Lychee Seed Saponins Improve Cognitive Function and Prevent Neuronal Injury via Inhibiting Neuronal Apoptosis in a Rat Model of Alzheimer’s Disease

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Information
2.2. Plant Material and Extraction
2.3. Analysis of the Chemical Profile of LSS by UHPLC-SPD Chromatogram
2.4. Experimental Animals
2.5. Morris Water Maze Test
2.6. Establishment of the Rat Model of AD
2.7. Drug Preparation and Administration
2.8. Histopathological Examination of Hippocampal Neurons of Rats
2.9. Apoptosis Analysis of Neurons of Rats by Morphology and TUNEL
2.10. RNA Isolation and Analysis of Caspase-3 mRNA Expression of the Brain Tissues of Rats by Reverse Transcription-Polymerase Chain Reaction
2.11. Cell Culture
2.12. Western Blotting Analysis of the Protein Expressions of Bcl-2 and Bax in PC12 Cells In Vitro
2.13. Statistical Analysis
3. Results
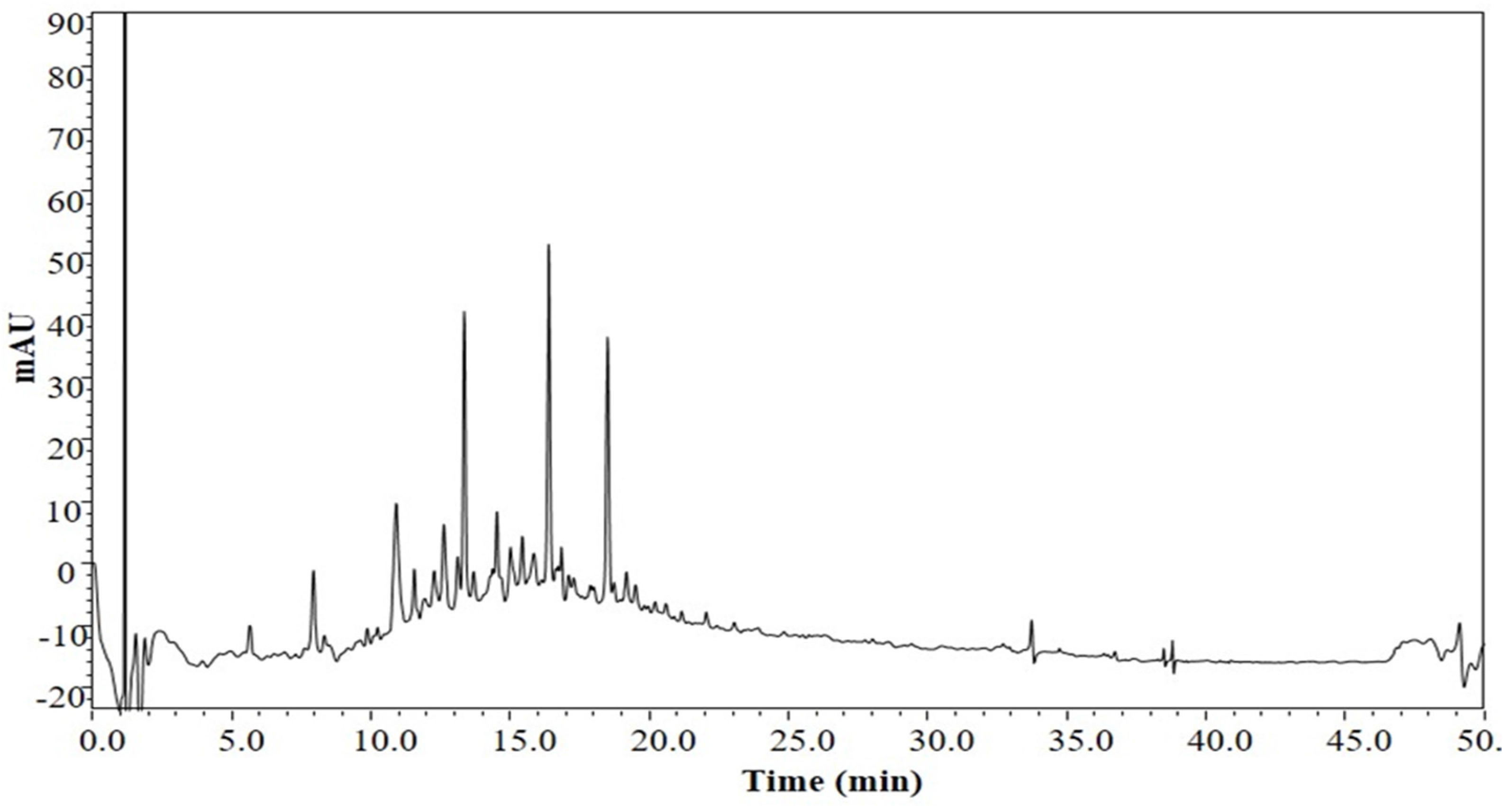
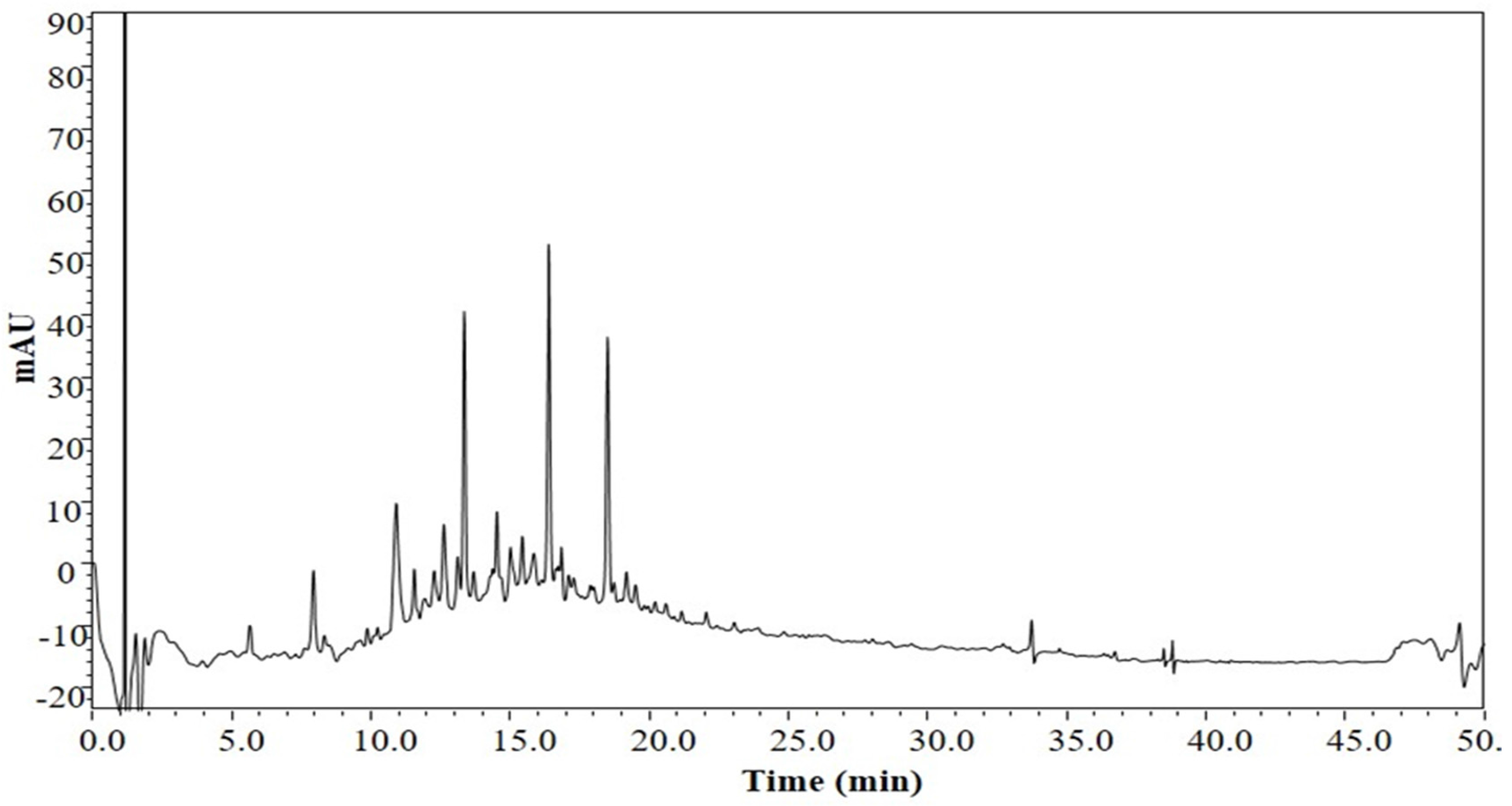
3.1. The Chemical Profile of LSS
3.2. Validation of the Rat Model of AD by Escape Latency Test
3.3. Effect of LSS on the Ability of Spatial Learning and Memory of the Rats with AD
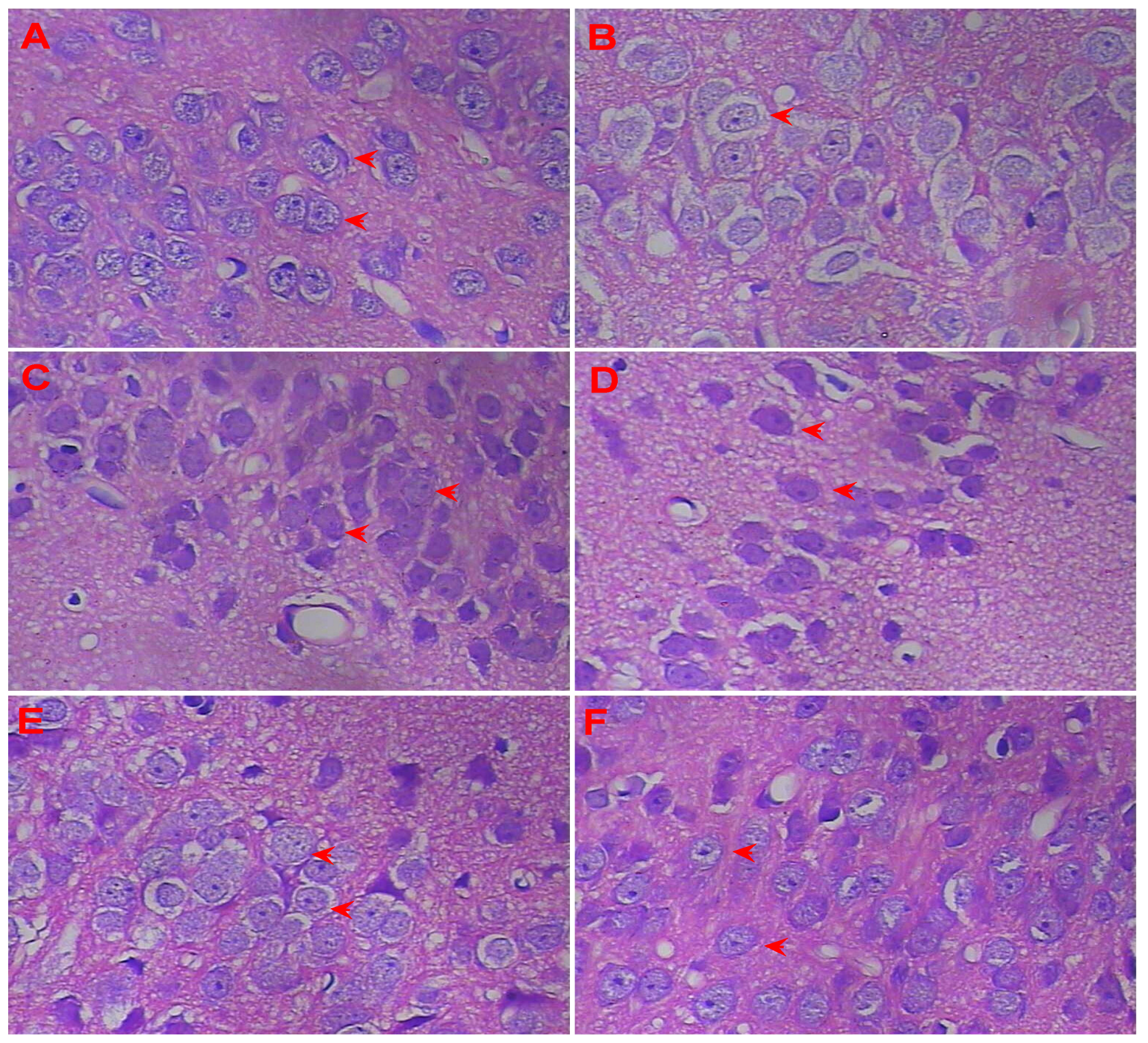
3.4. Effect of LSS on the Neuroprotection in the Rats with AD
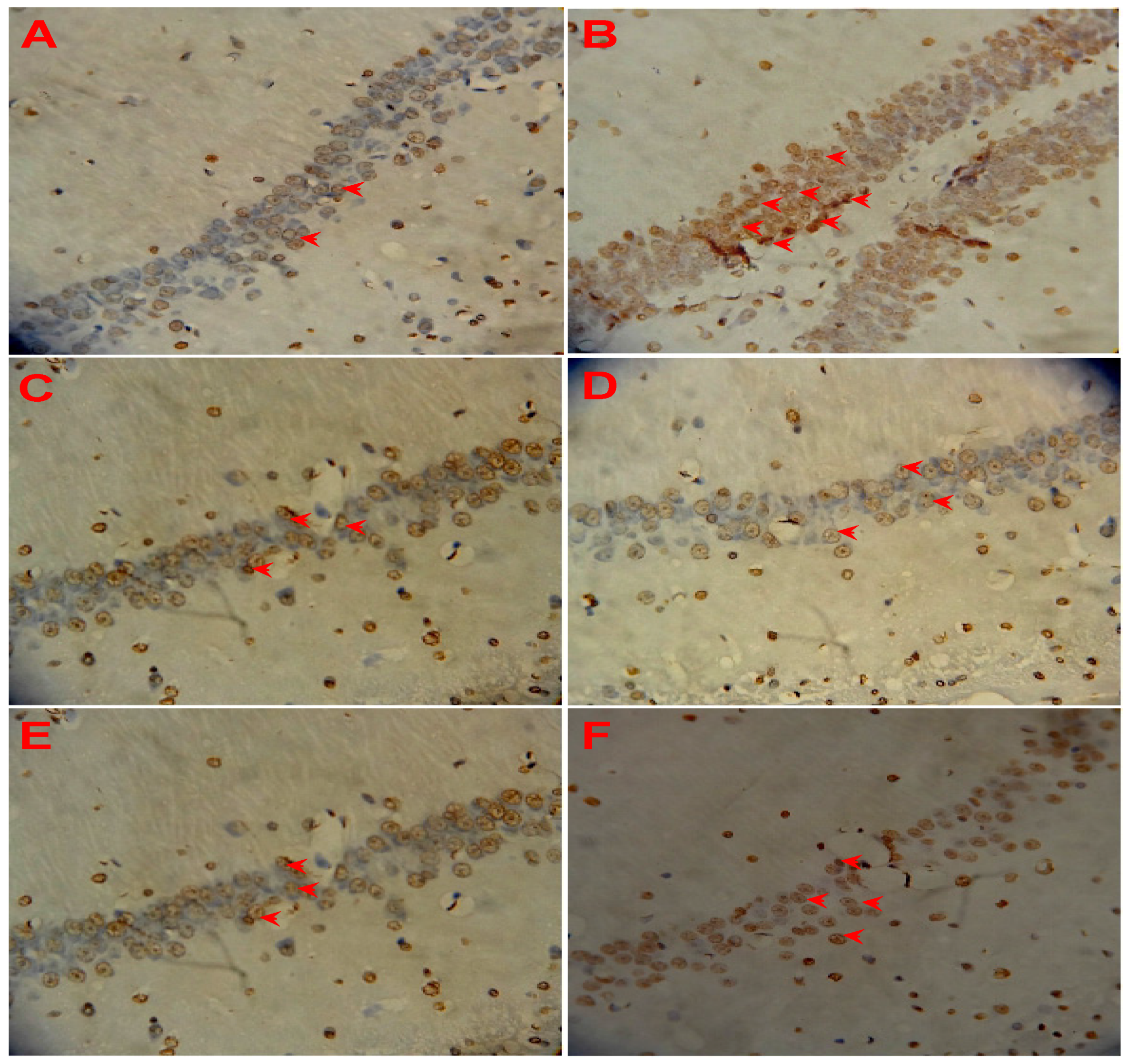
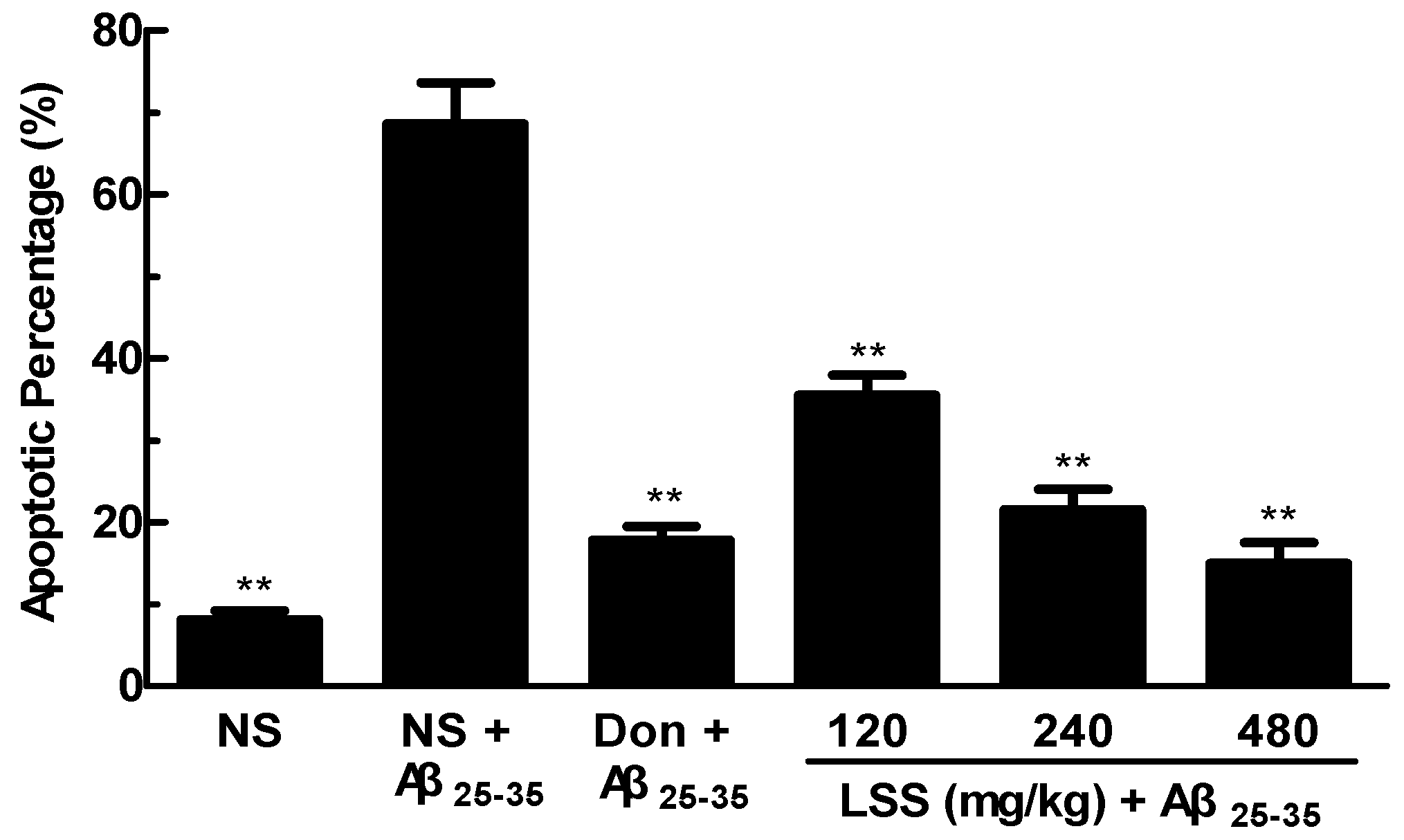
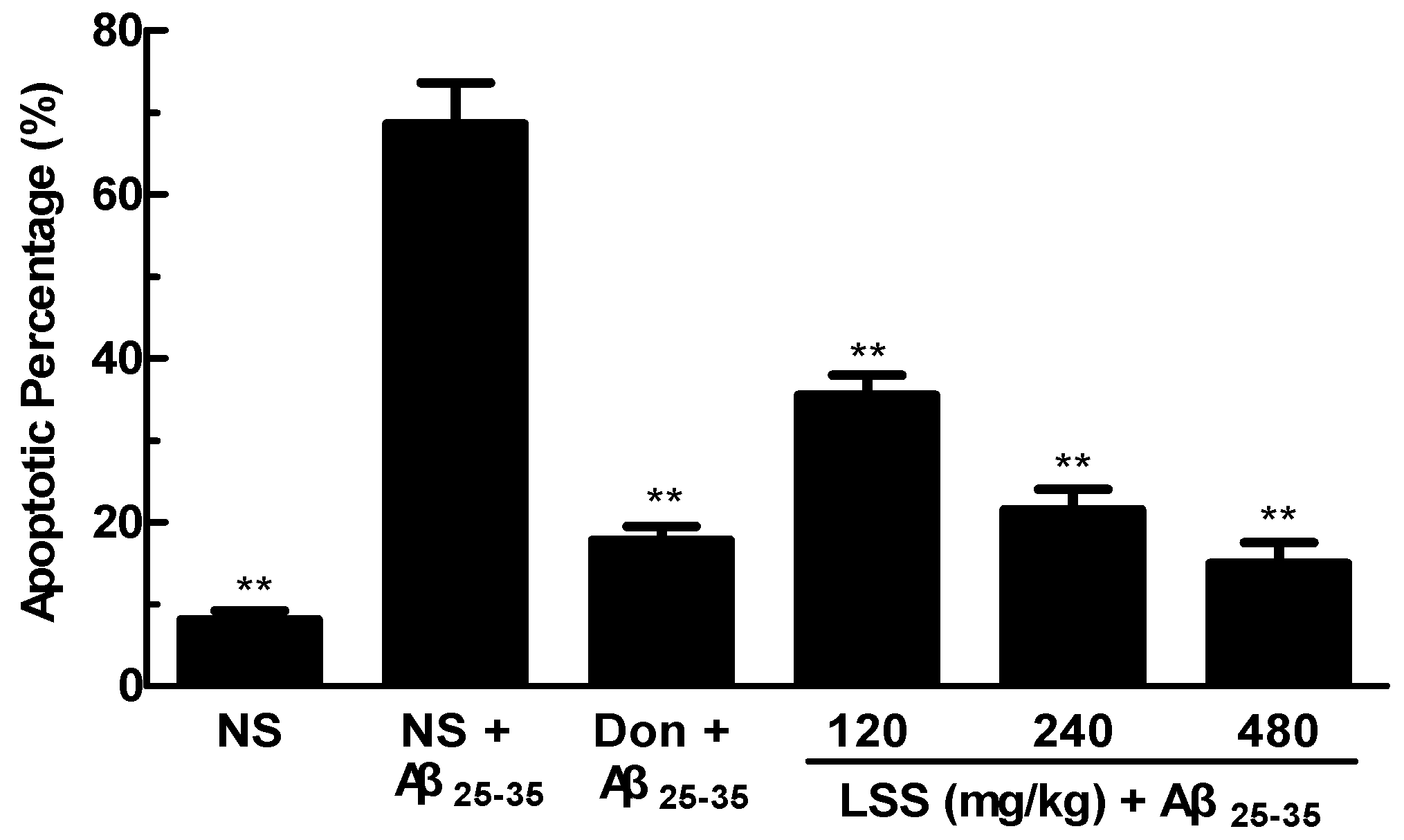
3.5. Effect of LSS on Neuronal Apoptosis in the Rats with AD
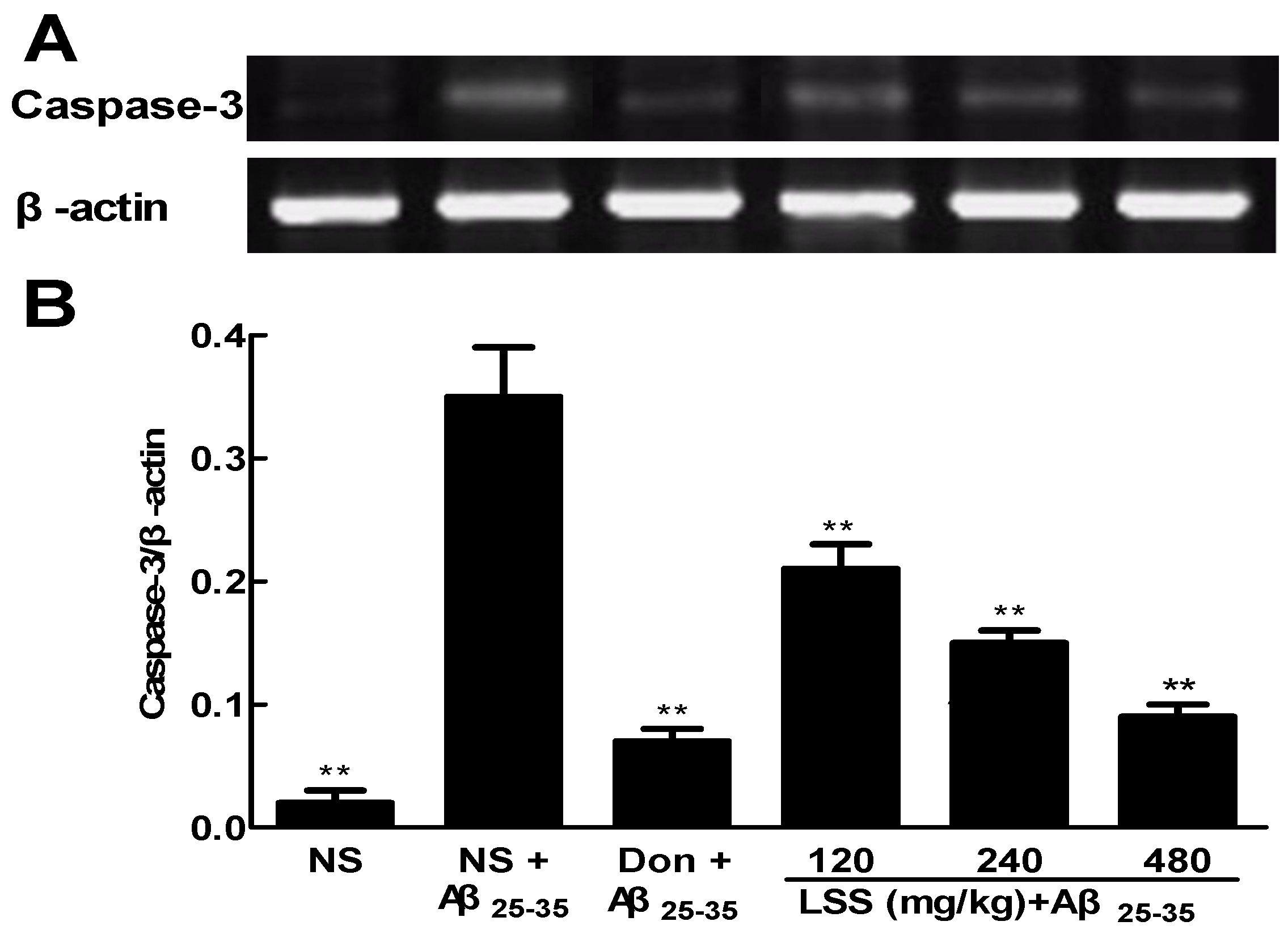
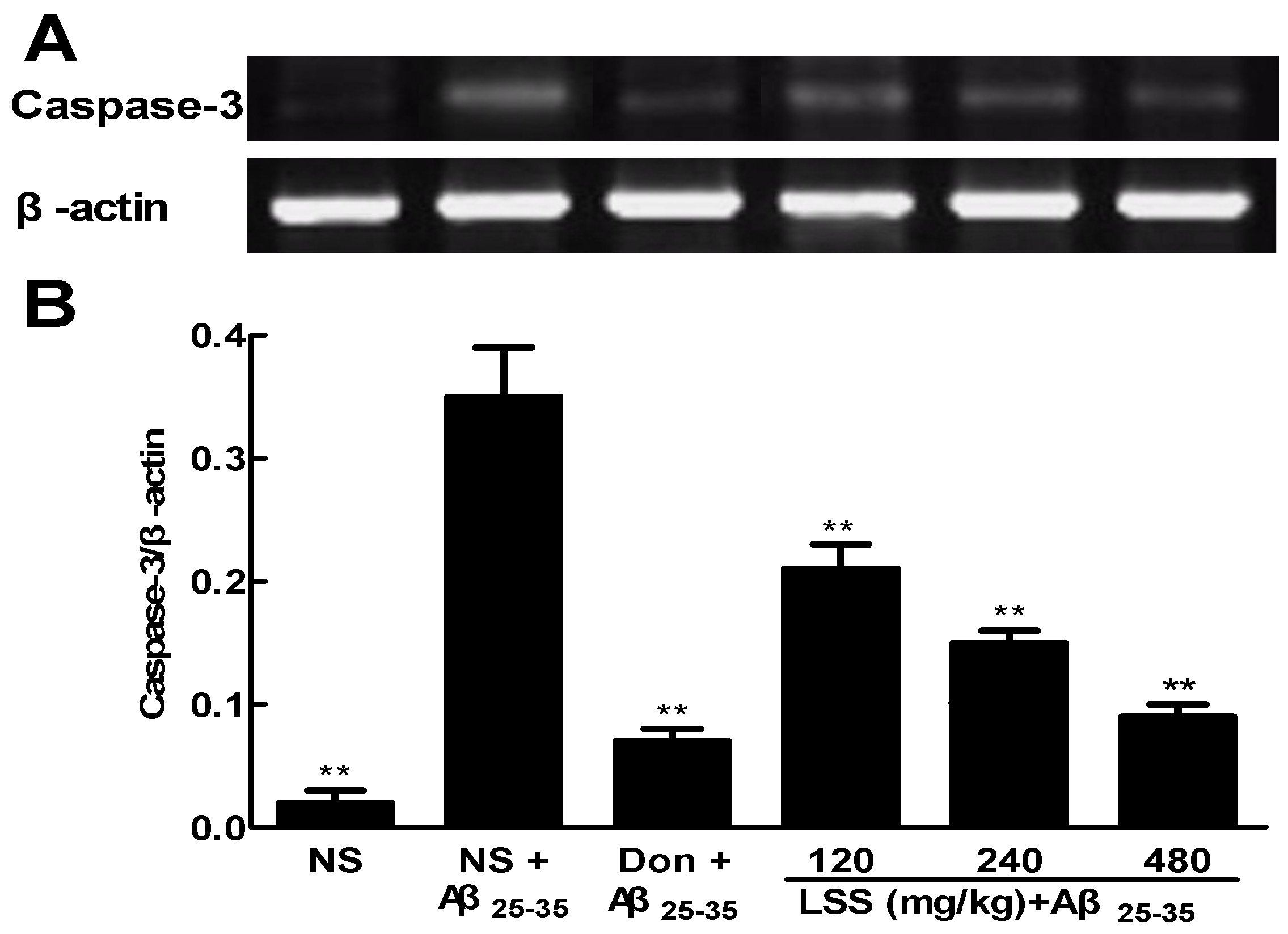
3.6. Effect of LSS on the mRNA Expression of Caspase-3 in the Neuronal Tissues of the Rats with AD
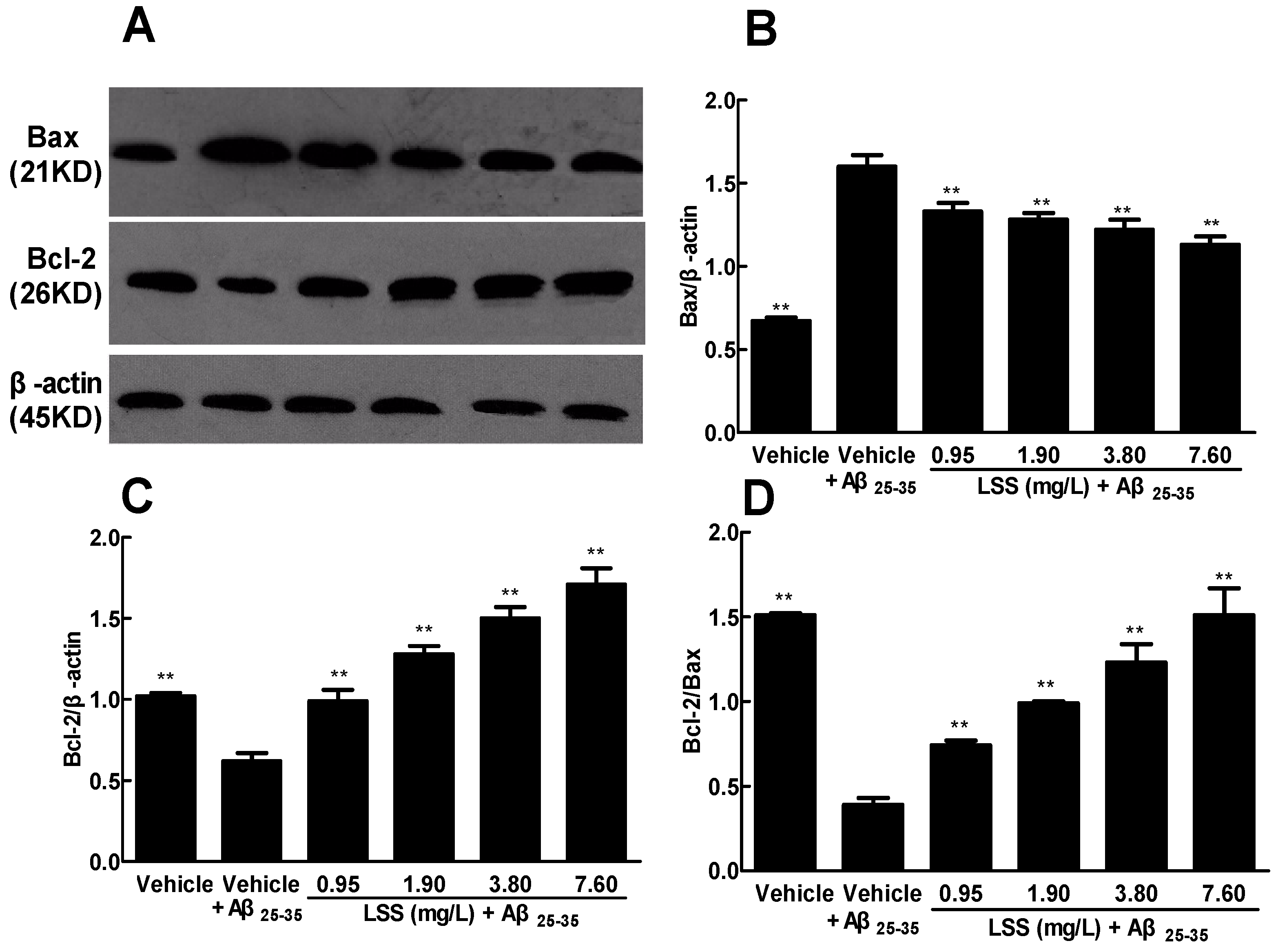
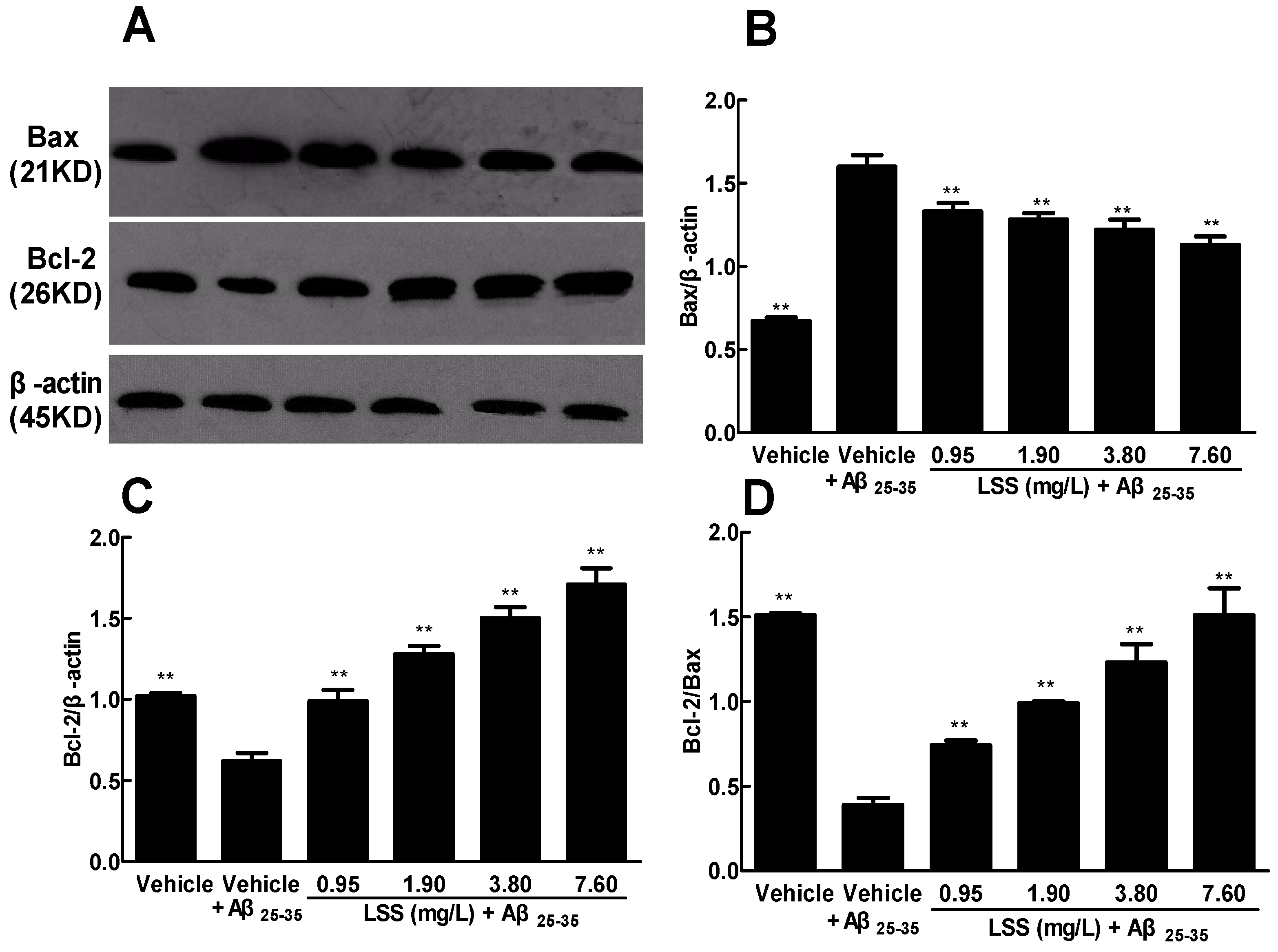
3.7. Effect of LSS on the Protein Expressions of Bax and Bcl-2 in the PC12 Cells Treated with Aβ25–35
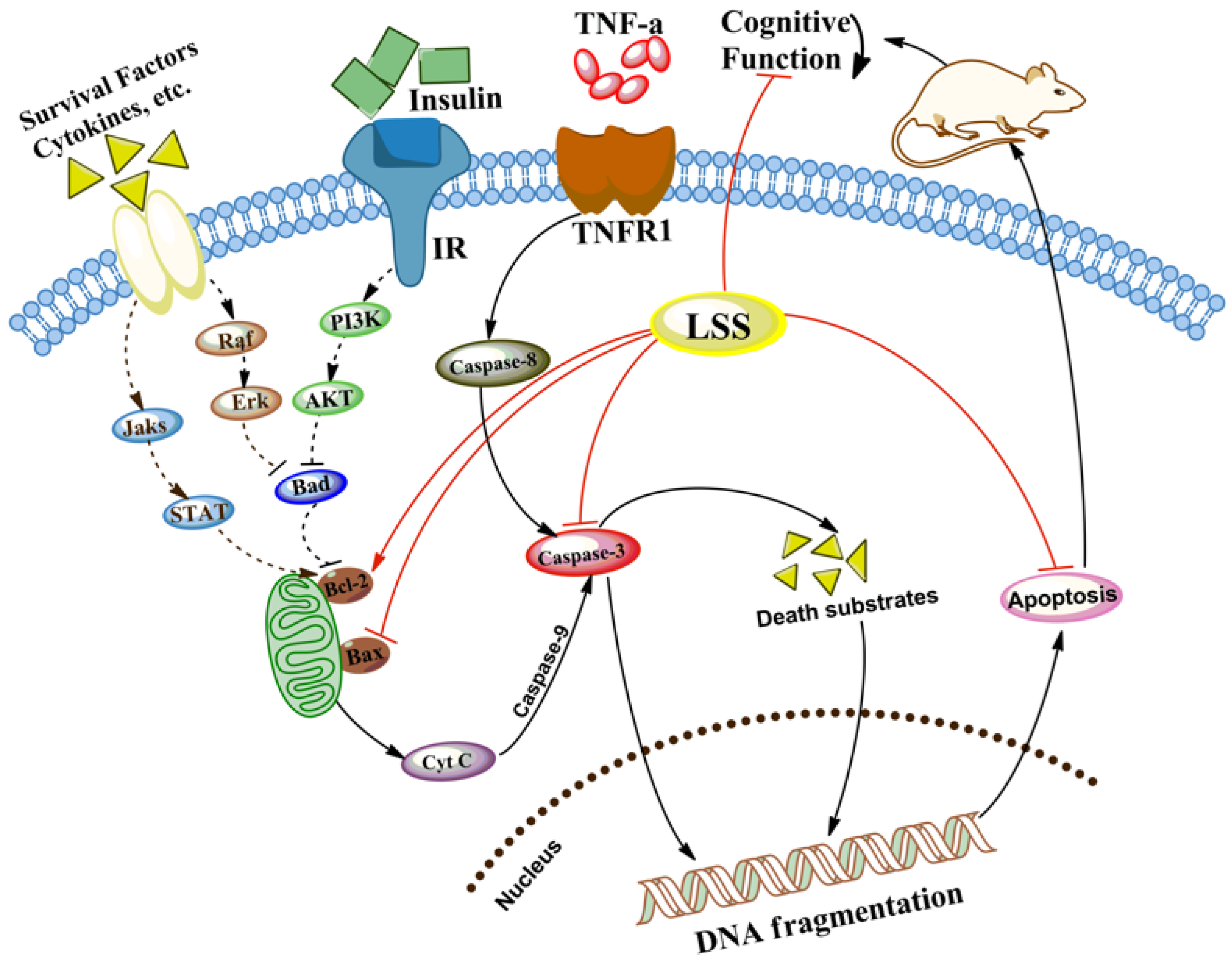
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cairns, N.J. Alzheimer’s disease: Neurodegeneration. In Encyclopedia of Neuroscience; Squire, L.R., Ed.; Academic Press: Oxford, UK, 2009; pp. 275–281. [Google Scholar]
- Kimura, N. Diabetes mellitus induces Alzheimer’s disease pathology: Histopathological evidence from animal models. Int. J. Mol. Sci. 2016, 17, 503. [Google Scholar] [CrossRef] [PubMed]
- Loo, D.T.; Copani, A.; Pike, C.J.; Whittemore, E.R.; Walencewicz, A.J.; Cotman, C.W. Apoptosis is induced by beta-amyloid in cultured central nervous system neurons. Proc. Natl. Acad. Sci. USA 1993, 90, 7951–7955. [Google Scholar] [CrossRef] [PubMed]
- Forloni, G.; Chiesa, R.; Smiroldo, S.; Verga, L.; Salmona, M.; Tagliavini, F.; Angeretti, N. Apoptosis mediated neurotoxicity induced by chronic application of beta amyloid fragment 25–35. Neuroreport 1993, 4, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Crews, L.; Masliah, E. Molecular mechanisms of neurodegeneration in Alzheimer’s disease. Hum. Mol. Gen. 2010, 19, R12–R20. [Google Scholar] [CrossRef] [PubMed]
- Obulesu, M.; Lakshmi, M.J. Apoptosis in Alzheimer’s disease: An understanding of the physiology, pathology and therapeutic avenues. Neurochem. Res. 2014, 39, 2301–2312. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.; Xiong, L.L.; Zhang, R.P.; Zheng, H.; Wang, L.; Qian, Z.Y.; Zhang, P.; Chen, Z.W.; Gao, F.B.; Wang, T.H. Injection of Aβ1–40 into hippocampus induced cognitive lesion associated with neuronal apoptosis and multiple gene expressions in the tree shrew. Apoptosis 2016, 21, 621–640. [Google Scholar] [CrossRef] [PubMed]
- Paquet, C.; Dumurgier, J.; Hugon, J. Pro-apoptotic kinase levels in cerebrospinal fluid as potential future biomarkers in Alzheimer’s disease. Front. Neurol. 2015, 6, 168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, L.L.; Ren, R.J.; Dammer, E.B.; Zhang, Y.F.; Huang, Y.; Chen, S.D.; Wang, G. MicroRNA-146a represses LRP2 translation and leads to cell apoptosis in Alzheimer’s disease. FEBS Lett. 2016, 590, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Porter, A.G.; Jänicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Srivastav, S.; Yadav, A.K.; Srikrishna, S.; Perry, G. Overview of Alzheimer’s disease and some therapeutic approaches targeting A by using several synthetic and herbal compounds. Oxid. Med. Cell. Longev. 2016, 2016, 7361613. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Kim, J.E.; Nam, S.H.; Goo, J.S.; Choi, S.I.; Choi, Y.H.; Bae, C.J.; Woo, J.M.; Cho, J.S.; Hwang, D.Y. Differential regulation of the biosynthesis of glucose transporters by the PI3K and MAPK pathways of insulin signaling by treatment with novel compounds from Liriope platyphylla. Int. J. Mol. Med. 2011, 27, 319–327. [Google Scholar] [PubMed]
- Shen, B.; Kwan, H.Y.; Ma, X.; Wong, C.O.; Du, J.; Huang, Y.; Yao, X.Q. cAMP activates TRPC6 channels via the phosphatidylinositol 3-kinase (PI3K)-protein kinase B (PKB)-mitogen-activated protein kinase kinase (MEK)-ERK1/2 signaling pathway. J. Biol. Chem. 2011, 86, 19439–19445. [Google Scholar] [CrossRef] [PubMed]
- Pramojanee, S.N.; Phimphilai, M.; Chattipakorn, N.; Chattipakorn, S.C. Possible roles of insulin signaling in osteoblasts. Endocr. Res. 2014, 39, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Heras-Sandoval, D.; Ferrera, P.; Arias, C. Amyloid-β protein modulates insulin signaling in presynaptic terminals. Neurochem. Res. 2012, 37, 1879–1885. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Ma, D.; Wang, Y.; Jiang, T.; Hu, S.; Zhang, M.; Yu, X.; Gong, C.X. Intranasal insulin ameliorates tau hyperphosphorylation in a rat model of type 2 diabetes. J. Alzheimers Dis. 2013, 33, 329–338. [Google Scholar]
- Li, J.W.; Li, L.L.; Chang, L.L.; Wang, Z.Y.; Xu, Y. Stem cell factor protects against neuronal apoptosis by activating AKT/ERK in diabetic mice. Braz. J. Med. Biol. Res. 2009, 42, 1044–1049. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Wang, J.; Zhao, M.; Liu, Y.; Wang, W.; Jiang, Y.M. Identification of polysaccharides from pericarp tissues of litchi (Litchi chinensis Sonn.) fruit in relation to their antioxidant activities. Carbohydr. Res. 2006, 341, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Sintes, R.; Hernández, F.; Lucas, J.J.; Avila, J. GSK-3 mouse models to study neuronal apoptosis and neurodegeneration. Front. Mol. Neurosci. 2011, 4, 45. [Google Scholar] [CrossRef] [PubMed]
- Cerioni, L.; Cantoni, O. Assessing Bad sub-cellular localization under conditions associated with prevention or promotion of mitochondrial permeability transition-dependent toxicity. Methods Mol. Biol. 2010, 648, 291–301. [Google Scholar] [PubMed]
- Lee, J.H.; Jeong, S.K.; Kim, B.C.; Park, K.W.; Dash, A. Donepezil across the spectrum of Alzheimer’s disease: Dose optimization and clinical relevance. Acta Neurol. Scand. 2015, 131, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Querfurth, H.W.; LaFerla, F.M. Alzheimer’s disease. N. Engl. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Godyń, J.; Jończyk, J.; Panek, D.; Malawska, B. Therapeutic strategies for Alzheimer’s disease in clinical trials. Pharmacol. Rep. 2016, 68, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Anekonda, T.S.; Reddy, P.H. Can herbs provide a new generation of drugs for treating Alzheimer’s disease? Brain Res. Rev. 2005, 50, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Farver, D. The use of “natural products” in clinical medicine. South Dakota J. Med. 1996, 49, 129–130. [Google Scholar]
- Kim, J.; Lee, H.J.; Lee, K.W. Naturally occurring phytochemicals for the prevention of Alzheimer’s disease. J. Neurochem. 2010, 112, 1415–1430. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.P. Beyond antioxidants: The cellular and molecular interactions of flavonoids and how these underpin their actions on the brain. Proc. Nutr. Soc. 2010, 69, 244–260. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.J.; Guo, J.W.; Pan, J.Q. Pharmacologic and pharmacodynamic effects of effective element of Litchi and Litchi seed. Pharm. Today 2009, 5, 7–9. [Google Scholar]
- Xiao, Z.J.; Guo, J.W.; Xu, F. Effect of litchi saponin and litchi flavones on insulin resistance in HepG2 cells. J. Pharm. Pract. 2015, 4, 316–318. [Google Scholar]
- Zhang, Y.M.; Yuan, H.; Tian, J.X.; Shen, L.; Yong, Y.H. Effects of saponin of litchi seed on gluconeogenesis and metabolism of blood lipid in mice. J. Hangzhou Teach. Coll. 2005, 6, 435–436. [Google Scholar]
- Xiao, L.Y.; Pan, Z.J.; Rao, W.N. The research of protective effect of Litchi seed of experimental liver injury in mice. Chin. J. Trad. Chin. Med. Pharm. 2005, 20, 42–44. [Google Scholar]
- Zhang, J.; Zhang, C. Research progress on the antineoplastic pharmacological effects and mechanisms of Litchi seeds. Chin. Med. 2015, 6, 20–26. [Google Scholar] [CrossRef]
- Li, W.; Zhu, Y.T.; Huang, Z.Y.; He, J.J.; Pei, J.; Song, J.P. Experimental studies on anti-fluvirus effect of Litchi seed in vivo. Chin. J. Ethnomed. Ethnopharm. 2011, 18, 34–36. [Google Scholar]
- Zhao, H.X.; Guo, K.; Cui, Y.D.; Wu, X.G.; Shang, Y.Z. Effect of Scutellaria. barbata flavonoids on abnormal changes of Bcl-2, Bax, Bcl-xL and Bak protein expression in mitochondrial membrane induced by composite Aβ(25–35). Chin. J. Pathophysiol. 2014, 30, 2262–2266. [Google Scholar]
- Ye, H.M.; Zhong, C.Y.; Huang, M.X.; Wang, C.Y.; Fang, X.; Chen, X.Y.; Lv, J.H. Effect of litchi seed aqueous extracts on learning and memory obstacles induced by d-galactose in mice and its mechanism. J. Chin. Med. Mater. 2013, 36, 438–440. [Google Scholar]
- Yang, Y.J.; Liang, B.M. Determination of anti-diabete saponins from Litchi chinensis sonn. Guangdong Pharm. J. 2004, 14, 13–15. [Google Scholar]
- Li, N.; Liu, B.; Dluzen, D.E.; Jin, Y. Protective effects of ginsenoside Rg2 against glutamate-induced neurotoxicity in PC12 cells. J. Ethnopharmacol. 2007, 111, 458–463. [Google Scholar] [CrossRef] [PubMed]
- Barnhart, C.D.; Yang, D.; Lein, P.J. Using the Morris water maze to assess spatial learning and memory in weanling mice. PLoS ONE 2015, 10, e0124521. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Guo, H.; Li, C.; Wang, D.; Wu, J.; Wang, C.; Xu, J.; Qin, R.A. Cognitive improvement of compound danshen in an Aβ25–35 peptide-induced rat model of Alzheimer’s disease. BMC Complement. Altern. Med. 2015, 15, 382. [Google Scholar] [CrossRef] [PubMed]
- Mehdizadeh, R.; Parizadeh, M.R.; Khooei, A.R.; Mehri, S.; Hosseinzadeh, H. Cardioprotective effect of saffron extract and safranal in isoproterenol-induced myocardial infarction in wistar rats. Iran. J. Basic Med. Sci. 2013, 16, 56–63. [Google Scholar] [PubMed]
- Fang, F.; Wang, A.P.; Yang, S.F. Antitumor activity of a novel recombinant mutant human tumor necrosis factor-related apoptosis-inducing ligand. Acta Pharmacol. Sin. 2005, 26, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.Q.; Bai, L.; Cai, H.; Li, J.L. The effect of eye-acupuncture on the expression of caspase-3 mRNA and caspase-8 mRNA in the ischemic penumbra. J. Clin. Acupunct. Moxib. 2010, 26, 51–54. [Google Scholar]
- Chen, X.; Sun, J.X.; Cao, C.; Anatomy, D.O. Effects of Aβ1–42 oligomer on cerebral cortical Bcl-2 and Caspase-3 expressions in AD rats. Acta Univ. Med. Anhui 2014, 49, 291–294. [Google Scholar]
- Sun, R.; Wang, K.; Wu, D.; Li, X.; Ou, Y. Protective effect of paeoniflorin against glutamate-induced neurotoxicity in PC12 cells via Bcl-2/Bax signal pathway. Folia Neuropathol. 2012, 50, 270–276. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Ji, O.; Li, J.Q. The pharmacological effect of Puerarin extract on learning and memory ability of rats with Alzheimer disease. Pharm. Clin. Chin. Mat. Med. 2014, 5, 39–42. [Google Scholar]
- Putcha, G.V.; Harris, C.A.; Moulder, K.L.; Easton, R.M.; Thompson, C.B.; Johnson, E.M. Intrinsic and extrinsic pathway signaling during neuronal apoptosis: Lessons from the analysis of mutant mice. J. Cell Biol. 2002, 157, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Danial, N.N.; Gramm, C.F.; Scorrano, L.; Zhang, C.Y.; Krauss, S.; Ranger, A.M.; Datta, S.R.; Greenberg, M.E.; Licklider, L.J.; Lowell, B.B.; et al. BAD and glucokinase reside in a mitochondrial complex that integrates glycolysis and apoptosis. Nature 2003, 424, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Renault, T.T.; Teijido, O.; Antonsson, B.; Dejean, L.M.; Manon, S. Regulation of Bax mitochondrial localization by Bcl-2 and Bcl-x (L): Keep your friends close but your enemies closer. Int. J. Biochem. Cell Biol. 2013, 45, 64–67. [Google Scholar] [CrossRef] [PubMed]
- D’Amelio, M.; Sheng, M.; Cecconi, F. Caspase-3 in the central nervous system: Beyond apoptosis. Trends Neurosci. 2012, 35, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Brentnall, M.; Rodriguez-Menocal, L.; De Guevara, R.L.; Cepero, E.; Boise, L.H. Caspase-9, caspase-3 and caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell Biol. 2013, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Shiri, R.; Yari, F.; Ahmadinejad, M.; Vaeli, S.; Tabatabei, M.R. The caspase-3 inhibitor (peptide Z-DEVD-FMK) affects the survival and function of platelets in platelet concentrate during storage. Blood Res. 2014, 49, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Kitazumi, I.; Maseki, Y.; Nomura, Y.; Shimanuki, A.; Sugita, Y.; Tsukahara, M. Okadaic acid induces DNA fragmentation via caspase-3-dependent and caspase-3-independent pathways in Chinese hamster ovary (CHO)-K1 cells. FEBS J. 2010, 277, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Momeni, H.R.; Soleimani-Mehranjani, M.; Shariatzadeh, M.A.; Haddadi, M. Caspase-mediated apoptosis in sensory neurons of cultured dorsal root Ganglia in adult mouse. Cell J. 2013, 15, 212–217. [Google Scholar] [PubMed]
- Jarero-Basulto, J.J.; Luna-Muñoz, J.; Mena, R.; Ktistofikova, Z.; Ripova, D.; Perry, G.; Binder, L.; Garcia-Sierra, F. Proteolytic cleavage of polymeric tau protein by caspase-3: Implications for Alzheimer disease. J. Neuropathol. Exp. Neurol. 2013, 72, 1145–1161. [Google Scholar] [CrossRef] [PubMed]
 LSS 480 mg/kg; (B) Number of times across the platform; (C) platform quadrant dwell time; and (D) percentage of the total distance run in the platform quadrant. There were 10 rats used for each experimental group and expressed as the mean ± SD. ** p < 0.01 vs. the AD rats treated with NS (control).
LSS 480 mg/kg; (B) Number of times across the platform; (C) platform quadrant dwell time; and (D) percentage of the total distance run in the platform quadrant. There were 10 rats used for each experimental group and expressed as the mean ± SD. ** p < 0.01 vs. the AD rats treated with NS (control).
LSS 480 mg/kg; (B) Number of times across the platform; (C) platform quadrant dwell time; and (D) percentage of the total distance run in the platform quadrant. There were 10 rats used for each experimental group and expressed as the mean ± SD. ** p < 0.01 vs. the AD rats treated with NS (control).
LSS 480 mg/kg; (B) Number of times across the platform; (C) platform quadrant dwell time; and (D) percentage of the total distance run in the platform quadrant. There were 10 rats used for each experimental group and expressed as the mean ± SD. ** p < 0.01 vs. the AD rats treated with NS (control).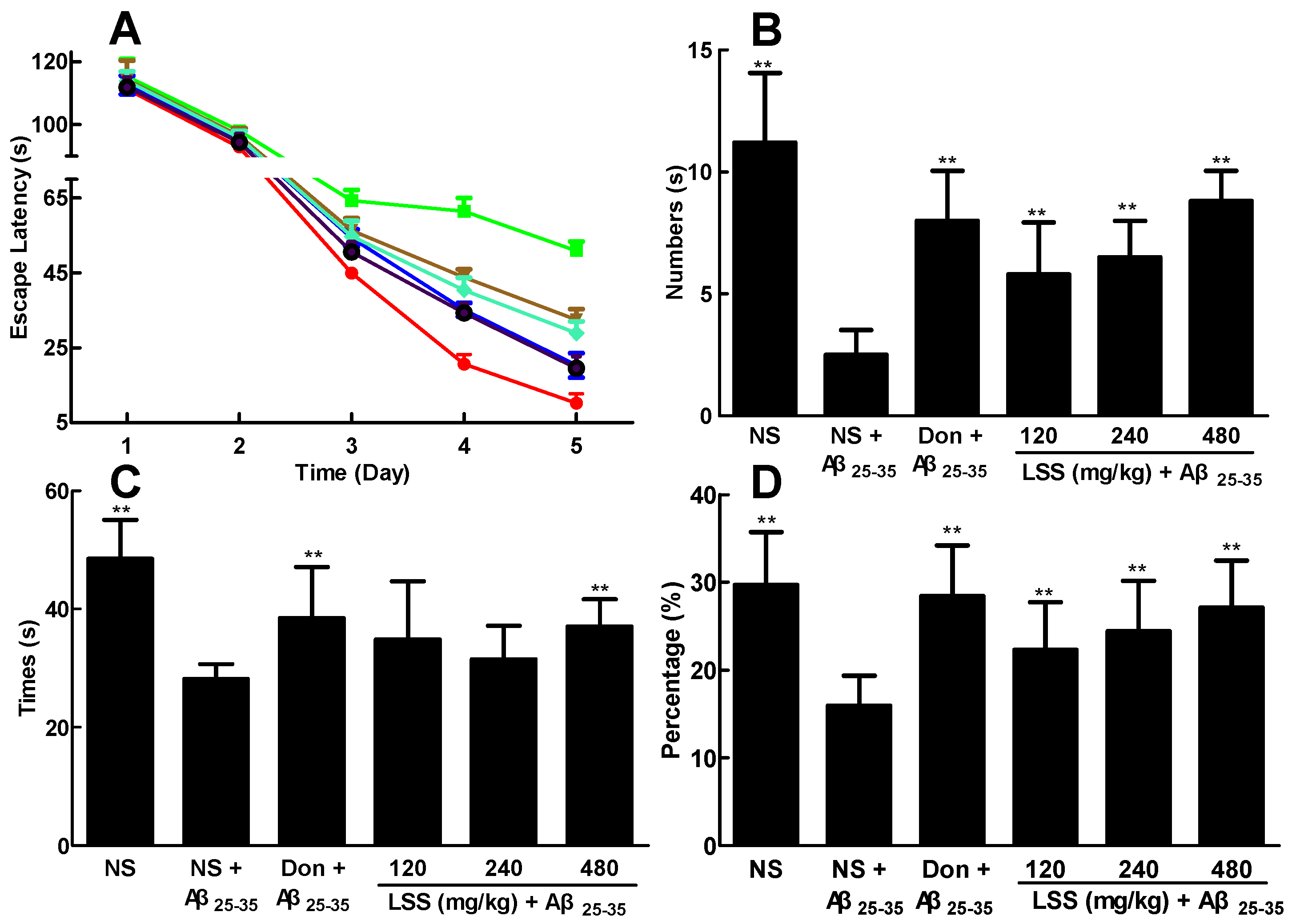

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mRNA | Primer Sequence | PCR Target (bp) | Cycle |
|---|---|---|---|
| Caspase-3 | F: 5′-GAACGATCGGACCTGTGG-3′ R: 5′-GGGTGCGGTAGAGTAAGC-3′ | 218 | 30 |
| β-actin | F: 5′-CTGGAAGGTGGACAGTGAG-3′ R: 5′-GAGGGAAATCGTGCGTGAC-3′ | 445 | 30 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Wu, J.; Yu, C.; Tang, Y.; Liu, J.; Chen, H.; Jin, B.; Mei, Q.; Cao, S.; Qin, D. Lychee Seed Saponins Improve Cognitive Function and Prevent Neuronal Injury via Inhibiting Neuronal Apoptosis in a Rat Model of Alzheimer’s Disease. Nutrients 2017, 9, 105. https://doi.org/10.3390/nu9020105
Wang X, Wu J, Yu C, Tang Y, Liu J, Chen H, Jin B, Mei Q, Cao S, Qin D. Lychee Seed Saponins Improve Cognitive Function and Prevent Neuronal Injury via Inhibiting Neuronal Apoptosis in a Rat Model of Alzheimer’s Disease. Nutrients. 2017; 9(2):105. https://doi.org/10.3390/nu9020105
Chicago/Turabian StyleWang, Xiuling, Jianming Wu, Chonglin Yu, Yong Tang, Jian Liu, Haixia Chen, Bingjin Jin, Qibing Mei, Shousong Cao, and Dalian Qin. 2017. "Lychee Seed Saponins Improve Cognitive Function and Prevent Neuronal Injury via Inhibiting Neuronal Apoptosis in a Rat Model of Alzheimer’s Disease" Nutrients 9, no. 2: 105. https://doi.org/10.3390/nu9020105
APA StyleWang, X., Wu, J., Yu, C., Tang, Y., Liu, J., Chen, H., Jin, B., Mei, Q., Cao, S., & Qin, D. (2017). Lychee Seed Saponins Improve Cognitive Function and Prevent Neuronal Injury via Inhibiting Neuronal Apoptosis in a Rat Model of Alzheimer’s Disease. Nutrients, 9(2), 105. https://doi.org/10.3390/nu9020105