Choline and Working Memory Training Improve Cognitive Deficits Caused by Prenatal Exposure to Ethanol

Abstract
1. Introduction
2. Methods
3. Statistical Analysis
4. Results
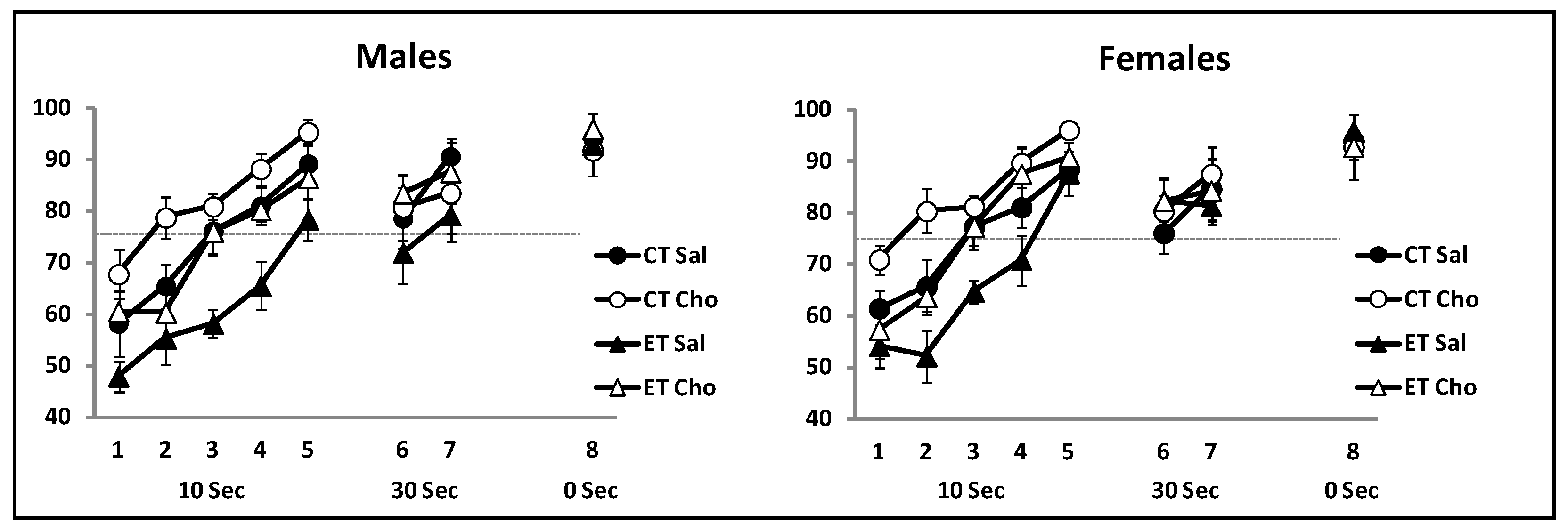
4.1. Delayed Non-Matching to Place
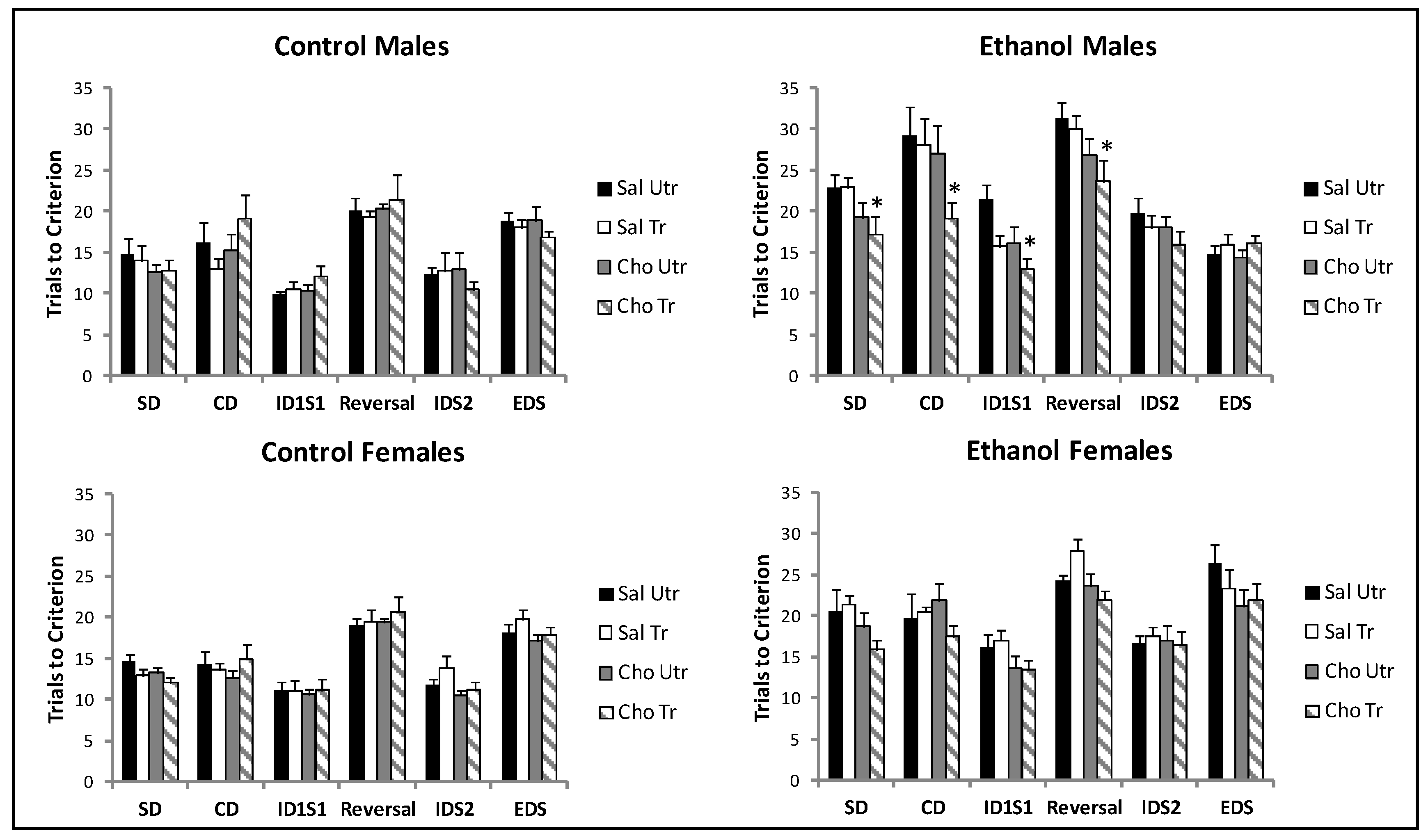
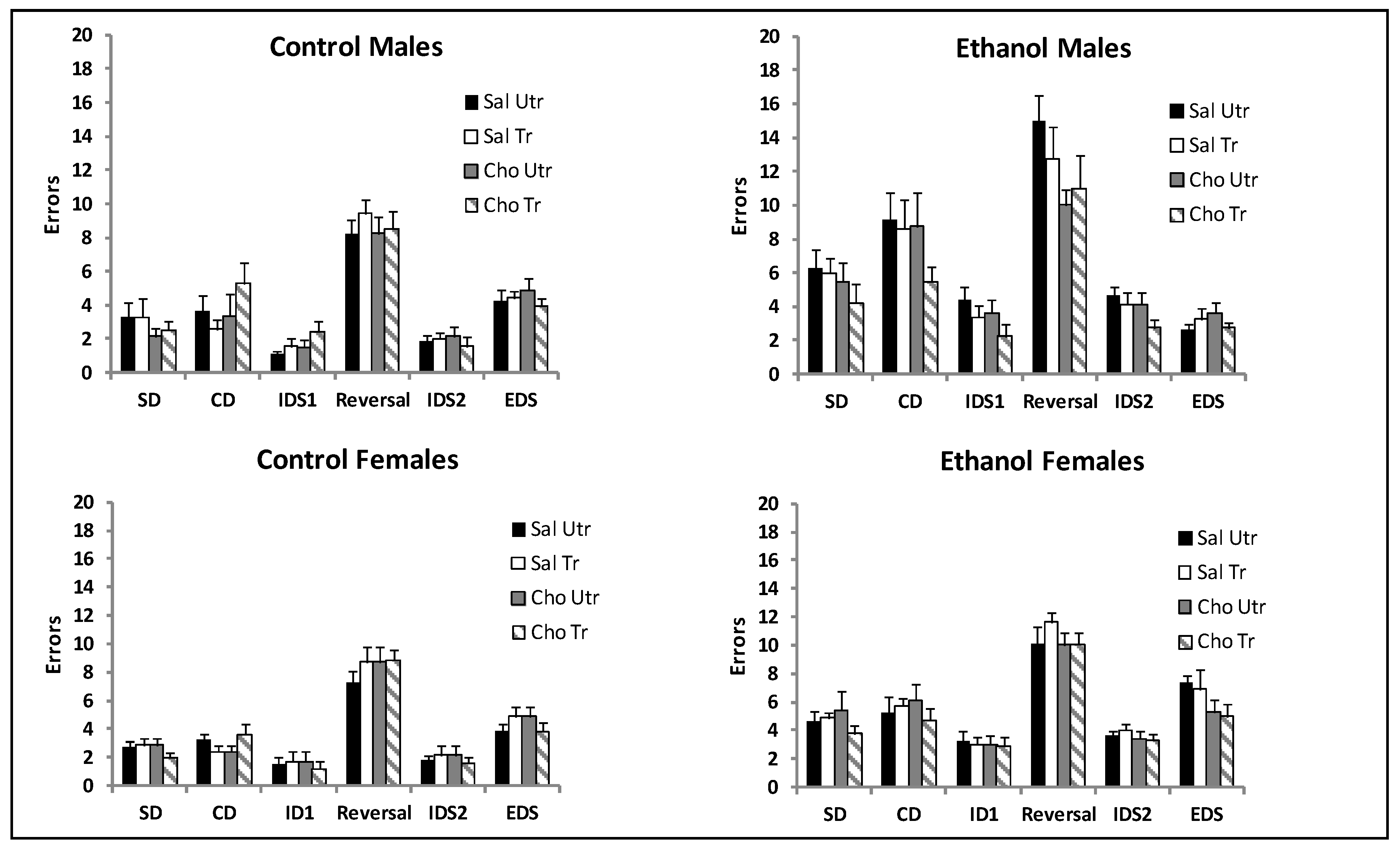
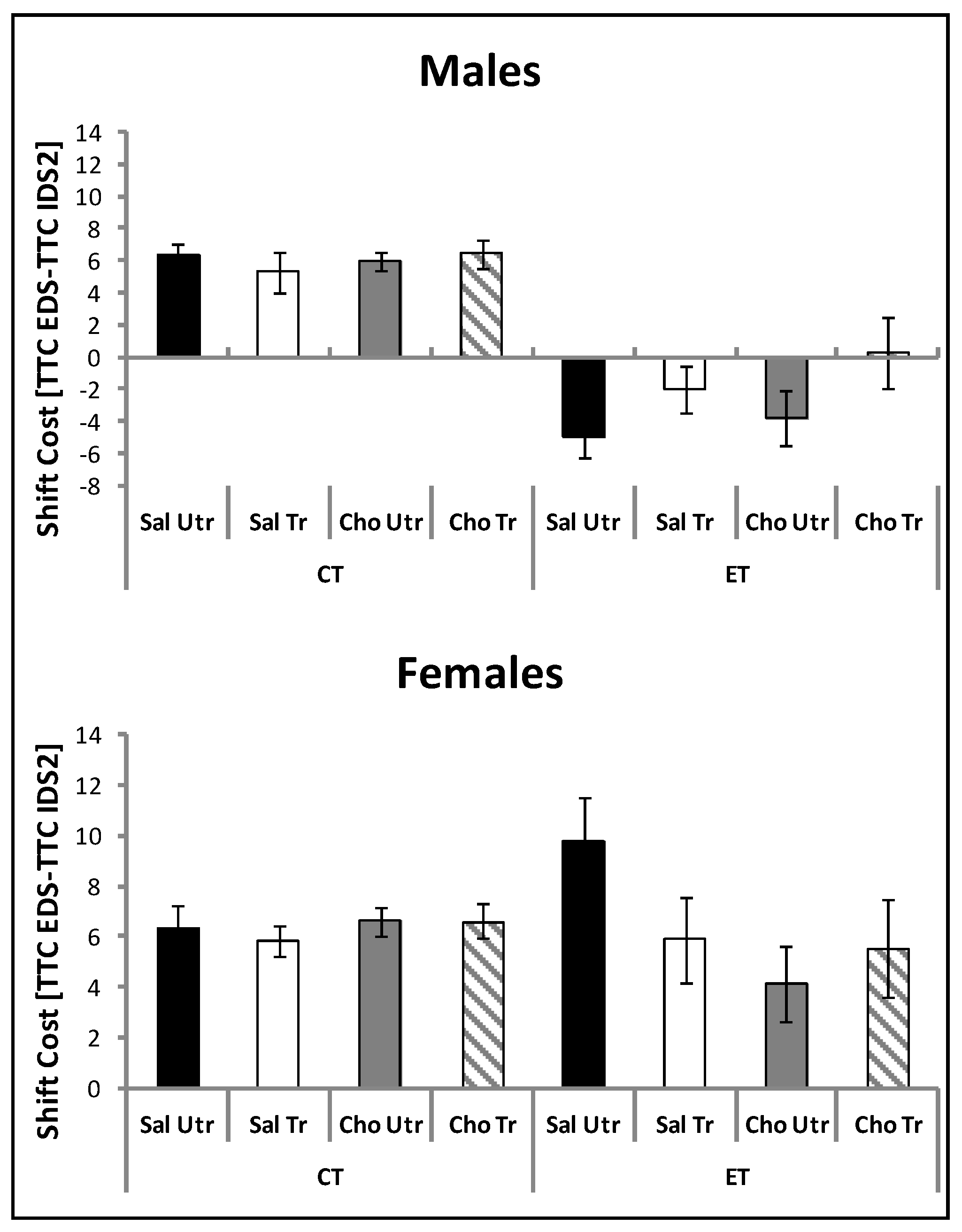
4.2. Attentional Set Shifting and Reversal Learning
5. Discussion
6. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- May, P.A.; Gossage, J.P.; Kalberg, W.O.; Robinson, L.K.; Buckley, D.; Manning, M. Prevalence and epidemiologic characteristics of FASD from various research methods with an emphasis on recent in-school studies. Dev. Disabil. Res. Rev. 2009, 15, 176–192. [Google Scholar] [CrossRef] [PubMed]
- May, P.A.; Blankenship, J.; Marais, A.S.; Gossage, J.P.; Kalberg, W.O.; Barnard, R. Approaching the prevalence of the full spectrum of fetal alcohol spectrum disorders in a South African population-based study. Alcohol. Clin. Exp. Res. 2013, 37, 818–830. [Google Scholar] [CrossRef] [PubMed]
- Connor, P.D.; Sampson, P.D.; Bookstein, F.L.; Barr, H.M.; Streissguth, A.P. Direct and indirect effects of prenatal alcohol damage on executive function. Dev. Neuropsychol. 2000, 18, 331–354. [Google Scholar] [CrossRef] [PubMed]
- Kodituwakku, P.W.; May, P.A.; Clericuzio, C.L.; Weers, D. Emotion-related learning in individuals prenatally exposed to alcohol: An investigation of the relation between set shifting, extinction of responses, and behavior. Neuropsychologia 2001, 39, 699–708. [Google Scholar] [CrossRef]
- Kodituwakku, P.W. Neurocognitive profile in children with fetal alcohol spectrum disorders. Dev. Disabil. Res. Rev. 2009, 15, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Kodituwakku, P.W. A neurodevelopmental framework for the development of interventions for children with fetal alcohol spectrum disorders. Alcohol 2010, 44, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Mattson, S.N.; Goodman, A.M.; Caine, C.; Delis, D.C.; Riley, E.P. Executive functioning in children with heavy prenatal alcohol exposure. Alcohol. Clin. Exp. Res. 1999, 23, 1808–1815. [Google Scholar] [CrossRef] [PubMed]
- McGee, C.L.; Schonfeld, A.M.; Roebuck-Spencer, T.M.; Riley, E.P.; Mattson, S.N. Children with heavy prenatal alcohol exposure demonstrate deficits on multiple measures of concept formation. Alcohol. Clin. Exp. Res. 2008, 32, 1388–1397. [Google Scholar] [CrossRef] [PubMed]
- Vaurio, L.; Riley, E.P.; Mattson, S.N. Differences in executive functioning in children with heavy prenatal alcohol exposure or attention-deficit/hyperactivity disorder. J. Int. Neuropsychol. Soc. 2008, 14, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Kirk, H.E.; Gray, K.; Riby, D.M.; Cornish, K.M. Cognitive training as a resolution for early executive function difficulties in children with intellectual disabilities. Res. Dev. Dis. 2015, 38, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Snyder, H.R.; Miyake, A.; Hankin, B.L. Advancing understanding of executive function impairments and psychopathology: Bridging the gap between clinical and cognitive approaches. Front. Psychol. 2015, 6, 328. [Google Scholar] [CrossRef] [PubMed]
- Kerns, K.A.; Macoun, S.; MacSween, J.; Pei, J.; Hutchison, M. Attention and working memory training: A feasibility study in children with neurodevelopmental disorders. Appl. Neuropsychol. Child 2017, 6, 120–137. [Google Scholar] [CrossRef] [PubMed]
- Nash, K.; Stevens, S.; Greenbaum, R.; Weiner, J.; Koren, G.; Rovet, J. Improving executive functioning in children with fetal alcohol spectrum disorders. Child Neuropsychol. 2015, 21, 191–209. [Google Scholar] [CrossRef] [PubMed]
- Meck, W.H.; Smith, R.A.; Williams, C.L. Organizational changes in cholinergic activity and enhanced visuospatial memory as a functio of choline administered prenatally or postnatally or both. Behav. Neurosci. 1989, 103, 1234–1241. [Google Scholar] [CrossRef] [PubMed]
- Meck, W.H.; Williams, C.L. Perinatal choline supplementation increases the threshold for chunking in spatial memory. Neuroreport 1997, 8, 3053–3059. [Google Scholar] [CrossRef] [PubMed]
- Tees, R.C.; Mohammadi, E. The effects of neonatal choline dietary supplementation on adult spatial and configural learning and memory in rats. Dev. Psychobiol. 1999, 35, 226–240. [Google Scholar] [CrossRef]
- Cheng, R.K.; Meck, W.H. Prenatal choline supplementation increases sensitivity to time by reducing non-scalar sources of variance in adult temporal processing. Brain Res. 2007, 1186, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Kable, J.A.; Coles, C.D.; Keen, C.L.; Uriu-Adams, J.; Jones, K.L.; Yevtushok, L. The impact of micronutrient supplementation in alcohol-exposed pregnancies on information processing skills in Ukrainian infants. Alcohol 2015, 49, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.D.; La Fiette, M.H.; Quinn, V.R.; Riley, E.P. Neonatal choline supplementation ameliorates the effects of prenatal alcohol exposure on a discrimination learning task in rats. Neurotoxicol. Teratol. 2000, 22, 703–711. [Google Scholar] [CrossRef]
- Thomas, J.D.; Abou, E.J.; Dominguez, H.D. Prenatal choline supplementation mitigates the adverse effects of prenatal alcohol exposure on development in rats. Neurotoxicol. Teratol. 2009, 31, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H.; da Costa, K.A. Choline: An essential nutrient for public health. Nutr. Rev. 2009, 67, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H. What choline metabolism can tell us about the underlying mechanisms of fetal alcohol spectrum disorders. Mol. Neurobiol. 2011, 44, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Mehedint, M.G.; Niculescu, M.D.; Craciunescu, C.N.; Zeisel, S.H. Choline deficiency alters global histone methylation and epigenetic marking at the Re1 site of the calbindin 1 gene. FASEB J. 2010, 24, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Meck, W.H.; Williams, C.L. Metabolic imprinting of choline by its availability during gestation: Implications for memory and attentional processing across the lifespan. Neurosci. Biobehav. Rev. 2003, 27, 385–399. [Google Scholar] [CrossRef]
- Meck, W.H.; Williams, C.L.; Cermak, J.M.; Blusztajn, J.K. Developmental periods of choline sensitivity provide an ontogenetic mechanism for regulating memory capacity and age-related dementia. Front. Integr. Neurosci. 2008, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Pyapali, G.K.; Turner, D.A.; Williams, C.L.; Meck, W.H.; Swartzwelder, H.S. Prenatal dietary choline supplementation decreases the threshold for induction of long-term potentiation in young adult rats. J. Neurophysiol. 1998, 79, 1790–1796. [Google Scholar] [PubMed]
- Montoya, D.; Swartzwelder, H.S. Prenatal choline supplementation alters hippocampal N-methyl-D-aspartate receptor-mediated neurotransmission in adult rats. Neurosci. Lett. 2000, 296, 85–88. [Google Scholar] [CrossRef]
- Wong-Goodrich, S.J.; Glenn, M.J.; Mellott, T.J.; Blusztajn, J.K.; Meck, W.H.; Williams, C.L. Spatial memory and hippocampal plasticity are differentially sensitive to the availability of choline in adulthood as a function of choline supply in utero. Brain Res. 2008, 1237, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, J.R.; Fuglestad, A.J.; Eckerle, J.K.; Kroupina, M.G.; Miller, N.C.; Boys, C.J. Choline supplementation in children with fetal alcohol spectrum disorders has high feasibility and tolerability. Nutr. Res. 2013, 33, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, J.R.; Fuglestad, A.J.; Eckerle, J.K.; Fink, B.A.; Hoecker, H.L.; Boys, C.J. Choline supplementation in children with fetal alcohol spectrum disorders: A randomized, double-blind, placebo-controlled trial. Am. J. Clin. Nutr. 2015, 102, 1113–1125. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Risbud, R.D.; Mattson, S.N.; Chambers, C.D.; Thomas, J.D. Randomized, double-blind, placebo-controlled clinical trial of choline supplementation in school-aged children with fetal alcohol spectrum disorders. Am. J. Clin. Nutr. 2016, 104, 1683–1692. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.D.; Garrison, M.; O’Neill, T.M. Perinatal choline supplementation attenuates behavioral alterations associated with neonatal alcohol exposure in rats. Neurotoxicol. Teratol. 2004, 26, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.D.; Idrus, N.M.; Monk, B.R.; Dominguez, H.D. Prenatal choline supplementation mitigates behavioral alterations associated with prenatal alcohol exposure in rats. Birth Defects Res. Part A Clin. Mol. Teratol. 2010, 88, 827–837. [Google Scholar] [CrossRef] [PubMed]
- Baddeley, A.D.; Hitch, G.J. Developments in the concept of working memory. Neuropsychology 1994, 8, 485–493. [Google Scholar] [CrossRef]
- Nettelbeck, T.; Burns, N.R. Processing speed, working memory and reasoning ability from childhood to old age. Personal. Individ. Differ. 2010, 48, 379–384. [Google Scholar] [CrossRef]
- Zhan, J.Y.; Wilding, J.; Cornish, K.; Shao, J.; Xie, C.H.; Wang, Y.X. Charting the developmental trajectories of attention and executive function in Chinese school-aged children. Child Neuropsychol. 2011, 17, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Floresco, S.B.; Seamans, J.K.; Phillips, A.G. Selective roles for hippocampal, prefrontal cortical, and ventral striatal circuits in radial-arm maze tasks with or without a delay. J. Neurosci. 1997, 17, 1880–1890. [Google Scholar] [PubMed]
- Izaki, Y.; Maruki, K.; Hori, K.; Nomura, M. Effects of rat medial prefrontal cortex temporal inactivation on a delayed alternation task. Neurosci. Lett. 2001, 315, 129–132. [Google Scholar] [CrossRef]
- Izaki, Y.; Akema, T. Gamma-band power elevation of prefrontal local field potential after posterior dorsal hippocampus-prefrontal long-term potentiation induction in anesthetized rats. Exp. Brain Res. 2008, 184, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Shaw, C.; Aggleton, J.P. The effects of fornix and medial prefrontal lesions on delayed non-matching-to-sample by rats. Behav. Brain Res. 1993, 54, 91–102. [Google Scholar] [CrossRef]
- Yoon, T.; Okada, J.; Jung, M.W.; Kim, J.J. Prefrontal cortex and hippocampus subserve different components of working memory in rats. Learn. Mem. 2008, 15, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, A.H.; Handa, R.J. Fetal alcohol exposure produces delay-dependent memory deficits in juvenile and adult rats. Alcohol. Clin. Exp. Res. 1997, 21, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Gur, R.C.; Richard, J.; Calkins, M.E.; Chiavacci, R.; Hansen, J.A.; Bilker, W.B. Age group and sex differences in performance on a computerized neurocognitive battery in children age 8–21. Neuropsychology 2012, 26, 251–265. [Google Scholar] [CrossRef] [PubMed]
- Satterthwaite, T.D.; Wolf, D.H.; Erus, G.; Ruparel, K.; Elliott, M.A.; Gennatas, E.D. Functional maturation of the executive system during adolescence. J. Neurosci. 2013, 33, 16249–16261. [Google Scholar] [CrossRef] [PubMed]
- Paolozza, A.; Rasmussen, C.; Pei, J.; Hanlon-Dearman, A.; Nikkel, S.M.; Andrew, G. Working memory and visuospatial deficits correlate with oculomotor control in children with fetal alcohol spectrum disorder. Behav. Brain Res. 2014, 263, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Koss, W.A.; Belden, C.E.; Hristov, A.D.; Juraska, J.M. Dendritic remodeling in the adolescent medial prefrontal cortex and the basolateral amygdala of male and female rats. Synapse 2014, 68, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Van Eden, C.G.; Uylings, H.B. Cytoarchitectonic development of the prefrontal cortex in the rat. J. Comp. Neurol. 1985, 241, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Birrell, J.M.; Brown, V.J. Medial frontal cortex mediates perceptual attentional set shifting in the rat. J. Neurosci. 2000, 20, 4320–4324. [Google Scholar] [PubMed]
- Mooney, S.M.; Miller, M.W. Effects of prenatal exposure to ethanol on systems matching: The number of neurons in the ventrobasal thalamic nucleus of the mature rat. Dev. Brain Res. 1999, 117, 121–125. [Google Scholar] [CrossRef]
- Wellmann, K.A.; George, F.; Brnouti, F.; Mooney, S.M. Docosahexaenoic acid partially ameliorates deficits in social behavior and ultrasonic vocalizations caused by prenatal ethanol exposure. Behav. Brain Res. 2015, 286, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Wellmann, K.A.; Mooney, S.M. Unilateral whisker clipping exacerbates ethanol-induced social and somatosensory behavioral deficits in a sex- and age-dependent manner. Physiol. Behav. 2015, 148, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Farr, K.L.; Montano, C.Y.; Paxton, L.L.; Savage, D.D. Prenatal ethanol exposure decreases hippocampal 3H-vinylidene kainic acid binding in 45-day-old rats. Neurotoxicol. Teratol. 1988, 10, 563–568. [Google Scholar] [CrossRef]
- Marino, M.D.; Cronise, K.; Lugo, J.N.; Kelly, S.J., Jr. Ultrasonic vocalizations and maternal-infant interactions in a rat model of fetal alcohol syndrome. Dev. Psychobiol. 2002, 41, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Shors, T.J.; Lewczyk, C.; Pacynski, M.; Mathew, P.R.; Pickett, J. Stages of estrous mediate the stress-induced impairment of associative learning in the female rat. Neuroreport 1998, 9, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R.M.; Rawlins, J.N. T-maze alternation in the rodent. Nat. Protoc. 2006, 1, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Mackintosh, N.J. Selective attention in animal discrimination learning. Psychol. Bull. 1965, 64, 124–150. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, A.H.; Handa, R.J. Fetal alcohol exposure alters the induction of immediate early gene mRNA in the rat prefrontal cortex after an alternation task. Alcohol. Clin. Exp. Res. 1995, 19, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, A.H.; Handa, R.J. Fetal alcohol-exposed rats exhibit differential response to cholinergic drugs on a delay-dependent memory task. Neurobiol. Learn. Mem. 1999, 72, 230–243. [Google Scholar] [CrossRef] [PubMed]
- Schneider, R.D.; Thomas, J.D. Adolescent choline supplementation attenuates working memory deficits in rats exposed to alcohol during the third trimester equivalent. ACER 2016, 40, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, D.A.; Akers, K.G.; Rice, J.P.; Johnson, T.E.; Candelaria-Cook, F.T.; Maes, L.I.; Rosenberg, M.; Valenzuela, C.F.; Savage, D.D. Prenatal exposure to moderate levels of ethanol alters social behavior in adult rats: Relationship to structural plasticity and immediate early gene expression in frontal cortex. Behav. Brain Res. 2010, 207, 290–304. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.I.; Magcalas, C.M.; Barto, D.; Fink, B.C.; Rice, J.P.; Bird, C.W.; Davies, S.; Pentkowski, N.S.; Savage, D.D.; Hamilton, D.A. Effects of sex and housing on social, spatial, and motor behavior in adult rats exposed to moderate levels of alcohol during prenatal development. Behav. Brain Res. 2016, 313, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.L.; Meck, W.H.; Heyer, D.D.; Loy, R. Hypertrophy of basal forebrain neurons and enhanced visuospatial memory in perinatally choline-supplemented rats. Brain Res. 1998, 794, 225–238. [Google Scholar] [CrossRef]
- Meck, W.H.; Smith, R.A.; Williams, C.L. Pre- and postnatal choline supplementation produces long-term facilitation of spatial memory. Dev. Psychobiol. 1988, 21, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Coutcher, J.B.; Cawley, G.; Wecker, L. Dietary choline supplementation increases the density of nicotine binding sites in rat brain. J. Pharmacol. Exp. Ther. 1992, 262, 1128–1132. [Google Scholar] [PubMed]
- Cermak, J.M.; Blusztajn, J.K.; Meck, W.H.; Williams, C.L.; Fitzgerald, C.M.; Rosene, D.L.; Loy, R. Prenatal availability of choline alters the development of acetylcholinesterase in the rat hippocampus. Dev. Neurosci. 1999, 21, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Cansev, M. Synaptogenesis: Modulation by availability of membrane phospholipid precursors. Neuromol. Med. 2016, 18, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Bloem, B.; Poorthuis, R.B.; Mansvelder, H.D. Cholinergic modulation of the medial prefrontal cortex: The role of nicotinic receptors in attention and regulation of neuronal activity. Front. Neural Circuits 2014, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Luchicchi, A.; Bloem, B.; Viana, J.N.; Mansvelder, H.D.; Role, L.W. Illuminating the role of cholinergic signaling in circuits of attention and emotionally salient behaviors. Front. Synaptic Neurosci. 2014, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Koukouli, F.; Maskos, U. The multiple roles of the alpha7 nicotinic acetylcholine receptor in modulating glutamatergic systems in the normal and diseased nervous system. Biochem. Pharmacol. 2015, 97, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Janiesch, P.C.; Kruger, H.S.; Poschel, B.; Hanganu-Opatz, I.L. Cholinergic control in developing prefrontal-hippocampal networks. J. Neurosci. 2011, 31, 17955–17970. [Google Scholar] [CrossRef] [PubMed]
- Linke, R.; Frotscher, M. Development of the rat septohippocampal projection: Tracing with DiI and electron microscopy of identified growth cones. J. Comp. Neurol. 1993, 332, 69–88. [Google Scholar] [CrossRef] [PubMed]
- Mechawar, N.; Watkins, K.C.; Descarries, L. Ultrastructural features of the acetylcholine innervation in the developing parietal cortex of rat. J. Comp. Neurol. 2002, 443, 250–258. [Google Scholar] [CrossRef] [PubMed]
- McAlonan, G.M.; Dawson, G.R.; Wilkinson, L.O.; Robbins, T.W.; Everitt, B.J. The effects of AMPA-induced lesions of the medial septum and vertical limb nucleus of the diagonal band of broca on spatial delayed non-matching to sample and spatial learning in the water maze. Eur. J. Neurosci. 1995, 7, 1034–1049. [Google Scholar] [CrossRef] [PubMed]
- Torres, E.M.; Perry, T.A.; Blockland, A.; Wilkinson, L.S.; Wiley, R.G.; Lappi, D.A.; Dunnett, S.B. Behavioural, histochemical and biochemical consequences of selective immunolesions in discrete regions of the basal forebrain cholinergic system. Neuroscience 1994, 63, 95–122. [Google Scholar] [CrossRef]
- Bolam, J.P.; Wainer, B.H.; Smith, A.D. Characterization of cholinergic neurons in the rat neostriatum. A combination of choline acetyltransferase immunocytochemistry, golgi-impregnation and electron microscopy. Neuroscience 1984, 12, 711–718. [Google Scholar] [CrossRef]
- Prado, V.F.; Janickova, H.; Al-Onaizi, M.A.; Prado, M.A. Cholinergic circuits in cognitive flexibility. Neuroscience 2017, 345, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Ragozzino, M.E.; Choi, D. Dynamic changes in acetylcholine output in the medial striatum during place reversal learning. Learn. Mem. 2004, 11, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Tzavos, A.; Jih, J.; Ragozzino, M.E. Differential effects of M1 muscarinic receptor blockade and nicotinic receptor blockade in the dorsomedial striatum on response reversal learning. Behav. Brain Res. 2004, 154, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Mihalick, S.M.; Crandall, J.E.; Langlois, J.C.; Krienke, J.D.; Dube, W.V. Prenatal ethanol exposure, generalized learning impairment, and medial prefrontal cortical deficits in rats. Neurotoxicol. Teratol. 2001, 23, 453–462. [Google Scholar] [CrossRef]
- Marquardt, K.; Sigdel, R.; Caldwell, K.; Brigman, J.L. Prenatal ethanol exposure impairs executive function in mice into adulthood. ACER 2014, 38, 2962–2968. [Google Scholar] [CrossRef] [PubMed]
- Skorput, A.G.; Gupta, V.P.; Yeh, P.W.; Yeh, H.H. Persistent interneuronopathy in the prefrontal cortex of young adult offspring exposed to ethanol in utero. J. Neurosci. 2015, 35, 10977–10988. [Google Scholar] [CrossRef] [PubMed]
- An, L.; Zhang, T. Prenatal ethanol exposure impairs spatial cognition and synaptic plasticity in female rats. Alcohol 2015, 49, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Dias, R.; Robbins, T.W.; Roberts, A.C. Dissociation in prefrontal cortex of affective and attentional shifts. Nature 1996, 380, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Dias, R.; Robbins, T.W.; Roberts, A.C. Primate analogue of the Wisconsin card sorting test: Effects of excitotoxic lesions of the prefrontal cortex in the marmoset. Behav. Neurosci. 1996, 110, 872–886. [Google Scholar] [CrossRef] [PubMed]
- Bissonette, G.B.; Martins, G.J.; Franz, T.M.; Harper, E.S.; Schoenbaum, G.; Powell, E.M. Double dissociation of the effects of medial and orbital prefrontal cortical lesions on attentional and affective shifts in mice. J. Neurosci. 2008, 28, 11124–11130. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Ragozzino, M.E. The involvement of the orbitofrontal cortex in learning under changing task contingencies. Neurobiol. Learn. Mem. 2005, 83, 125–133. [Google Scholar] [CrossRef] [PubMed]
- McAlonan, K.; Brown, V.J. Orbital prefrontal cortex mediates reversal learning and not attentional set shifting in the rat. Behav. Brain Res. 2003, 146, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Chase, E.A.; Tait, D.S.; Brown, V.J. Lesions of the orbital prefrontal cortex impair the formation of attentional set in rats. Eur. J. Neurosci. 2012, 36, 2368–2375. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, H.S.; Wickens, R.; Tait, D.S.; Brown, V.J.; Dunnett, S.B. Lesions of the dorsomedial striatum impair formation of attentional set in rats. Neuropharmacology 2013, 71, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Kodali, V.N.; Jacobson, J.L.; Lindinger, N.M.; Dodge, N.C.; Molteno, C.D.; Meintjes, E.M.; Jacobson, S.W. Differential recruitment of brain regions during response inhibition in children prenatally exposed to alcohol. ACER 2017, 41, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Panczakiewicz, A.L.; Glass, L.; Coles, C.D.; Kable, J.A.; Sowell, E.R.; Wozniak, J.R.; Jones, K.L.; Riley, E.P.; Mattson, S.N. Neurobehavioral deficits consistent across age and sex in youth with prenatal alcohol exposure. ACER 2016, 40, 1971–1981. [Google Scholar] [CrossRef] [PubMed]
- Treit, S.; Chen, Z.; Zhou, D.; Baugh, L.; Rasmussen, C.; Andrew, G.; Pei, J.; Beaulieu, C. Sexual dimorphism of volume reduction but not cognitive deficit in fetal alcohol spectrum disorders: A combined diffusion tensor imaging, cortical thickness and brain volume study. NeuroImage Clin. 2017, 15, 284–297. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | Male Control | Male Ethanol | Female Control | Female Ethanol |
|---|---|---|---|---|
| Saline Untrained | 8 | 8 | 8 | 8 |
| Saline Trained | 7 | 8 | 8 | 8 |
| Choline Untrained | 8 | 8 | 8 | 7 |
| Choline Trained | 7 | 8 | 8 | 9 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Waddell, J.; Mooney, S.M. Choline and Working Memory Training Improve Cognitive Deficits Caused by Prenatal Exposure to Ethanol. Nutrients 2017, 9, 1080. https://doi.org/10.3390/nu9101080
Waddell J, Mooney SM. Choline and Working Memory Training Improve Cognitive Deficits Caused by Prenatal Exposure to Ethanol. Nutrients. 2017; 9(10):1080. https://doi.org/10.3390/nu9101080
Chicago/Turabian StyleWaddell, Jaylyn, and Sandra M. Mooney. 2017. "Choline and Working Memory Training Improve Cognitive Deficits Caused by Prenatal Exposure to Ethanol" Nutrients 9, no. 10: 1080. https://doi.org/10.3390/nu9101080
APA StyleWaddell, J., & Mooney, S. M. (2017). Choline and Working Memory Training Improve Cognitive Deficits Caused by Prenatal Exposure to Ethanol. Nutrients, 9(10), 1080. https://doi.org/10.3390/nu9101080