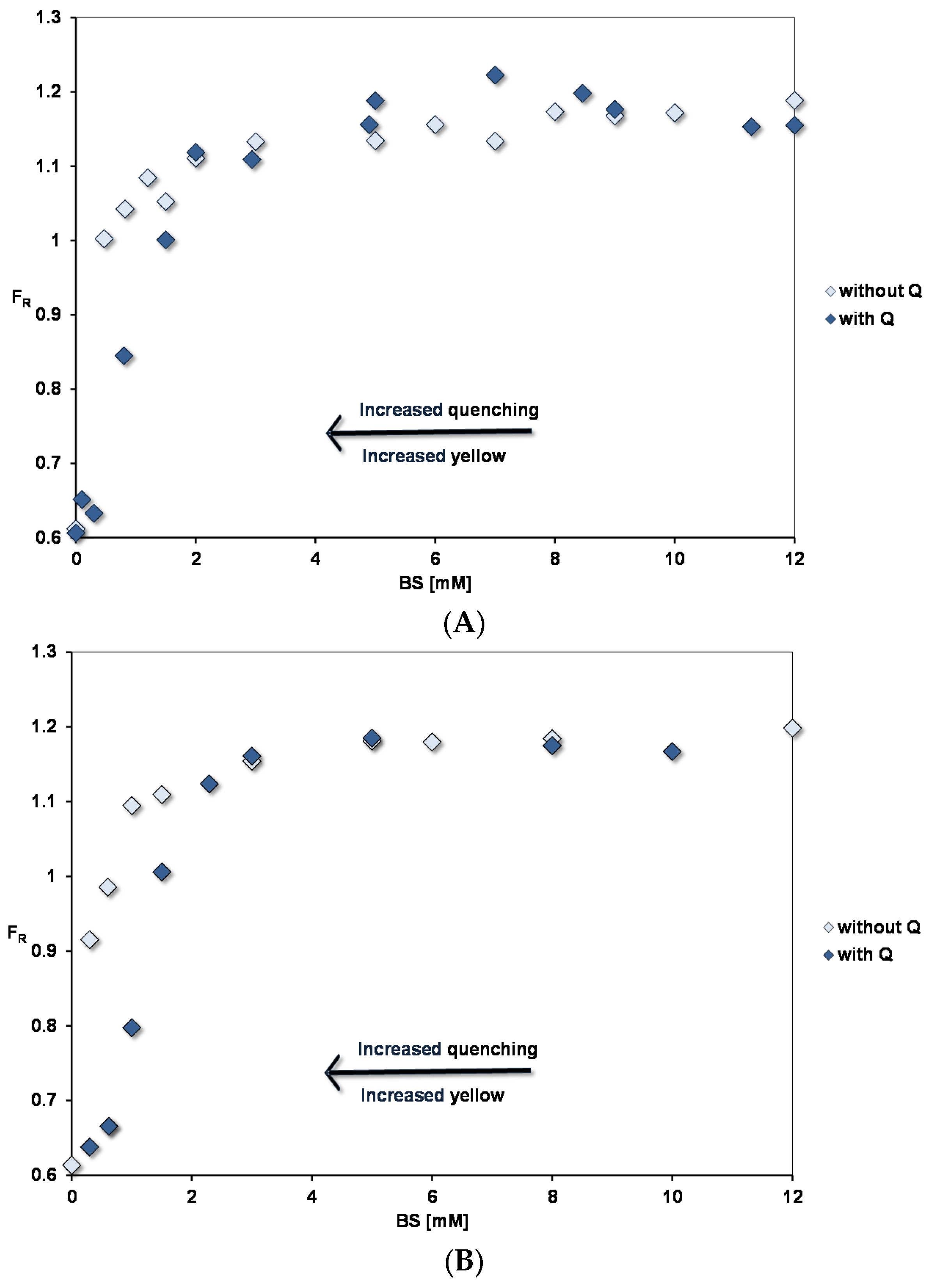
3.3.1. Pyrene Fluorescence
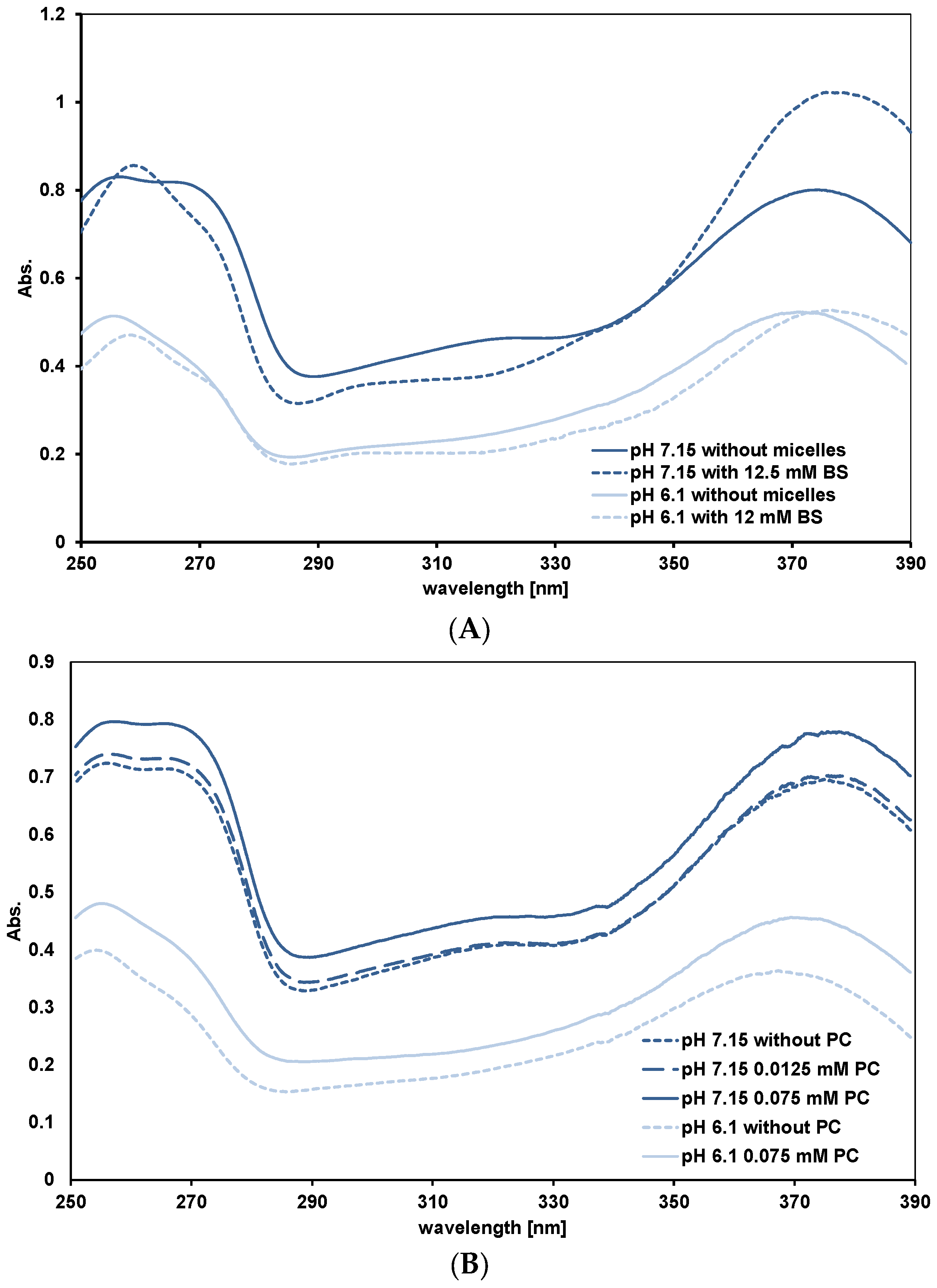
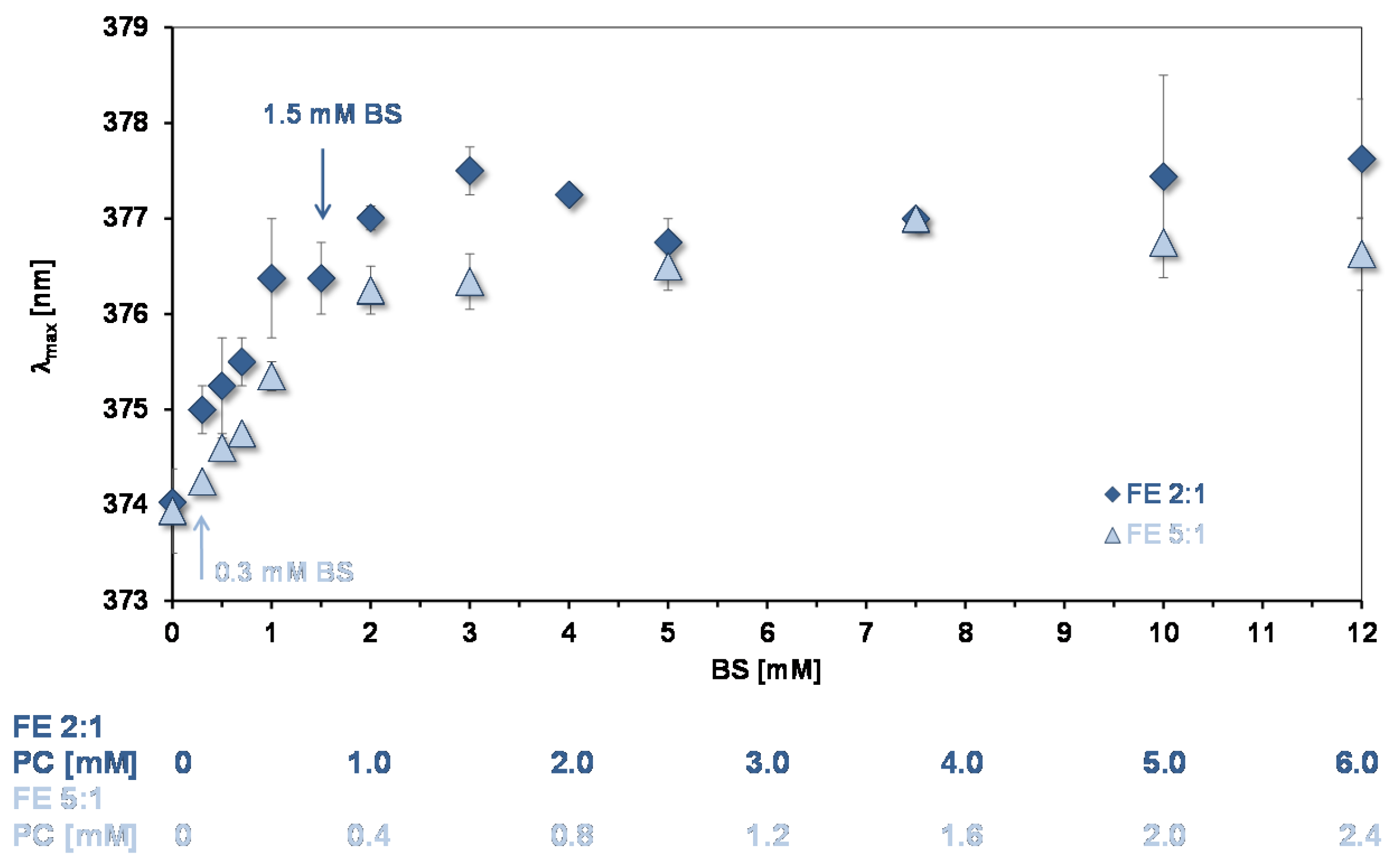
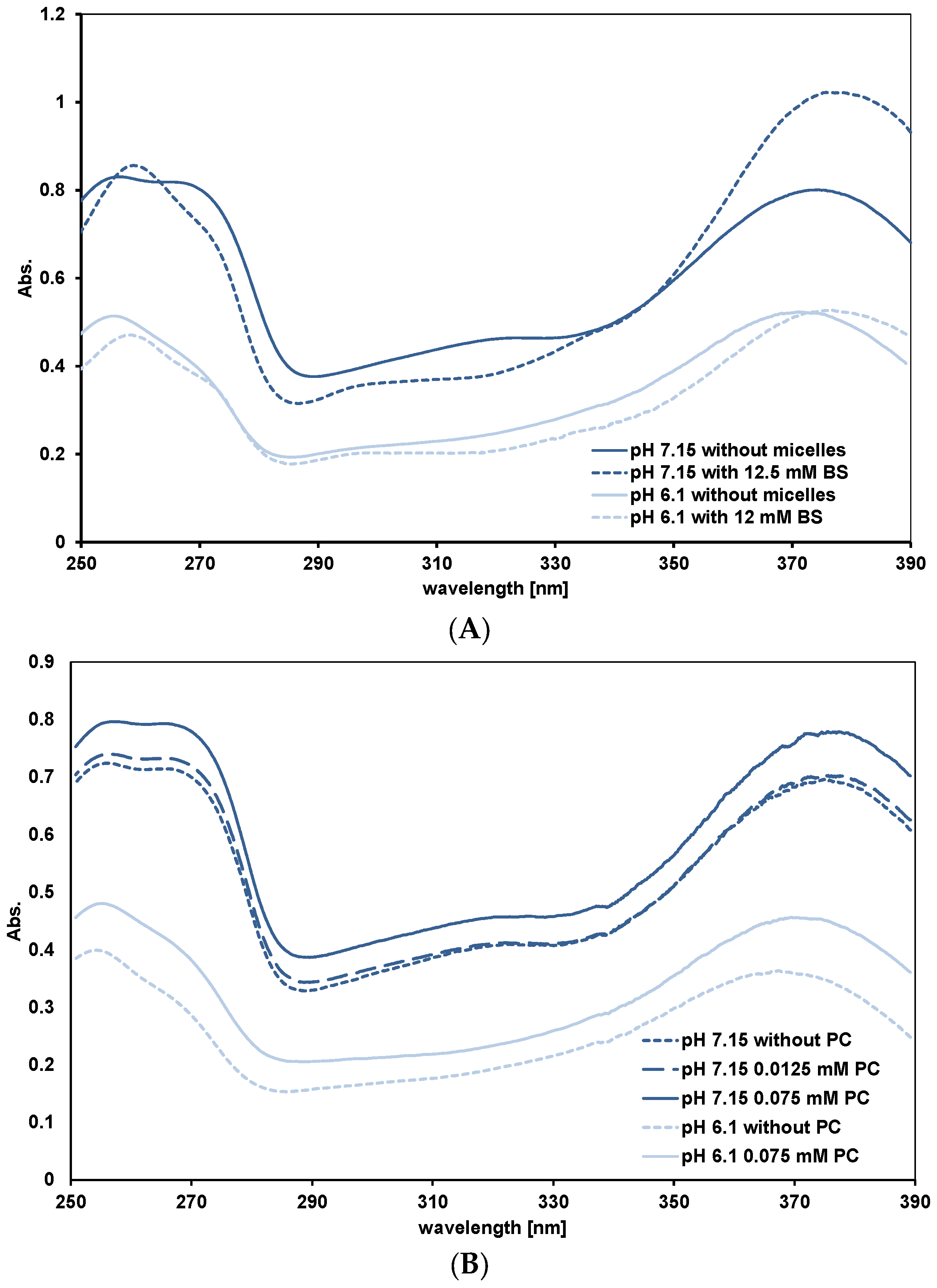
3.3.2. Quercetin Absorption Spectra
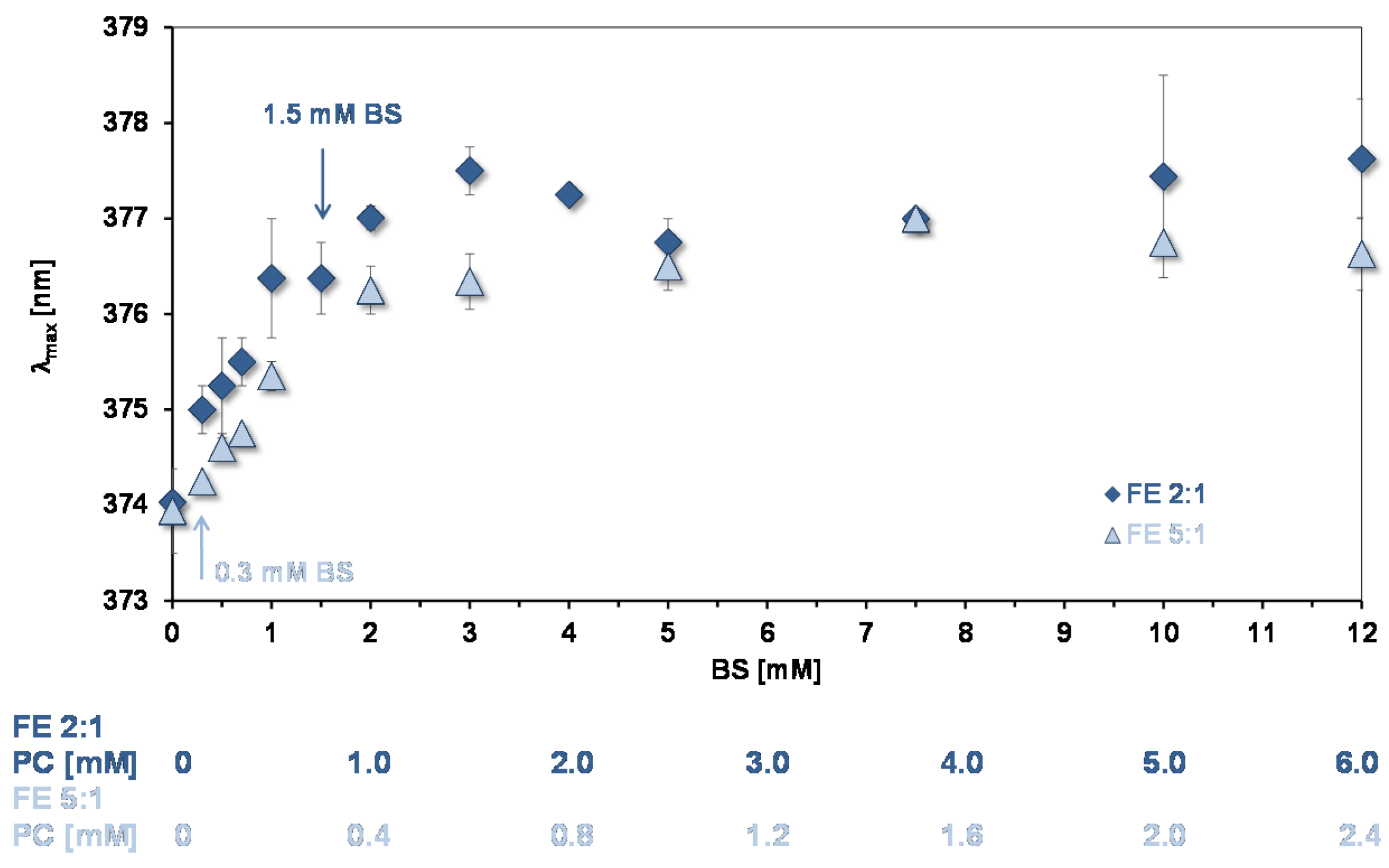
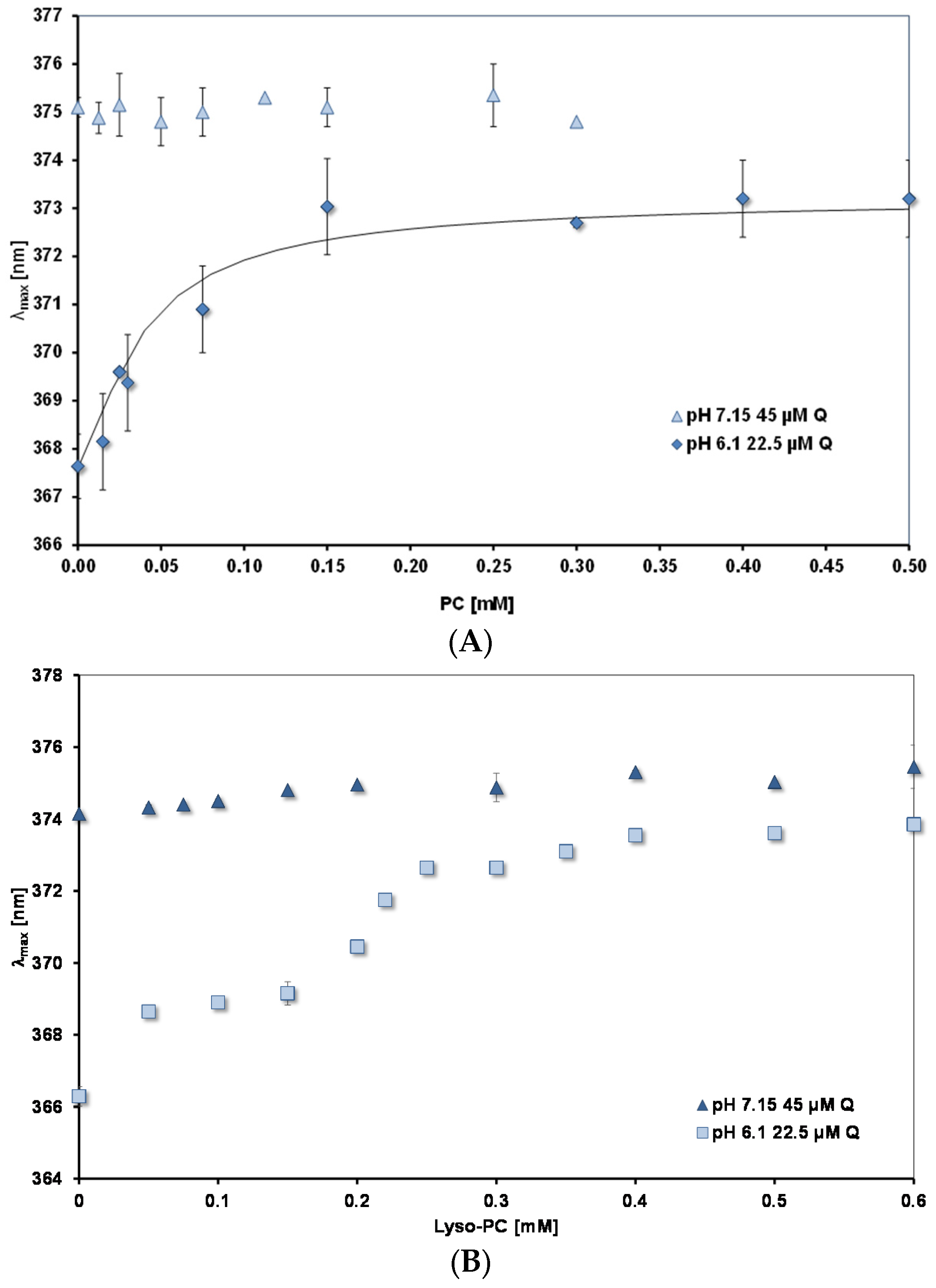
3.4. Affinity of Quercetin for Micelles and PC
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Hamalainen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: Genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-kappaB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-kappaB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediat. Inflamm. 2007, 2007, 45673. [Google Scholar] [CrossRef]
- Egert, S.; Bosy-Westphal, A.; Seiberl, J.; Kubitz, C.; Settler, U.; Plachta-Danielzik, S.; Wolffram, S. Quercetin reduces systolic blood pressure and plasma oxidized low-density lipoprotein concentrations in overweight subjects with a high-cardiovascular disease risk phenotype: A double-blinded, placebo-controlled cross-over study. Br. J. Nutr. 2009, 102, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Niaz, K.; Maqbool, F.; Hassen, F.I.; Abdollahi, M.; Venkata, K.C.N.; Nabari, S.M.; Bishayee, A. Molecular targets underlying the anticancer effects of quercetin: An update. Nutrients 2016, 8, 529. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, S.; Faris, A.N.; Comstock, A.T.; Wan, Q.; Nanua, S.; Hershenon, M.B.; Saijan, A.S. Quercetin inhibits rhinovirus replication in vitro and in vivo. Antivir. Res. 2012, 94, 258–271. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Bruno, R.S. Endogenous and exogenous mediators of quercetin bioavailability. J. Nutr. Biochem. 2015, 26, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yu, D.; Sun, J.; Liu, X.; Jiang, L.; Guo, H.; Ren, F. Interaction of plant phenols with food macronutrients: Characterisation and nutritional-physiological consequences. Nutr. Res. Rev. 2014, 27, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, K.; Plum, G.W.; Berrin, J.; Juge, N.; Jacob, R.; Naim, H.W.; Williamson, G.; Swallow, G.M.; Kroon, P.A. Deglycosylation by small intestinal β-glucosidases is a critical step in the absorption of dietary flavonoid glycosides in humans. Eur. J. Nutr. 2003, 42, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Day, A.J.; Canada, F.J.; Diaz, J.C.; Kroon, P.A.; Mclauchlin, R.; Faulds, C.B.; Plumb, G.W.; Morgan, M.R.S.; Williamson, G. Dietary flavonoid and isoflavone glycosides are hydrolysed by the lactase site of phlorizin hydrolase. FEBS Lett. 2000, 468, 166–170. [Google Scholar] [CrossRef]
- Buchweitz, M.; Kroon, P.A.; Rich, G.T.; Wilde, P.J. Quercetin solubilisation in bile salts: A comparison with sodium dodecyl sulphate. Food Chem. 2016, 211, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Barrington, R.D.; Needs, P.W.; Williamson, G.; Kroon, P.A. MK571 inhibits phase-2 conjugation of flavonols by Caco-2/TC7 cells, but does not specifically inhibit their apical efflux. Biochem. Pharmacol. 2015, 95, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Murota, K.; Shimizu, S.; Chujo, H.; Moon, J.H.; Terao, J. Efficiency of absorption and metabolic conversion of quercetin and its glucosides in human intestinal cell line Caco-2. Arch. Biochem. Biophys. 2000, 384, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, Y.; Ma, Y.; Yu, A.; Cai, F.; Shao, W.; Zhai, G. Formulation optimization and in situ absorption in rat intestinal tract of quercetin-loaded microemulsion. Colloid Surf. B 2009, 71, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Mah, E.; Davis, C.G.; Jalili, T.; Ferruzzi, M.G.; Chun, O.K.; Bruno, R.S. Dietary fat increases quercetin bioavailability in overweight adults. Mol. Nutr. Food Res. 2013, 57, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Lesser, S.; Cermak, R.; Wolffram, S. Bioavailability of quercetin in pigs is influenced by the dietary fat content. J. Nutr. 2004, 134, 1508–1511. [Google Scholar] [PubMed]
- Rothwell, J.A.; Day, A.J.; Morgan, M.R.A. Experimental determination of octanol-water partition coefficients of quercetin and related flavonoids. J. Agric. Food Chem. 2005, 53, 4355–4360. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Guo, R. Interaction between flavonoid, quercetin and surfactant aggregates with different charges. J. Coll. Interface Sci. 2006, 302, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Pawlikowska-Pawlega, B.; Dziubinska, H.; Krol, E.; Trebacz, K.; Jarosz-Wilkolazka, A.; Paduch, R.; Gawron, A.; Gruszecki, W.I. Characteristics of quercetin interactions with liposomal and vacuolar membranes. Biochim. Biophys. Acta 2014, 1838, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Zenkevich, I.G.; Eshchenko, A.Y.; Makarova, S.V.; Vitenberg, A.G.; Dobryakov, Y.G.; Utsal, V.A. Identification of the Products of Oxidation of Quercetin by Air Oxygen at Ambient Temperature. Molecules 2007, 12, 654–672. [Google Scholar] [CrossRef] [PubMed]
- Kalyanasundaram, K.; Thomas, J.K. Environmental effects on vibronic band intensities in pyrene monomer fluorescence and their application in studies of micellar systems. J. Am. Chem. Soc. 1977, 99, 2039–2044. [Google Scholar] [CrossRef]
- Zughaid, H.; Forbes, B.; Martin, G.P.; Patel, N. Bile salt composition is secondary to bile salt concentration in determining hydrocortisone and progesterone solubility in intestinal mimetic fluids. Int. J. Pharm. 2012, 422, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Carriere, F.; Renou, C.; Ransac, S.; Lopes, V.; De Caro, J.; Ferrato, F.; Beglinger, C. Inhibition of gastrointestinal lipolysis by Orlistat during digestion of test meals in healthy volunteers. Am. J. Physiol. 2001, 281, G16–G28. [Google Scholar]
- Donovan, J.M.; Jackson, A.A. Rapid determination by centrifugal ultrafiltration of inter-mixed micellar/vesicular (non-lecithin-associated) bile salt concentrations in model bile: Influence of Donnan equilibrium effects. J. Lipid Res. 1993, 34, 1121–1129. [Google Scholar] [PubMed]
- Rossi, S.S.; Converse, J.L.; Hofmann, A.F. High pressure liquid chromatography analysis of conjugated bile acids in human bile: Simultaneous resolution of sulfated and unsulfated lithocholyl amidates and the common conjugated bile acids. J. Lipid Res. 1987, 28, 589–595. [Google Scholar] [PubMed]
- Gordon, M.H.; Roedig-Penman, A. Antioxidant activity of quercetin and myricetin in liposomes. Chem. Phys. Lipids 1998, 97, 79–85. [Google Scholar] [CrossRef]
- Movileanu, L.; Neagoe, I.; Flonta, M.L. Interaction of the antioxidant flavonoid quercetin with planar lipid bilayers. Int. J. Pharm. 2000, 205, 135–146. [Google Scholar] [CrossRef]
- Madenci, D.; Egelhauf, S.U. Self-assembly in aqueous bile salt solutions. Curr. Opin. Colloid Interface 2010, 15, 109–115. [Google Scholar] [CrossRef]
- Crespy, V.; Morand, C.; Besson, C.; Manach, C.; Demigne, C.; Remesy, C. Quercetin, but not Its glycosides, Iis absorbed from the rat stomach. J. Agric. Food Chem. 2002, 50, 618–621. [Google Scholar] [CrossRef] [PubMed]
- Carriere, F.; Barrowman, J.A.; Verger, R.; René, L. Secretion and contribution to lipolysis of gastric and pancreatic lipases during a test meal in humans. Gastroenterology 1993, 105, 876–888. [Google Scholar] [CrossRef]
- Hernell, O.; Staggers, J.E.; Carey, M.C. Physical-chemical behaviour of dietary and biliary lipids during intestinal digestion and absorption 2. Phase analysis and aggregation states of luminal lipids during fat digestion in healthy adult human beings. Biochemistry 1990, 29, 2041–2056. [Google Scholar] [CrossRef] [PubMed]
- Artursson, P. Epithelial trasport of drugs in cell culture. I: A model for studying the passive diffusion of drugs over intestinal absorbtive (Caco-2) cells. J. Pharm. Sci. 1990, 79, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Yee, S. In vitro permeability across Caco-2 cells (colonic) can predict in vivo (small intestine) absorption in man—Fact or myth. Pharm. Res. 1997, 14, 763–766. [Google Scholar] [CrossRef] [PubMed]
- Borel, P.; Grolier, P.; Partier, A.; Lafont, H.; Lairon, D.; Azais-Braesco, V. Carotenoids in biological emulsions: Solubility, surface-to-core distribution, and release from lipid droplets. J. Lipid Res. 1996, 37, 250–261. [Google Scholar] [PubMed]
- Nugent, S.G.; Kumar, D.; Rampton, D.S.; Evans, D.F. Intestinal luminal pH in inflammatory bowel disease: Possible determinants and implications for therapy with aminosalicylate and other drugs. Gut 2001, 48, 541–547. [Google Scholar] [CrossRef]
- Li, H.; Zhao, X.; Ma, Y.; Zhai, G.; Li, L.; Lou, H. Enhancement of gastrointestinal absorption of quercetin by solid lipid nanoparticles. J. Control. Release 2009, 133, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Rechner, A. Colonic metabolism of dietary polyphenols. Influence of structure on microbial fermentation products. Free Radic. Biol. Med. 2004, 36, 212–225. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BS:PC | (OA) mM | (PC) mM | (BS) mM | |
|---|---|---|---|---|
| Fed state (FE) | 2:1 | 0.3 | 0.75 | 1.5 |
| Fed state (FE) | 5:1 | 0.3 | 0.3 | 1.5 |
| Fasted state (FA) | 2:1 | 0.03 | 0.15 | 0.3 |
| Fasted state (FA) | 5:1 | 0.03 | 0.06 | 0.3 |
| Condition | Solubility (µM) |
|---|---|
| pH 4 | 20.3 (4.9) |
| pH 7.15 | 33.9 (0.6) |
| pH 7.15 + 1 mM BS | 36.0 (1.1) |
| pH6.1 | 21.6 (1.5) |
| pH 6.1 + 1 mM BS | 28.2 (1.1) |
| Olive oil | 1000 (100) |
| Lipid | FR |
|---|---|
| BS micelles a | 1.26 |
| FE5:1 micelles | 1.17 |
| FE2:1 micelles | 1.15 |
| PC liposomes | 0.95 |
| LysoPC micelles 0.5 mM | 0.86 |
| LysoPC micelles 0.3 mM | 0.84 |
| pH | Micelles/Lipid | λmax (nm) a | % Bound a |
|---|---|---|---|
| 7.15 | FE 2:1 | 376.4 (1.1) | 72.7 (2.1) |
| FE2:1 + LysoPC b | 75.0 (1.5) | ||
| FE 5:1 | 376.4 (0.7) | 52.3 (8.3) | |
| FE5:1 + LysoPC b | 60.0 (1.5) | ||
| FA 2:1 | 377.1 (0.3) | 49.5 (7.1) | |
| FA 5:1 | 377.0 (0.4) | 25.0 (7.7) | |
| PC (0.3 mM) | 374.8 (0.2) | 70.4 (1.7) | |
| LysoPC (0.3 mM) | 374.9 (0.3) | 71.7 (1.3) | |
| BS | 377.9 (0.1) | ||
| 6.1 | PC (0.3 mM) | 372.9 (0.2) | 82.7 0.9 |
| LysoPC (0.3 mM) | 372.7 (0.1) | 76.8 (0.7) | |
| BS | 376.2 (0.1) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rich, G.T.; Buchweitz, M.; Winterbone, M.S.; Kroon, P.A.; Wilde, P.J. Towards an Understanding of the Low Bioavailability of Quercetin: A Study of Its Interaction with Intestinal Lipids. Nutrients 2017, 9, 111. https://doi.org/10.3390/nu9020111
Rich GT, Buchweitz M, Winterbone MS, Kroon PA, Wilde PJ. Towards an Understanding of the Low Bioavailability of Quercetin: A Study of Its Interaction with Intestinal Lipids. Nutrients. 2017; 9(2):111. https://doi.org/10.3390/nu9020111
Chicago/Turabian StyleRich, Gillian T., Maria Buchweitz, Mark S. Winterbone, Paul A. Kroon, and Peter J. Wilde. 2017. "Towards an Understanding of the Low Bioavailability of Quercetin: A Study of Its Interaction with Intestinal Lipids" Nutrients 9, no. 2: 111. https://doi.org/10.3390/nu9020111
APA StyleRich, G. T., Buchweitz, M., Winterbone, M. S., Kroon, P. A., & Wilde, P. J. (2017). Towards an Understanding of the Low Bioavailability of Quercetin: A Study of Its Interaction with Intestinal Lipids. Nutrients, 9(2), 111. https://doi.org/10.3390/nu9020111